
Effects of Leaf Size and Defensive Traits on the Contribution of Soil Fauna to Litter Decomposition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Preparation
2.2. Meta-Analysis
2.3. Statistical Analysis
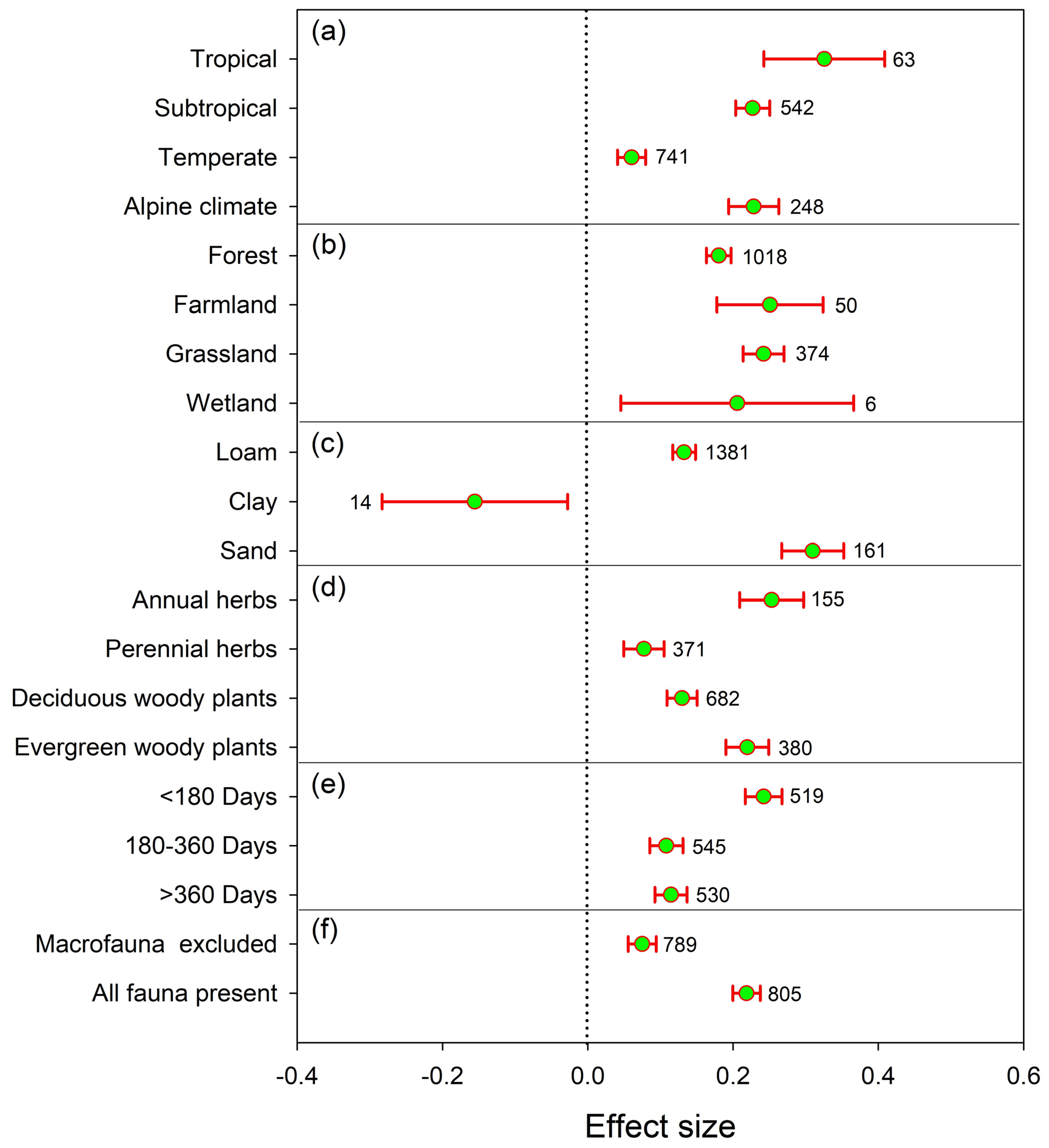
3. Results
3.1. Effects of Soil Fauna on Litter Decomposition
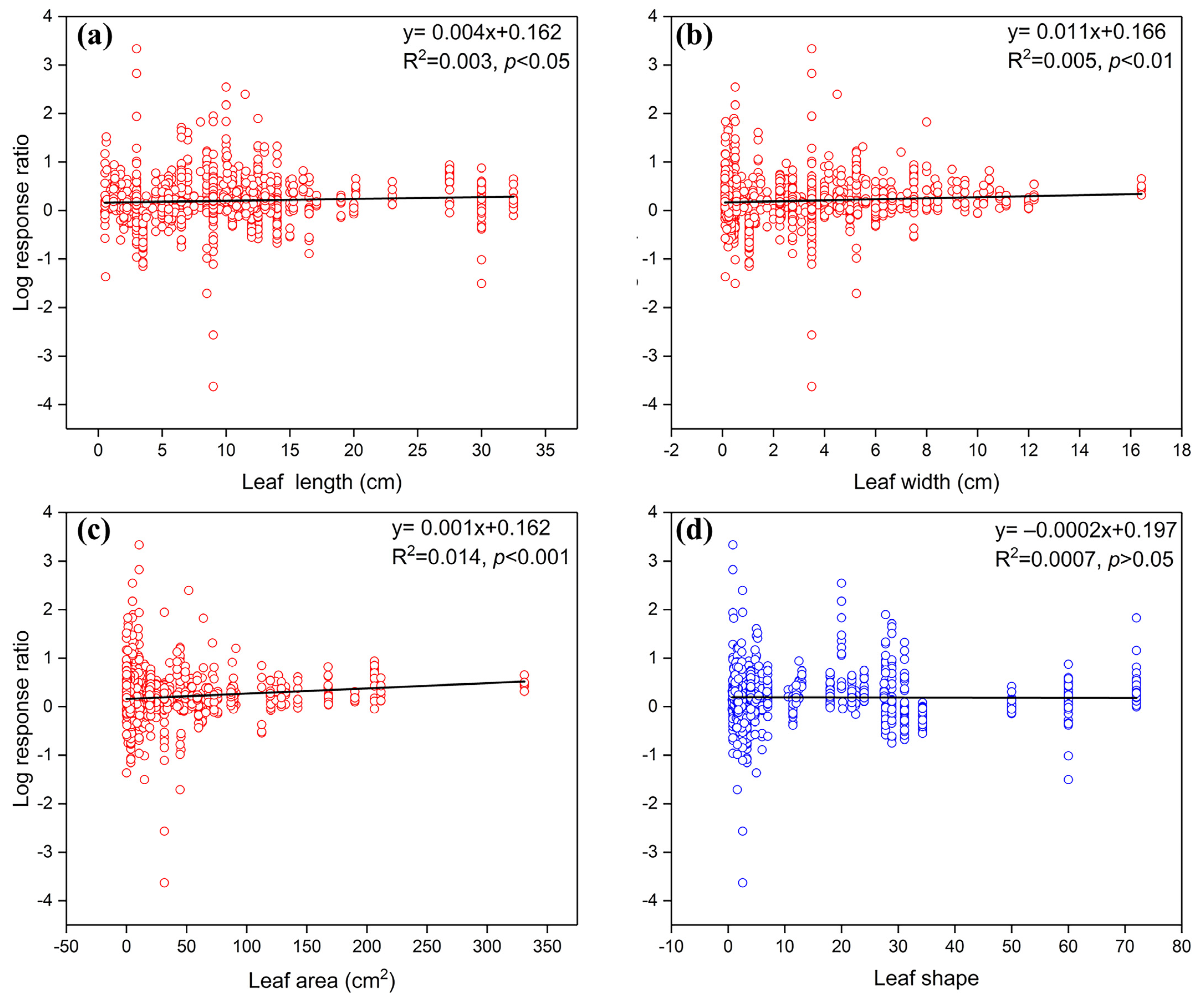
3.2. Effects of Litter Quality on Soil Fauna Modulate Litter Decomposition
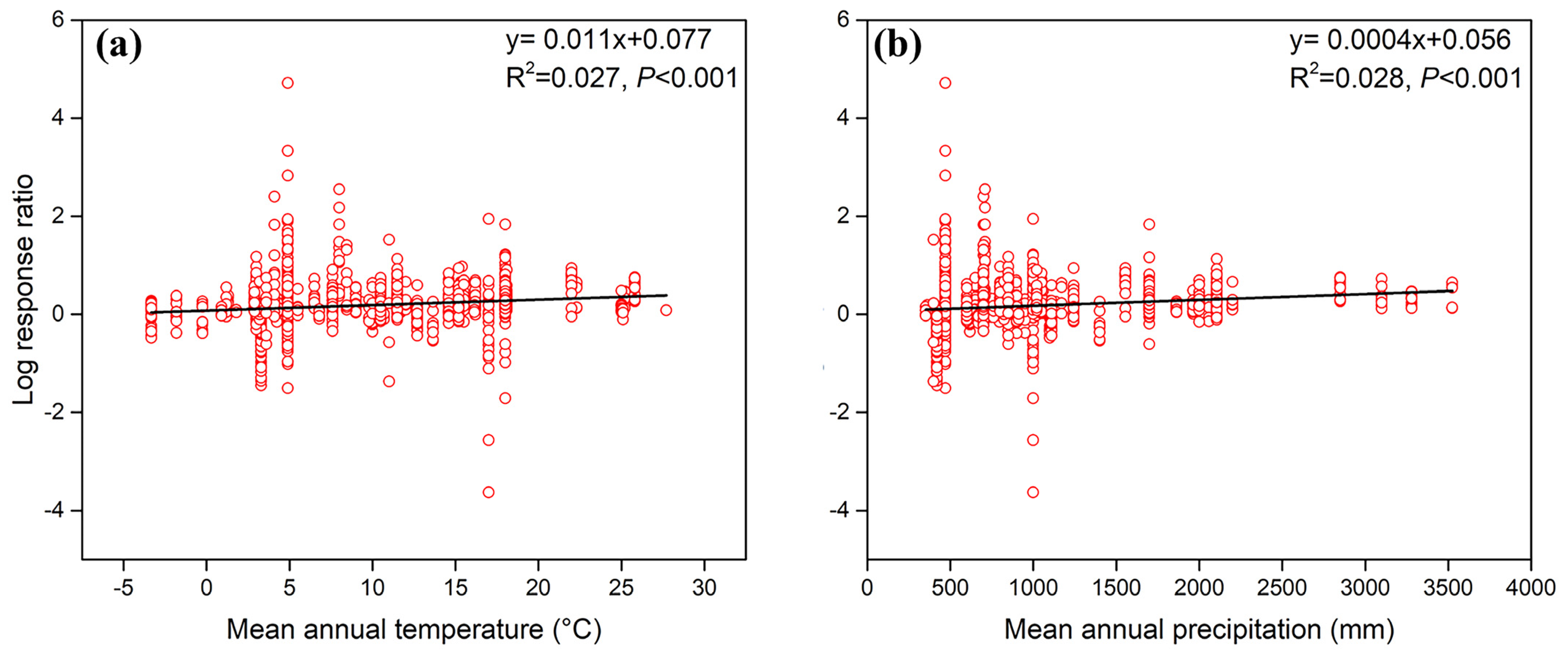
3.3. Effects of Climate on Soil Fauna Modulate Litter Decomposition
4. Discussion
4.1. Positive Effect of Soil Fauna on Litter Decomposition
4.2. Climate and Litter Quality Moderate the Effects of Soil Fauna on Litter Decomposition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dias, A.T.C.; Cornelissen, J.H.C.; Berg, M.P. Litter for life: Assessing the multifunctional legacy of plant traits. J. Ecol. 2017, 105, 1163–1168. [Google Scholar] [CrossRef]
- Zhang, W.P.; Fornara, D.; Yang, H.; Yu, R.P.; Callaway, R.M.; Li, L. Plant litter strengthens positive biodiversity–ecosystem functioning relationships over time. Trends Ecol. Evol. 2023, 38, 473–484. [Google Scholar] [CrossRef]
- Guo, L.B.; Sims, R.E.H. Litter decomposition and nutrient release via litter decomposition in New Zealand eucalypt short rotation forests. Agric. Ecosyst. Environ. 1999, 75, 133–140. [Google Scholar] [CrossRef]
- Giweta, M. Role of litter production and its decomposition, and factors affecting the processes in a tropical forest ecosystem: A review. J. Ecol. Environ. 2020, 44, 81–89. [Google Scholar] [CrossRef]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- García-Palacios, P.; Shaw, E.A.; Wall, D.H.; Hättenschwiler, S. Temporal dynamics of biotic and abiotic drivers of litter decomposition. Ecol. Lett. 2016, 19, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Butenschoen, O.; Scheu, S.; Eisenhauer, N. Interactive effects of warming, soil humidity and plant diversity on litter decomposition and microbial activity. Soil Biol. Biochem. 2011, 43, 1902–1907. [Google Scholar] [CrossRef]
- Schimel, J.P.; Gulledge, J.M.; Clein-Curley, J.S.; Lindstrom, J.E.; Braddock, J.F. Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biol. Biochem. 1999, 31, 831–838. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Djukic, I.; Kepfer-Rojas, S.; Schmidt, I.K.; Larsen, K.S.; Beier, C.; Berg, B.; Verheyen, K.; Caliman, A.; Paquette, A.; Gutiérrez-Girón, A.; et al. Early stage litter decomposition across biomes. Sci. Total Environ. 2018, 628, 1369–1394. [Google Scholar] [CrossRef]
- Parton, W.; Silver, W.L.; Burke, I.C.; Grassens, L.; Harmon, M.E.; Currie, W.S.; King, J.Y.; Adair, E.C.; Brandt, L.A.; Hart, S.C.; et al. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 361–364. [Google Scholar] [CrossRef]
- Peng, Y.; Peñuelas, J.; Vesterdal, L.; Yue, K.; Peguero, G.; Fornara, D.A.; Hedenec, P.; Steffens, C.; Wu, F.Z. Responses of soil fauna communities to the individual and combined effects of multiple global change factors. Ecol. Lett. 2022, 25, 1961–1973. [Google Scholar] [CrossRef]
- Jones, T.H.; Bradford, M.A. Assessing the functional implications of soil biodiversity in ecosystems. Ecol. Res. 2001, 16, 845–858. [Google Scholar] [CrossRef]
- Coleman, D.C.; Whitman, W.B. Linking species richness, biodiversity and ecosystem function in soil systems. Pedobiologia 2005, 49, 479–497. [Google Scholar] [CrossRef]
- Decaëns, T. Macroecological patterns in soil communities. Glob. Ecol. Biogeogr. 2010, 19, 287–302. [Google Scholar] [CrossRef]
- Pennisi, E. Spare a thought for the teeming ecosystem beneath your feet. Science 2020, 370, 1255–1256. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wang, F.; Fanin, N.; Pang, M.; Dou, P.; Wang, H.; Qian, S.; Zhao, L.; Yang, Y.; Mi, X.; et al. Soil fauna promote litter decomposition but do not alter the relationship between leaf economics spectrum and litter decomposability. Soil Biol. Biochem. 2019, 136, 107519. [Google Scholar] [CrossRef]
- Zimmer, M.; Kautz, G.; Topp, W. Do woodlice and earthworms interact synergistically in leaf litter decomposition? Funct. Ecol. 2005, 19, 7–16. [Google Scholar] [CrossRef]
- Cárcamo, H.A.; Abe, T.A.; Prescott, C.E.; Holl, F.B.; Chanway, C.P. Influence of millipedes on litter decomposition, N mineralization, and microbial communities in a coastal forest in British Columbia, Canada. Can. Eur. J. For. Res. 2000, 30, 817–826. [Google Scholar] [CrossRef]
- Anderson, J.M. Succession, diversity and trophic relationships of some soil animals in decomposing leaf litter. J. Anim. Ecol. 1975, 44, 475–495. [Google Scholar] [CrossRef]
- David, J.P.; Ferran, A.; Gambier, J.; Meyran, J.C. Taste sensitivity of detritivorous mosquito larvae to decomposed leaf litter. J. Chem. Ecol. 2002, 28, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, P.M.; McNamara, N.P.; Chaplow, J.; Stott, A.W.; Black, H.I.J. Translocation of surface litter carbon into soil by Collembola. Soil Biol. Biochem. 2006, 38, 2655–2664. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Z.; Warren, M.W.; Chen, J. Mechanical fragmentation enhances the contribution of Collembola to leaf litter decomposition. Eur. J. Soil Biol. 2012, 53, 23–31. [Google Scholar] [CrossRef]
- Wickings, K.; Grandy, A.S. The oribatid mite Scheloribates moestus (Acari: Oribatida) alters litter chemistry and nutrient cycling during decomposition. Soil Biol. Biochem. 2011, 43, 351–358. [Google Scholar] [CrossRef]
- Elkins, N.Z.; Whitford, W.G. The role of microarthropods and nematodes in decomposition in a semi-arid ecosystem. Oecologia 1982, 55, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Koenning, S.R.; Hu, S. Root-parasitic nematodes enhance soil microbial activities and nitrogen mineralization. Microb. Ecol. 2003, 46, 134–144. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in terrestrial ecosystems. In The Quarterly Review of Biology; Anderson, D.J., Ed.; University of California Press: Berkely, CA, USA, 1979; Volume 56, p. 96. [Google Scholar] [CrossRef]
- Seastedt, T.R. The role of microarthropods in decomposition and mineralization processes. Annu. Rev. Entomol. 1984, 29, 25–46. [Google Scholar] [CrossRef]
- Tresch, S.; Frey, D.; Le Bayon, R.C.; Zanetta, A.; Rasche, F.; Fliessbach, A.; Moretti, M. Litter decomposition driven by soil fauna, plant diversity and soil management in urban gardens. Sci. Total Environ. 2019, 658, 1614–1629. [Google Scholar] [CrossRef]
- Ristok, C.; Leppert, K.N.; Scherer-Lorenzen, M.; Niklaus, P.A.; Bruelheide, H. Soil macrofauna and leaf functional traits drive the decomposition of secondary metabolites in leaf litter. Soil Biol. Biochem. 2019, 135, 429–437. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Santiago, L.S. Extending the leaf economics spectrum to decomposition: Evidence from a tropical forest. Ecology 2007, 88, 1126–1131. [Google Scholar] [CrossRef]
- de la Riva, E.G.; Prieto, I.; Villar, R. The leaf economic spectrum drives leaf litter decomposition in Mediterranean forests. Plant Soil 2019, 435, 353–366. [Google Scholar] [CrossRef]
- Mori, A.S.; Ota, A.T.; Fujii, S.; Seino, T.; Kabeya, D.; Okamoto, T.; Ito, M.T.; Kaneko, N.; Hasegawa, M. Biotic homogenization and differentiation of soil faunal communities in the production forest landscape: Taxonomic and functional perspectives. Oecologia 2015, 177, 533–544. [Google Scholar] [CrossRef]
- Fujii, S.; Berg, M.P.; Cornelissen, J.H.C. Living litter: Dynamic trait spectra predict fauna composition. Trends Ecol. Evol. 2020, 35, 886–896. [Google Scholar] [CrossRef]
- Walker, T.W.N.; Schrodt, F.; Allard, P.M.; Defossez, E.; Jassey, V.E.J.; Schuman, M.C.; Alexander, J.M.; Baines, O.; Baldy, V.; Bardgett, R.D.; et al. Leaf metabolic traits reveal hidden dimensions of plant form and function. Sci. Adv. 2023, 9, eadi4029. [Google Scholar] [CrossRef]
- Dossa, G.G.O.; Schaefer, D.; Zhang, J.L.; Tao, J.P.; Cao, K.F.; Corlett, R.T.; Cunningham, A.B.; Xu, J.C.; Cornelissen, J.H.C.; Harrison, R.D. The cover uncovered: Bark control over wood decomposition. J. Ecol. 2018, 106, 2147–2160. [Google Scholar] [CrossRef]
- Hanley, M.E.; Lamont, B.B.; Fairbanks, M.M.; Rafferty, C.M. Plant structural traits and their role in anti-herbivore defence. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 157–178. [Google Scholar] [CrossRef]
- Luo, Y.C.; Wang, L.; Cao, T.T.; He, W.H.; Lu, S.X.; Li, F.; Zhang, Z.W.; Chang, T.; Tian, X.J. Legacy effect of plant chemical defence substances on litter decomposition. Plant Soil 2023, 487, 93–108. [Google Scholar] [CrossRef]
- Xu, X.; Sun, Y.; Sun, J.J.; Cao, P.H.; Wang, Y.C.; Chen, H.Y.H.; Wang, W.F.; Ruan, H.H. Cellulose dominantly affects soil fauna in the decomposition of forest litter: A meta-analysis. Geoderma 2020, 378, 114620. [Google Scholar] [CrossRef]
- Zan, P.; Mao, Z.; Sun, T. Effects of soil fauna on litter decomposition in Chinese forests: A meta-analysis. PeerJ 2022, 10, e12747. [Google Scholar] [CrossRef]
- Shi, P.J.; Liu, M.D.; Yu, X.J.; Gielis, J.; Ratkowsky, D.A. Proportional relationship between leaf area and the product of leaf length and width of four types of special leaf shapes. Forests 2019, 10, 178. [Google Scholar] [CrossRef]
- Cristofori, V.; Rouphael, Y.; Gyves, E.M.D.; Bignami, C. A simple model for estimating leaf area of hazelnut from linear measurements. Sci. Hortic. 2007, 113, 221–225. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L. Leaf shape alters the coefficients of leaf area estimation models for Saussurea stoliczkai in central Tibet. Photosynthetica 2012, 50, 337–342. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zou, D.T.; Shrestha, N.; Xu, X.T.; Wang, Q.G.; Jia, W.; Wang, Z.H. Spatiotemporal variation in leaf size and shape in response to climate. J. Plant Ecol. 2020, 13, 87–96. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Zhou, X.H.; Zhang, B.C.; Lu, M.; Luo, Y.Q.; Liu, L.L.; Li, B. Different responses of soil respiration and its components to nitrogen addition among biomes: A meta-analysis. Glob. Chang. Biol. 2014, 20, 2332–2343. [Google Scholar] [CrossRef]
- Wang, D.J.; Zhou, H.K.; Yao, B.Q.; Wang, W.Y.; Dong, S.K.; Shang, Z.H.; She, Y.D.; Ma, L.; Huang, X.T.; Zhang, Z.H.; et al. Effects of nutrient addition on degraded alpine grasslands of the Qinghai-Tibetan Plateau: A meta-analysis. Agric. Ecosyst. Environ. 2020, 301, 106970. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 15 November 2023).
- Kampichler, C.; Bruckner, A. The role of microarthropods in terrestrial decomposition: A meta-analysis of 40 years of litterbag studies. Biol. Rev. 2009, 84, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Effects of macrofauna on soil properties in tropical ecosystems. Agric. Ecosyst. Environ. 1988, 24, 101–116. [Google Scholar] [CrossRef]
- Couteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and liter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhao, R.D.; Tian, Q.X.; Huang, L.; Wang, X.G.; Liu, F. Predominant effects of litter chemistry on lignin degradation in the early stage of leaf litter decomposition. Plant Soil 2019, 442, 453–469. [Google Scholar] [CrossRef]
- Bokhorst, S.; Wardle, D.A. Microclimate within litter bags of different mesh size: Implications for the ‘arthropod effect’ on litter decomposition. Soil Biol. Biochem. 2013, 58, 147–152. [Google Scholar] [CrossRef]
- Bradford, M.A.; Tordoff, G.M.; Jones, T.H.; Newington, J.E. Microbiota, fauna, and mesh size interactions in litter decomposition. Oikos 2002, 99, 317–323. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Rahman, M.M.; Tsukamoto, J.; Rahman, M.M.; Yoneyama, A.; Mostafa, K.M. Lignin and its effects on litter decomposition in forest ecosystems. Chem. Ecol. 2013, 29, 540–553. [Google Scholar] [CrossRef]
- Wang, X.F.; Liu, J.F.; Gao, W.Q.; Deng, Y.P.; Ni, Y.Y.; Xiao, Y.H.; Kang, F.F.; Wang, Q.; Lei, J.P.; Jiang, Z.P. Defense pattern of Chinese cork oak across latitudinal gradients: Influences of ontogeny, herbivory, climate and soil nutrients. Sci. Rep. 2016, 6, 27269. [Google Scholar] [CrossRef]
- McClaugherty, C.; Berg, B. Cellulose, lignin and nitrogen concentrations as rate regulating factors in late stages of forest litter decomposition. Pedobiologia 1987, 30, 101–112. [Google Scholar] [CrossRef]
- Fioretto, A.; Di Nardo, C.; Papa, S.; Fuggi, A. Lignin and cellulose degradation and nitrogen dynamics during decomposition of three leaf litter species in a Mediterranean ecosystem. Soil Biol. Biochem. 2005, 37, 1083–1091. [Google Scholar] [CrossRef]
- Rosenthal, R.; Rubin, D.B. A simple, general purpose display of magnitude of experimental effect. J. Educ. Psychol. 1982, 74, 166–169. [Google Scholar] [CrossRef]
- Cumming, G.; Fidler, F.; Kalinowski, P.; Lai, J. The statistical recommendations of the American Psychological Association Publication Manual: Effect sizes, confidence intervals, and meta-analysis. Aust. J. Psychol. 2012, 64, 138–146. [Google Scholar] [CrossRef]
- Ciccarelli, D.; Bona, C.; Carta, A. Coordination between leaf and root traits in Mediterranean coastal dune plants. Coord. Chem. Rev. 2023, 25, 973–980. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Intercept | MAT (°C) | MAP (100 mm) | MAP × MAT | Df | R² | AIC |
|---|---|---|---|---|---|---|---|
| lm0 | 0.183 *** | – | – | – | 1592 | – | 1894.837 |
| lm1 | 0.077 *** | 0.011 *** | – | – | 1591 | 0.027 | 1851.592 |
| lm2 | 0.056 ** | – | 0.012 *** | – | 1591 | 0.028 | 1850.542 |
| lm3 | 0.033 | 0.007 *** | 0.008 *** | – | 1590 | 0.035 | 1839.638 |
| lm4 | −0.028 | 0.013 *** | 0.015 *** | −0.0005 * | 1589 | 0.037 | 1837.566 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Yuan, F.; Xie, W.; Zuo, J.; Zhou, H. Effects of Leaf Size and Defensive Traits on the Contribution of Soil Fauna to Litter Decomposition. Forests 2024, 15, 481. https://doi.org/10.3390/f15030481
Wang D, Yuan F, Xie W, Zuo J, Zhou H. Effects of Leaf Size and Defensive Traits on the Contribution of Soil Fauna to Litter Decomposition. Forests. 2024; 15(3):481. https://doi.org/10.3390/f15030481
Chicago/Turabian StyleWang, Dangjun, Fang Yuan, Wuyang Xie, Juan Zuo, and Huakun Zhou. 2024. "Effects of Leaf Size and Defensive Traits on the Contribution of Soil Fauna to Litter Decomposition" Forests 15, no. 3: 481. https://doi.org/10.3390/f15030481