


Multi-Year Monitoring of Asclepias syriaca L. Spread in the Deliblato Sands Protected Reserve in Serbia
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
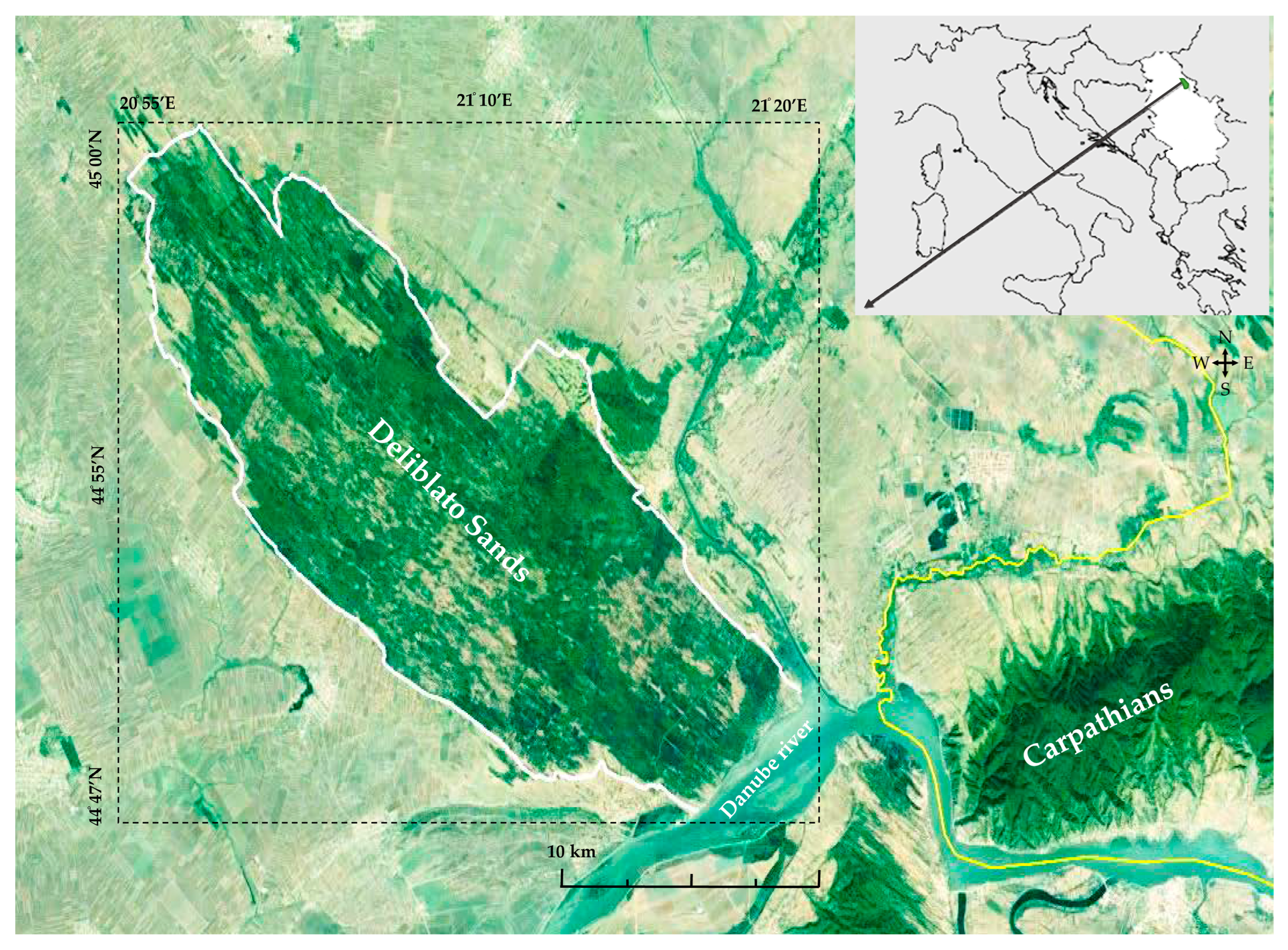
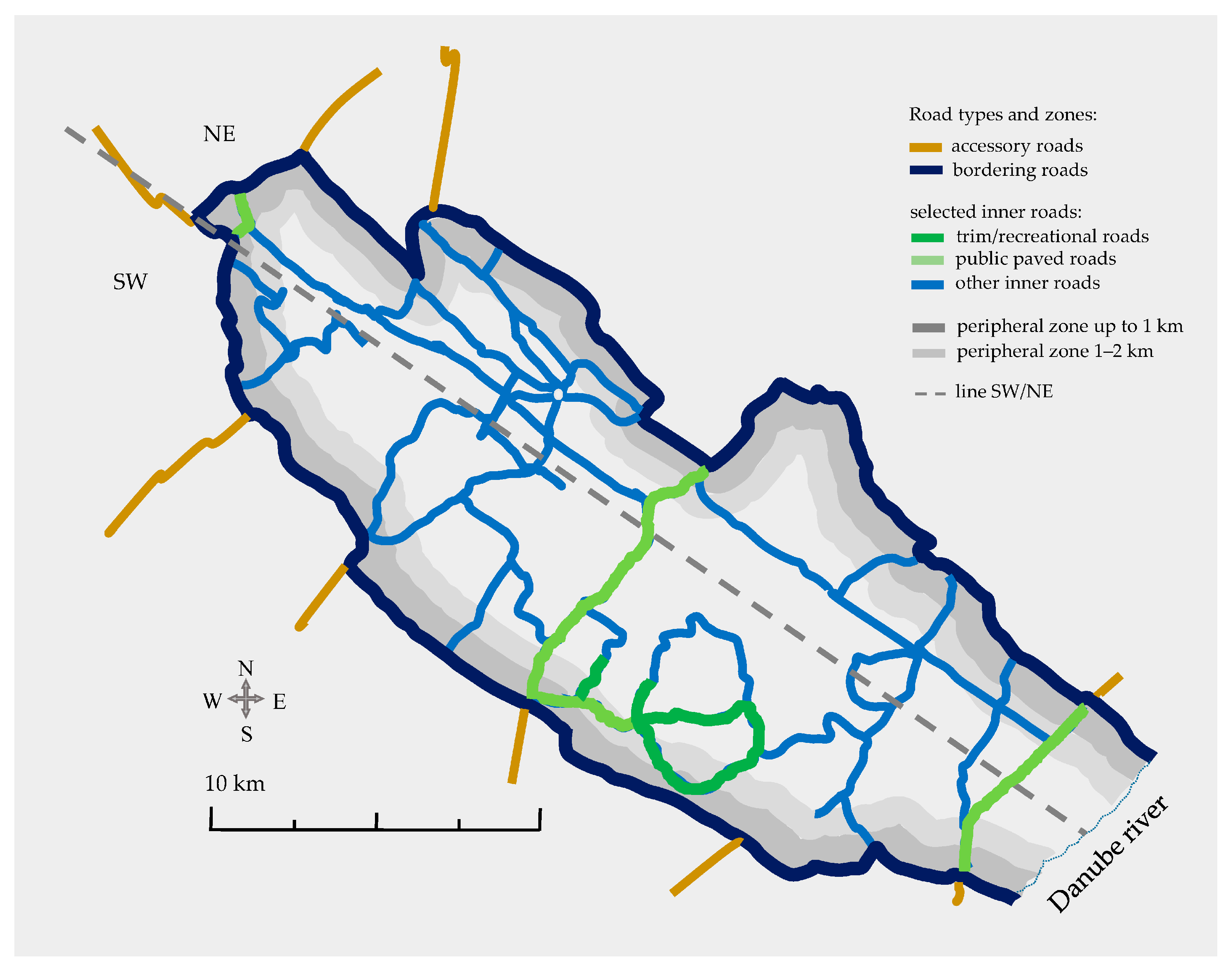
2.1. Research Object and Study Area
2.2. Field Methods
2.3. Data Analysis
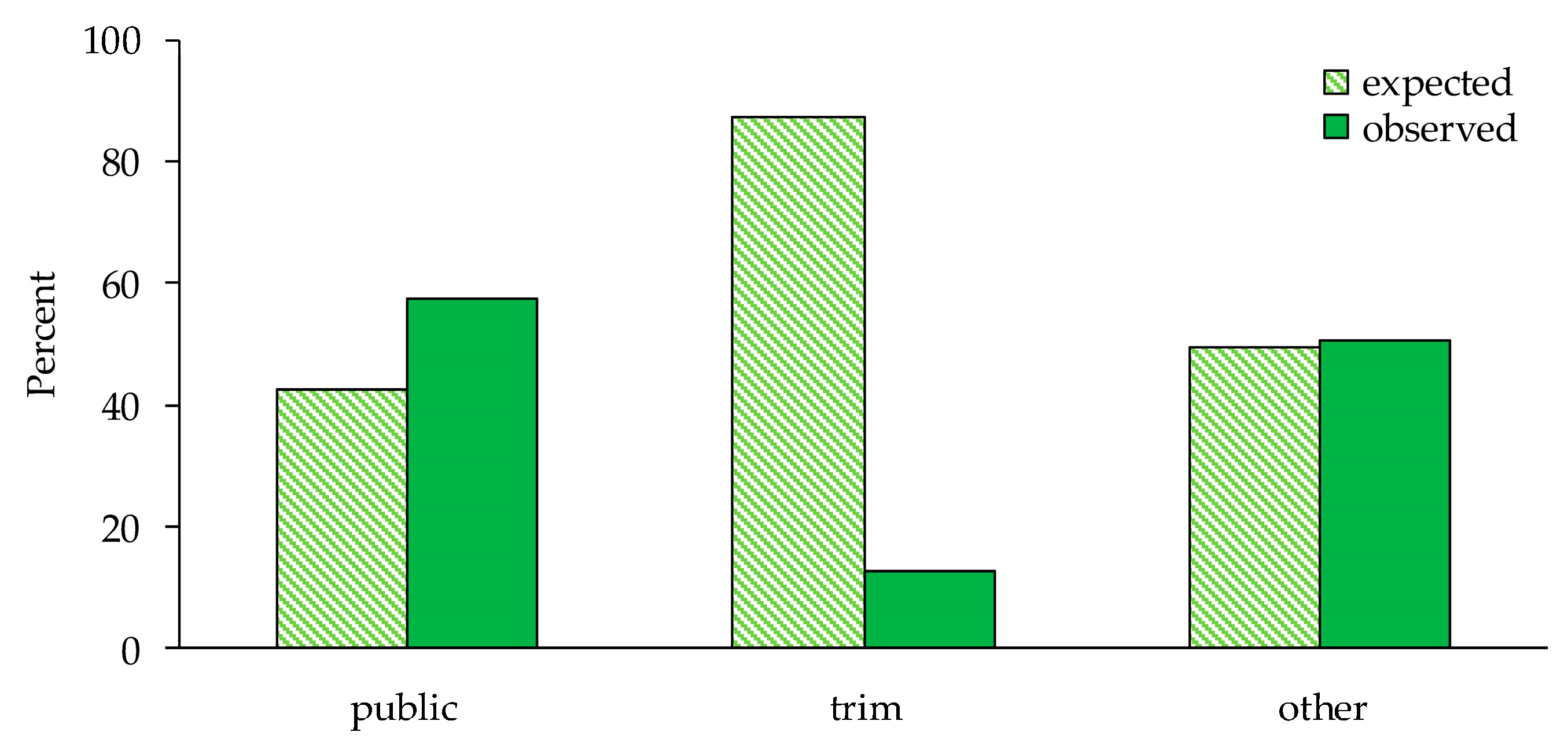
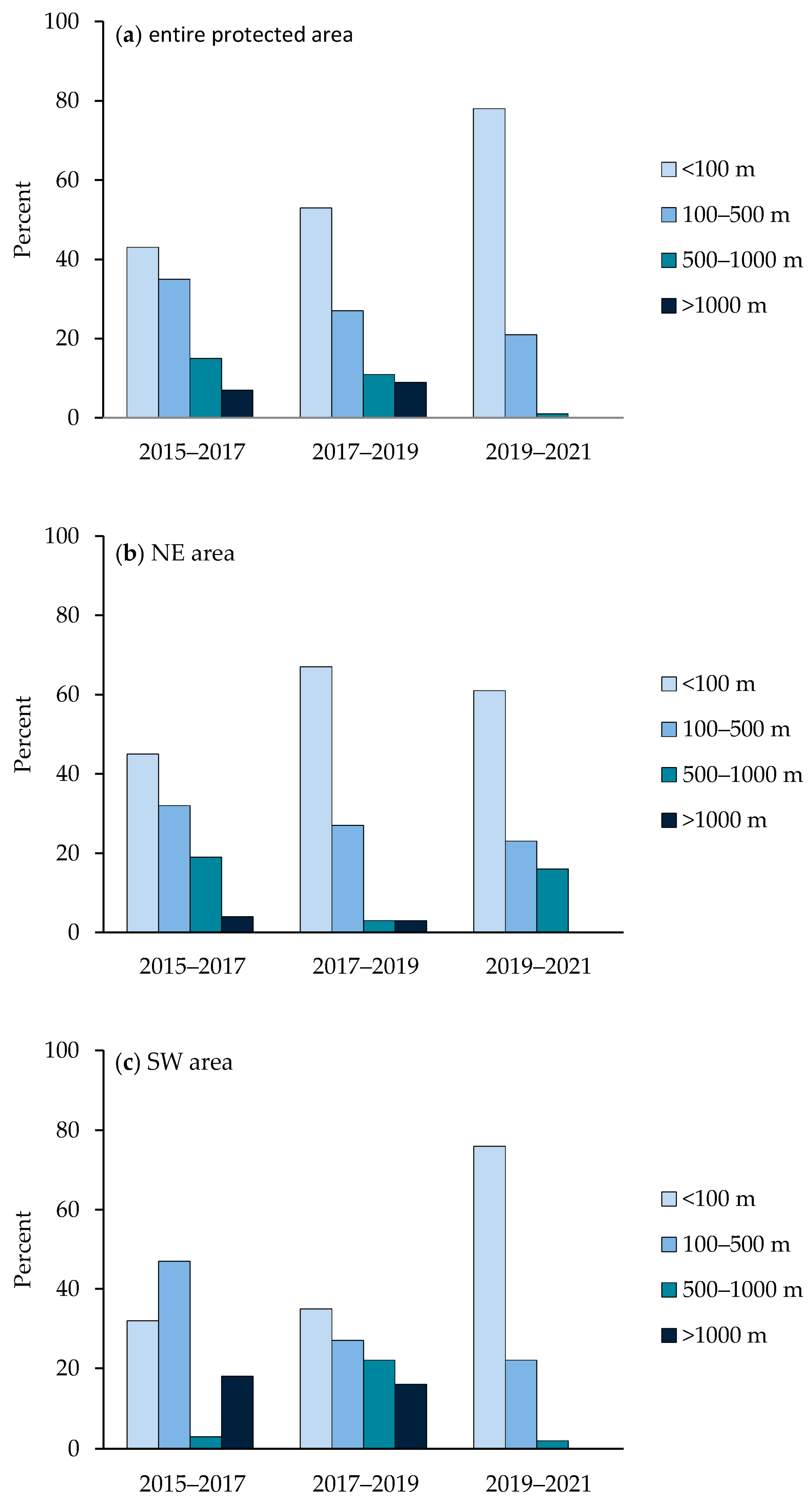
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Rejmanek, M.; Westbrooks, R. Introduced species: A significant component of human-caused global change. N. Z. J. Ecol. 1997, 21, 1–16. [Google Scholar]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef]
- Mollot, G.; Pantel, J.H.; Romanuk, T.N. The Effects of Invasive Species on the Decline in Species Richness: A Global Meta-Analysis. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Massol, F., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 56, pp. 61–83. [Google Scholar] [CrossRef]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- McGeoch, M.; Jetz, W. Measure and Reduce the Harm Caused by Biological Invasions. One Earth 2019, 1, 171–174. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Herrera, I.; Ferrer, P.; Jose, R.; Benzo, D.; Flores, S.; Garcia, B.; Jafet, M. An invasive succulent plant (Kalanchoe daigremontiana) influences soil carbon and nitrogen mineralization in a neotropical semiaridzone. Pedosphere 2018, 28, 632–643. [Google Scholar] [CrossRef]
- Shah, K.K.; Tiwari, I.; Tripathi, S.; Subedi, S.; Shrestha, J. Invasive alien plant species: A threat to biodiversity and agriculture in Nepal. Agriways 2020, 8, 62–73. [Google Scholar] [CrossRef]
- Shackleton, R.T.; Foxcroft, L.C.; Pyšek, P.; Wood, L.E.; Richardson, D.M. Assessing biological invasions in protected areas after 30 years: Revisiting nature reserves targeted by the 1980s SCOPE programme. Biol. Conserv. 2020, 243, 08424. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Kleijn, D.; Raemakers, I. A retrospective analysis of pollen host plant use by stable and declining bumble bee species. Ecology 2008, 89, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Tiedeken, E.J.; Egan, P.A.; Stevenson, P.C.; Wright, G.A.; Brown, M.J.F.; Power, E.F.; Farrell, I.; Matthews, S.M.; Stout, J.C. Nectar chemistry modulates the impact of an invasive plant on native pollinators. Funct. Ecol. 2016, 30, 885–893. [Google Scholar] [CrossRef]
- Mandelik, Y.; Winfree, R.; Neeson, T.; Kremen, C. Complementary habitat use by wild bees in agro-natural landscapes. Ecol. Appl. 2012, 22, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.L.; Traveset, A. A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecol. Lett. 2009, 12, 716–728. [Google Scholar] [CrossRef] [PubMed]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- Conroy, M.J.; Runge, M.C.; Nichols, J.D.; Stodola, K.W.; Cooper, R.J. Conservation in the face of climate change: The roles of alternative models, monitoring, and adaptation in confronting and reducing uncertainty. Biol. Conserv. 2011, 144, 1204–1213. [Google Scholar] [CrossRef]
- Foxcroft, L.C.; Pyšek, P.; Richardson, D.M.; Genovesi, P.; MacFadayen, S. Plant invasion science in protected areas: Progress and priorities. Biol. Invasions 2017, 19, 1353–1378. [Google Scholar] [CrossRef]
- Foxcroft, L.C.; Richardson, D.M.; Pyšek, P.; Genovesi, P. Invasive alien plants in protected areas: Threats, opportunities, and the way forward. In Plant Invasions in Protected Areas: Patterns, Problems and Challenges; Foxcroft, L.C., Richardson, D.M., Pyšek, P., Genovesi, P., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 621–639. [Google Scholar]
- Hulme, P.E. Protected land: Threat of invasive species. Science 2018, 361, 561–562. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pauchard, A.; Pyšek, P.; Vila, M.; Alba, C.; Blackburn, T.M.; Bullock, J.M.; Chytrý, M.; Dawson, W.; Dunn, A.M.; et al. Challenging the view that invasive non-native plants are not a significant threat to the floristic diversity of Great Britain. Proc. Nat. Acad. Sci. USA 2015, 112, E2988–E2989. [Google Scholar] [CrossRef]
- Pyšek, P.; Genovesi, P.; Pergl, J.; Monaco, A.; Wild, J. Plant invasions of protected areas in Europe: An old continent facing new problems. In Invading Nature; Foxcroft, L.C., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 209–240. [Google Scholar]
- Anačkov, G.T.; Rat, M.M.; Radak, B.D.; Igić, R.S.; Vukov, D.M.; Rućando, M.M.; Krstivojević, M.M.; Radulović, S.B.; Cvijanović, D.L.; Milić, D.M.; et al. Alien invasive neophytes of the Southeastern part of the Pannonian Plain. Cent. Eur. J. Biol. 2013, 8, 1032–1043. [Google Scholar] [CrossRef]
- Stevanović, V.B.; Vasić, V.F. Pregled antropogenih faktora koji ugrožavaju biodiverzitet Jugoslavije. In Biodiverzitet Jugoslavije-sa Pregledom Vrsta od Međunarodnog Značaja; Stevanović, V., Vasić, V., Eds.; Biološki Fakultet: Beograd, Serbia, 1995; pp. 19–35. [Google Scholar]
- Stevanović, V. Crvena knjiga flore Srbije = the Red data book of flora of Serbia. In Ministarstvo za Životnu Sredinu Republike SRBIJE, Biološki Fakultet Univerziteta u Beogradu; Zavod za Zaštitu Prirode Republike Srbije: Beograd, Serbia, 1999. [Google Scholar]
- Bhowmik, P.C. Biology and Control of Common Milkweed. Rev. Weed Sci. 1994, 6, 227–250. [Google Scholar]
- Evangelista, R.L. Milkweed seed wing removal to improve oil extraction. Ind. Crops Prod. 2007, 25, 210–217. [Google Scholar] [CrossRef]
- Mehran, A.; Mahmoud, N.; Mahmoud, Y.; Mehraban, S.M. Use of Iranian Milkweed Seed Oil to Increase Oxidative Stability of Olive Cultivar Roghani Oil. Int. J. Food Eng. 2017, 13, 20150315. [Google Scholar] [CrossRef]
- Martin, A.R.; Burnside, O.C. Common milkweed—Weed on the increase. Weeds Today 1980, 11, 19–20. [Google Scholar]
- Dvirna, T.S. Asclepias syriaca L. in the Romensko-Poltavsky Geobotanical District (Ukraine). Russ. J. Biol. Invasions 2018, 9, 29–37. [Google Scholar] [CrossRef]
- Bakacsy, L. Invasion impact is conditioned by initial vegetation states. Community Ecol. 2019, 20, 11–19. [Google Scholar] [CrossRef]
- Stef, R.; Manea, D.; Grozea, I.; Alexandru, C.; Gheorghescu, B.; Arsene, G.; Carabet, A. Asclepias syriaca a new segetal species in Romania. In Agriculture for Life, Life for Agriculture, Series A; Agronomy: Bucharest, Romania, 2022; pp. 6–8. [Google Scholar]
- Tokarska-Guzik, B.; Pisarczyk, E. Risk Assessment of Asclepias syriaca. Circabc Europa. 2015. Available online: https://circabc.europa.eu/sd/a/8dbd637b-6d8b-4608-b2b1-b51dd21cacde/Asclepias%20syriaca%20RA.pdf (accessed on 21 January 2024).
- Pravilnik o Listama Štetnih Organizama i Listama Biljaka, Biljnih Proizvoda i Propisanih Objekata. Grupa A2 Štetni Organizmi za Koje je Poznato da su Prisutni na Ograničenom Području Republike Srbije i Čije je Unošenje i Širenje u Republiku Srbiju Zabranjeno. Službeni Glasnik Republike Srbije 57. 2015. Available online: http://www.pravno-informacioni-sistem.rs/SlGlasnikPortal/eli/rep/sgrs/ministarstva/pravilnik/2010/7/1/reg (accessed on 21 January 2024).
- Dar, P.A.; Reshi, Z.A.; Shah, M.A. Roads act as corridors for the spread of alien plant species in the mountainous regions: A case study of Kashmir Valley, India. Trop. Ecol. 2015, 56, 183–190. [Google Scholar]
- Joly, M.; Bertrand, P.; Gbangou, R.Y.; White, M.C.; Dubé, J.; Lavoie, C. Paving the Way for Invasive Species: Road Type and the Spread of Common Ragweed (Ambrosia artemisiifolia). Environ. Manag. 2011, 48, 514–522. [Google Scholar] [CrossRef]
- Meunier, G.; Lavoie, C. Roads as Corridors for Invasive Plant Species: New Evidence from Smooth Bedstraw (Galium mollugo). Invasive Plant Sci. Manag. 2012, 5, 92–100. [Google Scholar] [CrossRef]
- Lemke, A.; Kowarik, I.; von der Lippe, W. How traffic facilitates population expansion of invasive species along roads: The case of common ragweed in Germany. J. Appl. Ecol. 2019, 56, 413–422. [Google Scholar] [CrossRef]
- Forman, J.; Child, L.; Brock, J.H.; Brundu, G.; Prach, K.; Pysěk, K.; Wade, P.M.; Williamson, M. The introduction of American plant species into Europe: Issues and consequences. In Proceedings of the 6th International Conference on the Ecology and Management of Alien Plant Invasions (EMAPi), Loughborough, UK, 12–15 September 2001. [Google Scholar]
- Dark, S.J. The biogeography of invasive alien plants in California: An application of GIS and spatial regression analysis. Divers. Distrib. 2004, 10, 1–9. [Google Scholar] [CrossRef]
- Essl, F. Ausbreitung und beginnende Einbürgerung von Spiraea japonica in Österreich. Bot. Helv. 2005, 115, 1–14. [Google Scholar] [CrossRef]
- Miller, N.P.; Matlack, G.R. Biodiversity research: Population expansion in an invasive grass, Microstegium vimineum: A test of the channelled diffusion model. Divers. Distrib. 2010, 16, 816–826. [Google Scholar] [CrossRef]
- Brothers, T.S.; Spingarn, A. Forest fragmentation and alien plant invasion of central Indiana old-growth forests. Conserv. Biol. 1992, 6, 91–100. [Google Scholar] [CrossRef]
- Eeel, F. Invasionsgeschichte und pflanzensoziologischer Anschluss der Aleppohirse (Sorghum halepense) am Beispiel des ostlichen Oberosterreich. Tuexenia 2005, 25, 251–268. [Google Scholar]
- Christen, D.C.; Matlack, G.R. The habitat and conduit functions of roads in the spread of three invasive plant species. Biol. Invasions 2009, 11, 453–465. [Google Scholar] [CrossRef]
- Bradly, D.; Petrovskaya, N. Propagation of invasive plant species in the presence of a road. J. Theor. Biol. 2022, 548, 111196. [Google Scholar] [CrossRef]
- Rauschert, E.S.J.; Mortensen, D.A.; Bloser, S.M. Human-mediated dispersal via rural road maintenance can move invasive propagules. Biol. Invasions 2017, 19, 2047–2058. [Google Scholar] [CrossRef]
- Dietz, H.; Edwards, P.J. Recognition that causal processes change during plant invasion helps explain conflicts in evidence. Ecology 2006, 87, 1359–1367. [Google Scholar] [CrossRef]
- Garnier, A.; Lecomte, J. Using a spatial nd stage-structured invasion model to assess the spread of feral populations of transgenic oilseed rape. Ecol. Modell. 2006, 194, 141–149. [Google Scholar] [CrossRef]
- Follak, S.; Bakacsy, L.; Essl, F.; Hochfellner, L.; Lapin, K.; Schwarz, M.; Tokarska-Guzik, B.; Wołkowycki, D. Monograph of invasive plants in Europe N°6: Asclepias syriaca L. Bot. Lett. 2021, 168, 422–451. [Google Scholar] [CrossRef]
- Bhowmik, P.C.; Bandeen, J.D. The biology of Canadian weeds: 19. Asclepias syriaca L. Can. J. Plant. Sci. 1976, 56, 579–589. [Google Scholar] [CrossRef]
- Stjepanović-Veseličić, L. Psamofitska vegetacija živih peskova Srbije. Srpska akademija nauka. Inst. Ekol. Biogeogr. Zb. Rad. 1956, 7, 3–27. [Google Scholar]
- Pavlović, P.; Kostić, N.; Karadžić, B.; Mitrović, M. The Soils of Serbia. In World Soils Book Series; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar]
- Erdős, L.; Tölgyesi, C.; Horzse, M.; Tolnay, D.; Hurton, Á.; Schulcz, N.; Körmöczi, L.; Lengyel, A.; Bátori, Z. Habitat complexity of the Pannonian forest-steppe zone and its nature conservation implications. Ecol. Complex 2014, 17, 107–118. [Google Scholar] [CrossRef]
- Erdős, L.; Ambarlı, D.; Anenkhonov, O.A.; Bátori, Z.; Cserhalmi, D.; Kiss, M.; Kröel-Dulay, G.; Liu, H.; Magnes, M.; Molnár, Z.; et al. The edge of two worlds: A new review and synthesis on Eurasian forest- steppes. Appl. Veg. Sci. 2018, 21, 345–362. [Google Scholar] [CrossRef]
- Antić, M.; Avdalović, V.; Jović, N. Evolution, Genetic Association and Ecological Value of Certain Deliblato Sands Types, Deliblato sand, Book of Papers I; Yugoslav Agricultural and Forest Centre: Pancevo, Serbia, 1969; pp. 47–66. [Google Scholar]
- Google Earth Pro 7.3.6.9345. Available online: https://www.google.com/earth/about/versions/ (accessed on 21 January 2024).
- SAS Institute Inc. SAS/STAT ® 9.1 User’s Guide; SAS Institute Inc., SAS Campus Drive: Cary, NC, USA, 2004; pp. 1429–1556. [Google Scholar]
- Tobin, P.C. Managing invasive species. F1000Research 2018, 7, 1686. [Google Scholar] [CrossRef] [PubMed]
- Robertson, P.A.; Mill, A.C.; Adriaens, T.; Moore, N.; Vanderhoeven, S.; Essl, F.; Booy, O. Risk management assessment improves the cost-effectiveness of invasive species prioritisation. Biology 2021, 10, 1320. [Google Scholar] [CrossRef]
- Gazoulis, I.; Antonopoulos, N.; Kanatas, P.; Karavas, N.; Bertoncelj, I.; Travlos, I. Invasive Alien Plant Species- Raising Awareness of a Threat to Biodiversity and Ecological Connectivity (EC) in the Adriatic-Ionian Region. Diversity 2022, 14, 387. [Google Scholar] [CrossRef]
- Simberloff, D. Eradication—Preventing invasions at the outset. Weed. Sci. 2003, 51, 247–253. [Google Scholar] [CrossRef]
- Jermanek, M. What tools do we have to detect invasive plant species. In Weed Risk Assessment; Groves, R.H., Panetta, F.D., Virtue, J.G., Eds.; CSIRO Publishing: Collingwood, Australia, 2001; pp. 3–9. [Google Scholar]
- Maxwell, B.; Lehnhoff, E.; Rew, L. The Rationale for Monitoring Invasive Plant Populations as a Crucial Step for Management. Invasive Plant Sci. Manag. 2009, 2, 1–9. [Google Scholar] [CrossRef]
- Bradley, B.A.; Allen, J.M.; O’Neill, M.W.; Wallace, R.D.; Bargeron, C.T.; Richburg, J.A.; Stinson, K. Invasive species risk assessments need more consistent spatial abundance data. Ecosphere 2018, 9, e02302. [Google Scholar] [CrossRef]
- Baard, J.A.; Kraaij, T. Use of a rapid roadside survey to detect potentially invasive plant species along the Garden Route, South Africa. Koedoe 2019, 61, 1–10. [Google Scholar] [CrossRef]
- Vila, M.; Ibáñez, I. Plant invasions in the landscape. Landsc. Ecol. 2011, 26, 461–472. [Google Scholar] [CrossRef]
- Paukova, Ž.; Knapekova, M.; Hauptvogl, M. Mapping of alien species of Asclepias syriaca and Fallopia japonica populations in the agricultural landscape. J. Cent. Eur. Agric. 2014, 15, 12–22. [Google Scholar] [CrossRef]
- Obradović, M. Analiza ekoloških uslova u procesu useljavanja biljaka u Vojvodinu. Zbonik Rad. PMF Novom. Sadu 1976, 6, 305–316. [Google Scholar]
- Kovacs, F. Óbecse Határának Virágos Növényei; Szegedi Nyomda: Szeged, Hungary, 1929; Available online: http://acta.bibl.u-szeged.hu/id/eprint/57209 (accessed on 21 January 2024).
- Igić, R.; Boža, P.; Anačkov, G.; Vukov, D.; Polić, D.; Borišev, M. Asclepias syriaca L. (cigansko perje) u flori Vojvodine. Zb. Rad. PMF. Ser. Biol. 2002, 31, 26–32. [Google Scholar]
- Bagi, I. Common milkweed (Asclepias syriaca L.). In The Most Important Invasive Plants in Hungary; Botta-Dukat, Z., Balogh, L., Eds.; Institute of Ecology and Botany-Hungarian Academy of Science: Budapest, Hungary, 2008; pp. 151–160. [Google Scholar]
- Jarić, S.; Mitrović, M.; Vrbničanin, S.; Karadžić, B.; Djurdjević, L.; Kostić, O.; Mačukanović Jocić, M.; Gajić, G.; Pavlović, P. A contribution to studies of the ruderal vegetation of southern Srem, Serbia. Arch. Biol. Sci. 2011, 63, 1181–1197. [Google Scholar] [CrossRef]
- Lazarević, P.; Stojanović, V.; Jelić, I.; Perić, R.; Krsteski, B.; Ajtić, R.; Sekulić, N.; Branković, S.; Sekulić, G.; Bjedov, V. Preliminarni spisak invazivnih vrsta u republici Srbiji sa opštim merama kontrole i suzbijanja kao potpora budućim zakonskim aktima. J. Nat. Conserv. 2012, 62, 5–31. [Google Scholar]
- Popov, M. Rasprostranjenost, Biološke Karakteristike i Suzbijanje Asclepias syriaca L. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2016. [Google Scholar]
- Christen, D.C.; Matlack, G. The role of roadsides in plant invasions: A demographic approach. Conserv. Biol. 2006, 20, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Benefield, C.; DiTomaso, J.; Kyser, G.; Orloff, S.; Churches, K.; Marcum, D.; Nader, G. Success of mowing to control yellow starthistle depends on timing and plant’s branching form. Calif. Agric. 1999, 53, 17–21. [Google Scholar] [CrossRef]
- Parendes, L.A.; Jones, J.A. Role of Light Availability and Dispersal in Exotic Plant Invasion Along Roads and Streams in the Hj Andrews Experimental Forest, Oregon. Conserv. Biol. 2000, 14, 64–75. [Google Scholar] [CrossRef]
- Gelbard, J.L.; Belnap, J. Roads as Conduits for Exotic Plant Invasions in a Semiarid Landscape. Conserv. Biol. 2003, 17, 420–432. [Google Scholar] [CrossRef]
- Follak, S.; Schleicher, C.; Schwarz, M. Roads support the spread of invasive Asclepias syriaca in Austria. Die Bodenkult. J. Land Manag. Food Environ. 2018, 69, 257–265. [Google Scholar] [CrossRef]
- Taylor, K.; Brummer, T.; Taper, M.L.; Wing, A.; Rew, L.J. Human-mediated long-distance dispersal: An empirical evaluation of seed dispersal by vehicles. Diversity Distrib. 2012, 18, 942–951. [Google Scholar] [CrossRef]
- Szilassi, P.; Soóky, A.; Bátori, Z.; Hábenczyus, A.A.; Frei, K.; Tölgyesi, C.; van Leeuwen, B.; Tobak, Z.; Csikós, N. Natura 2000 Areas, Road, Railway, Water, and Ecological Networks May Provide Pathways for Biological Invasion: A Country Scale Analysis. Plants 2021, 10, 2670. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.G.; Rocliffe, S.; Haddaway, N.R.; Dunn, A.M. The Role of Tourism and Recreation in the Spread of Non-Native Species: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0140833. [Google Scholar] [CrossRef] [PubMed]
- Bakacsy, L.; Tobak, Z.; van Leeuwen, B.; Szilassi, P.; Biró, C.; Szatmári, J. Drone-Based Identification and Monitoring of Two Invasive Alien Plant Species in Open Sand Grasslands by Six RGB Vegetation Indices. Drones 2023, 7, 207. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Kotowska, D.; Pärt, T.; Skórka, P.; Auffret, A.G.; Żmihorski, M. Scale dependence of landscape heterogeneity effects on plant invasions. J. Appl. Ecol. 2022, 59, 1313–1323. [Google Scholar] [CrossRef]
- Levine, J.M. Species diversity and biological invasions: Relating local process to community pattern. Science 2000, 288, 852–854. [Google Scholar] [CrossRef]
- Melbourne, B.A.; Cornell, H.V.; Davies, K.F.; Dugaw, C.J.; Elmendorf, S.; Freestone, A.L.; Hall, R.J.; Harrison, S.; Hastings, A.; Holland, M.; et al. Invasion in a heterogeneous world: Resistance, coexistence or hostile takeover? Ecol. Lett. 2007, 10, 77–94. [Google Scholar] [CrossRef]
- Zalai, M.; Poczok, L.; Dorner, Z.; Körösi, K.; Pálinkás, Z.; Szalai, M.; Pintér, O. Developing control strategies against common milkweed (Asclepias syriaca L.) on ruderal habitats. Herbologia 2017, 16, 69–84. [Google Scholar] [CrossRef]
- Bakacsy, L.; Bagi, I. Survival and regeneration ability of clonal common milkweed (Asclepias syriaca L.) after a single herbicide treatment in natural open sand grasslands. Sci. Rep. 2020, 10, 14222. [Google Scholar] [CrossRef] [PubMed]
- Kazinczi, G.; Mikulás, J.; Horváth, J.; Torma, M.; Hunyadi, K. Allelopathic effects of Asclepias syriaca roots on crops and weeds. Allelopath. J. 1999, 6, 267–270. [Google Scholar] [CrossRef]
- Popov, M.; Prvulović, D.; Šućur, J.; Vidović, S.; Samardžić, N.; Stojanović, T.; Konstantinović, B. Chemical Characterization of common mikweed (Asclepias Syriaca, L.) root extracts and their influence on maize (Zea mays L.), soybean (Glycine max (L.) Merr.) and sunflower (Helianthus annuus L.) seed germination and seedling growth. Appl. Ecol. Environ. Res. 2021, 19, 4219–4230. [Google Scholar] [CrossRef]
- Gaertner, M.; Fisher, J.; Sharma, G.; Esler, K. Insights into invasion and restoration ecology: Time to collaborate towards a holistic approach to tackle biological invasions. NeoBiota 2012, 12, 57–76. [Google Scholar] [CrossRef]
- Foxcroft, L.C.; Jarošík, V.; Pyšek, P.; Richardson, D.M.; Rouget, M. Protected-area boundaries as filters of plant invasions. Conserv. Biol. 2011, 25, 400–405. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
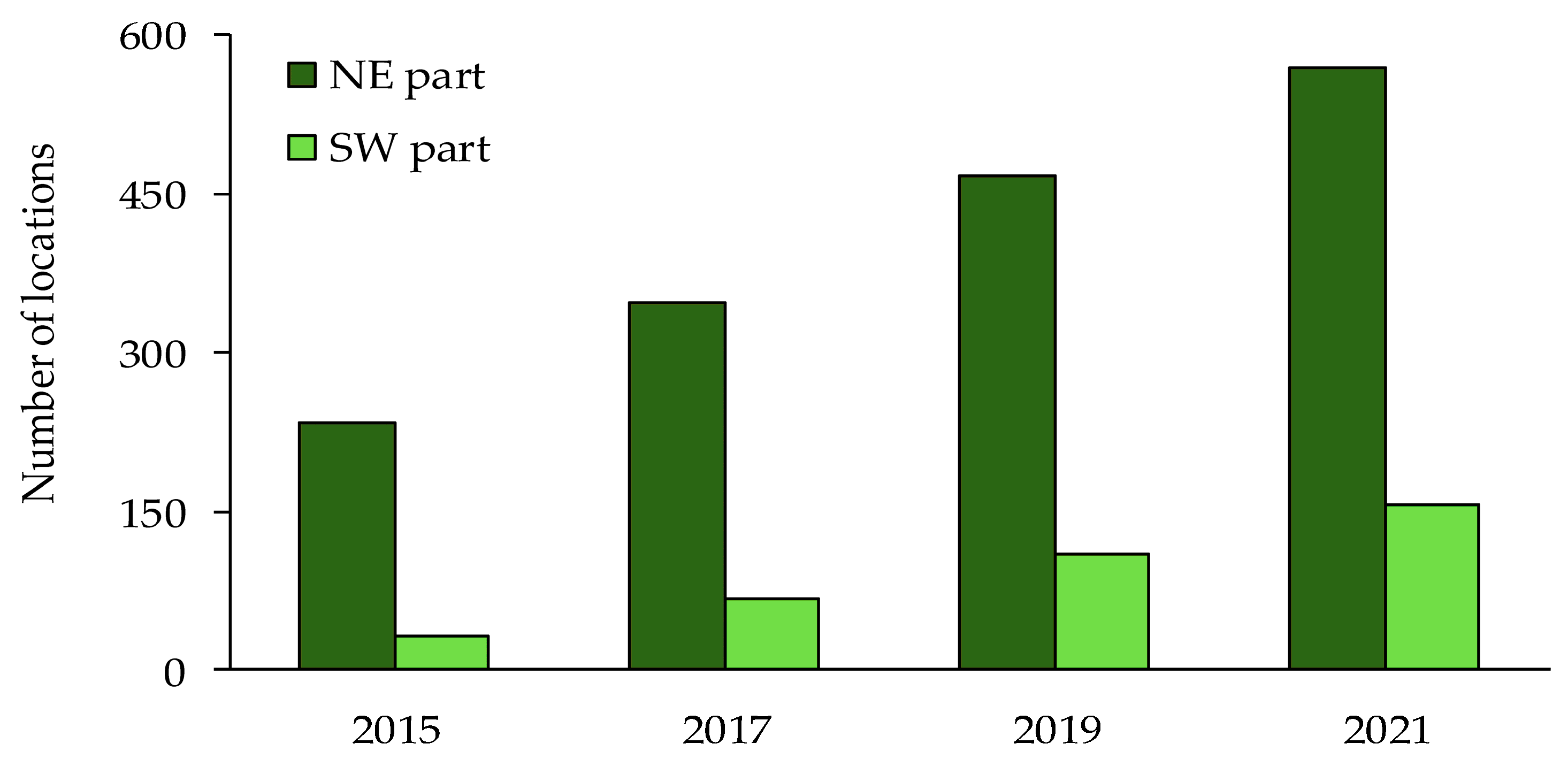{kind=link}
{kind=link}
{kind=link}
| Observed | Expected | χ2 | p | |
|---|---|---|---|---|
| (a) | Accessory and bordering roads vs. inner roads | |||
| NE part | ||||
| Accessory and bordering roads | 608 | 358 | 303.473 | 0.0001 |
| Selected inner roads | 235 | 485 | ||
| SW part | ||||
| Accessory and bordering roads | 33 | 23.9 | 5.425 | 0.0199 |
| Selected inner roads | 34 | 43.1 | ||
| (b) | Rate of emergence of new locations | |||
| 2015–2017 | 55% | 40 | 10.55 | 0.0051 |
| 2017–2019 | 39% | 40 | ||
| 2019–2021 | 26% | 40 | ||
| (c) | Dispersal pressure north-east vs. south-west, accessory and bordering roads | |||
| North-east | 608 | 356.2 | 400.681 | 0.0001 |
| South-west | 33 | 284.8 | ||
| 1 km peripheral zone | ||||
| North-east 1 km | 212 | 136 | 97.480 | 0.0001 |
| South-west 1 km | 29 | 105 | ||
| Observed | Expected | χ2 | p | |
|---|---|---|---|---|
| (a) | Peripheral (1 km) vs. remaining internal zone | |||
| The entire protected zone | ||||
| Peripheral zone 1 km | 241 | 125.6 | 128.202 | 0.0001 |
| Remaining inner zone | 486 | 601.4 | ||
| NE part | ||||
| Peripheral zone 1 km | 212 | 114.9 | 102.775 | 0.0001 |
| Remaining inner zone | 358 | 455.1 | ||
| Peripheral zone 2 km | 393 | 282.2 | 86.160 | 0.0001 |
| Remaining inner zone | 177 | 287.8 | ||
| SW part | ||||
| Peripheral zone 1 km | 31 | 22.6 | 3.647 | 0.0562 |
| Remaining inner zone | 126 | 134.4 | ||
| Peripheral zone 2 km | 67 | 55.4 | 3.753 | 0.0572 |
| Remaining inner zone | 90 | 101.6 | ||
| (b) | Frequency of occurrence of new locations | |||
| Type of selected inner roads (three roads) | ||||
| Public (paved) roads | 122 | 89.8 | 52.498 | 0.0001 |
| Recreational roads | 8 | 55.4 | ||
| Other inner roads | 597 | 581.8 | ||
| Type of selected inner roads (two roads) | ||||
| Public (paved) roads | 122 | 96 | 8.127 | 0.0044 |
| Other inner roads | 597 | 623 | ||
| Recreational roads | 8 | 52.6 | 41.418 | 0.0001 |
| Other inner roads | 597 | 552.4 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avramov, S.; Miljković, D.; Barišić Klisarić, N.; Živković, U.; Tarasjev, A. Multi-Year Monitoring of Asclepias syriaca L. Spread in the Deliblato Sands Protected Reserve in Serbia. Forests 2024, 15, 347. https://doi.org/10.3390/f15020347
Avramov S, Miljković D, Barišić Klisarić N, Živković U, Tarasjev A. Multi-Year Monitoring of Asclepias syriaca L. Spread in the Deliblato Sands Protected Reserve in Serbia. Forests. 2024; 15(2):347. https://doi.org/10.3390/f15020347
Chicago/Turabian StyleAvramov, Stevan, Danijela Miljković, Nataša Barišić Klisarić, Uroš Živković, and Aleksej Tarasjev. 2024. "Multi-Year Monitoring of Asclepias syriaca L. Spread in the Deliblato Sands Protected Reserve in Serbia" Forests 15, no. 2: 347. https://doi.org/10.3390/f15020347