
Diversity of Endomycorrhizal Fungi in Argan Forest Stands: Implications for the Success of Reforestation Programs
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
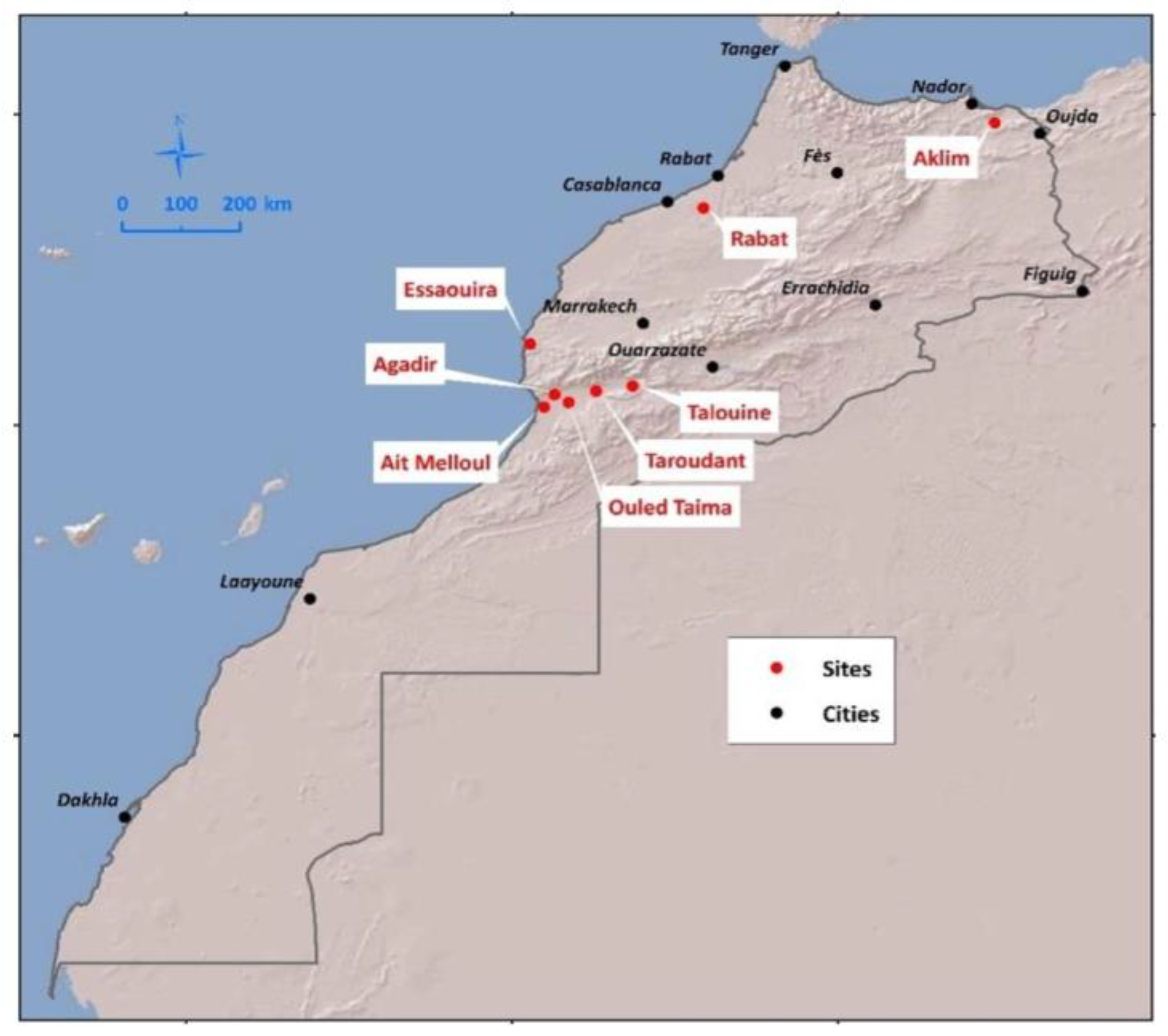
2.1. Sampling Sites in Argan Ecosystems
2.2. Collection of Soil and Root Samples
2.3. Physical and Chemical Analyses of Sampled Soil
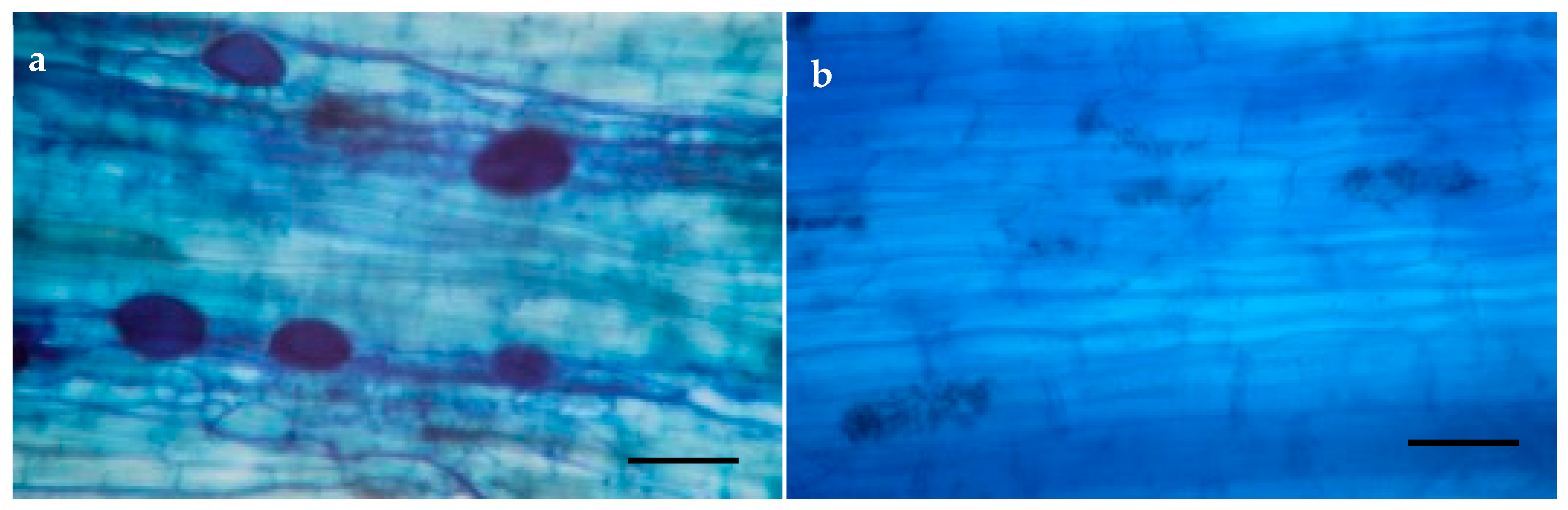
2.4. AMF Root Colonization
2.5. Extraction of AMF Spores from Rhizosphere Soil Mixtures and Identification of Spores
2.5.1. Spores’ Density
2.5.2. Species Richness
2.5.3. Occurrence Rate
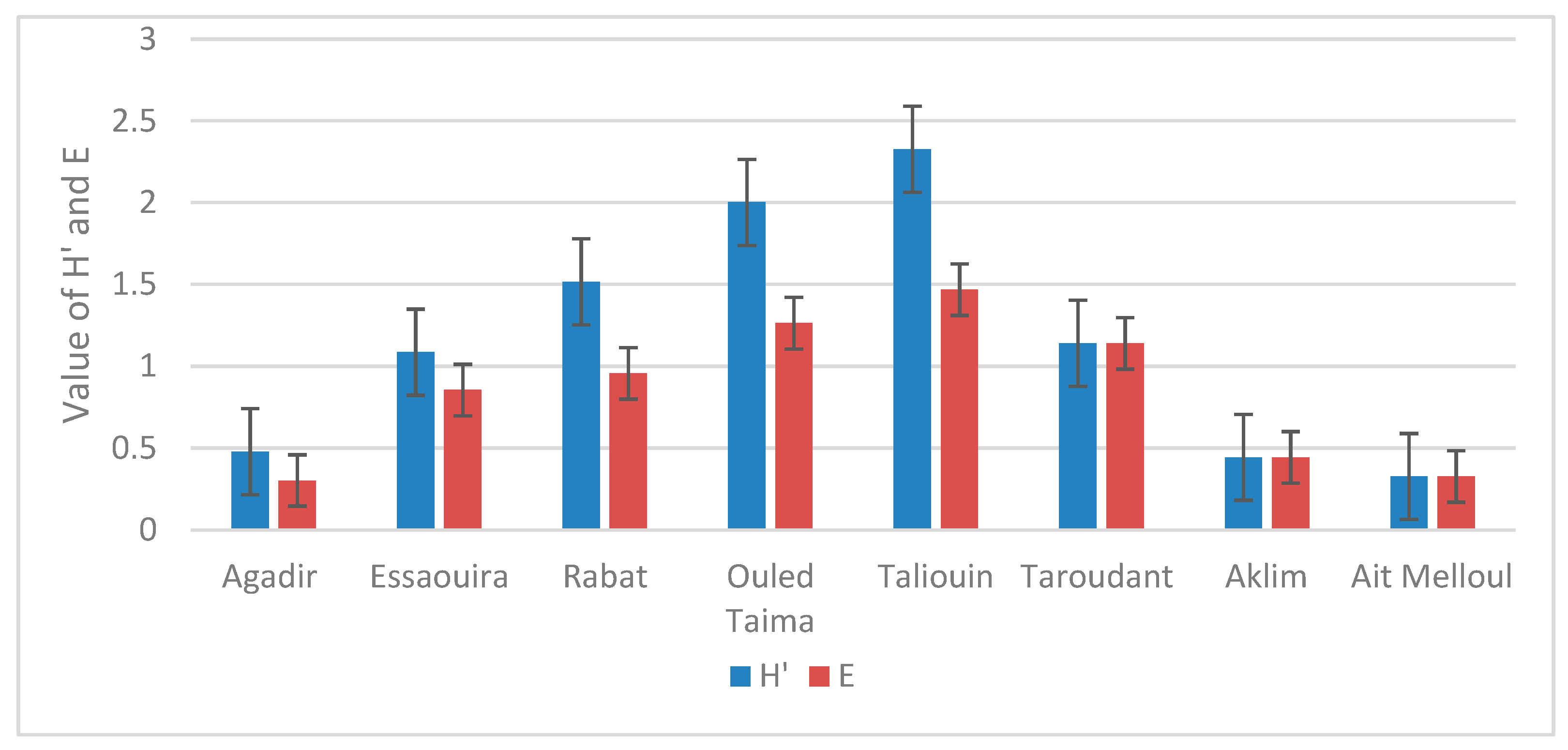
2.5.4. Shannon and Pielou Index
2.6. Statistical Analyses
3. Results
3.1. Soil Physical Parameters
3.2. Root Colonization and Spore Density of AMF
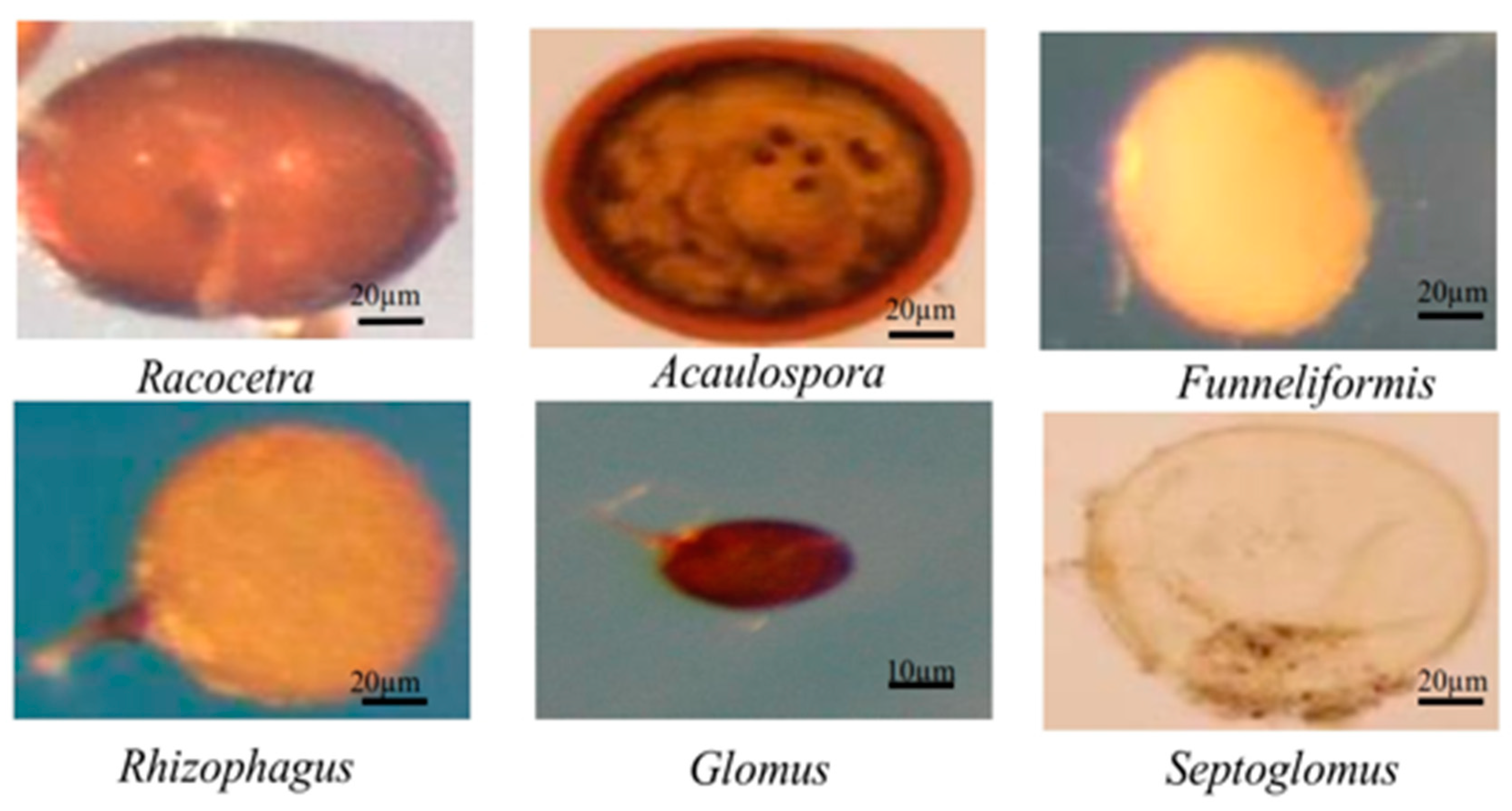
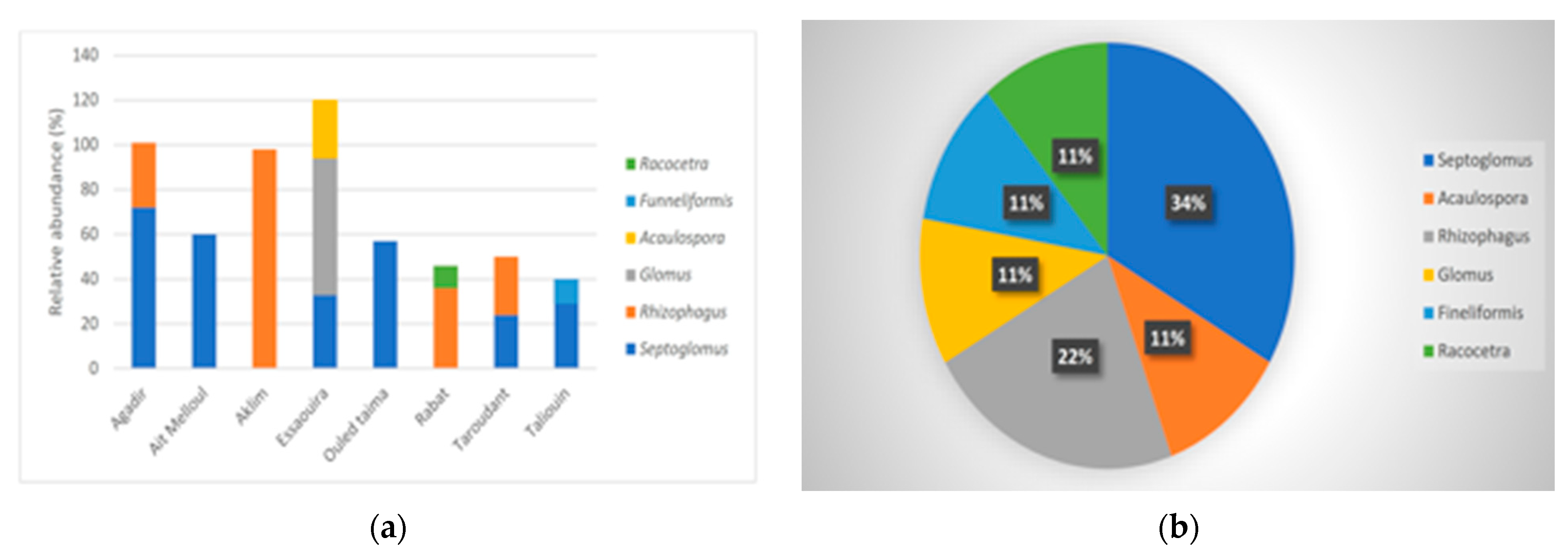
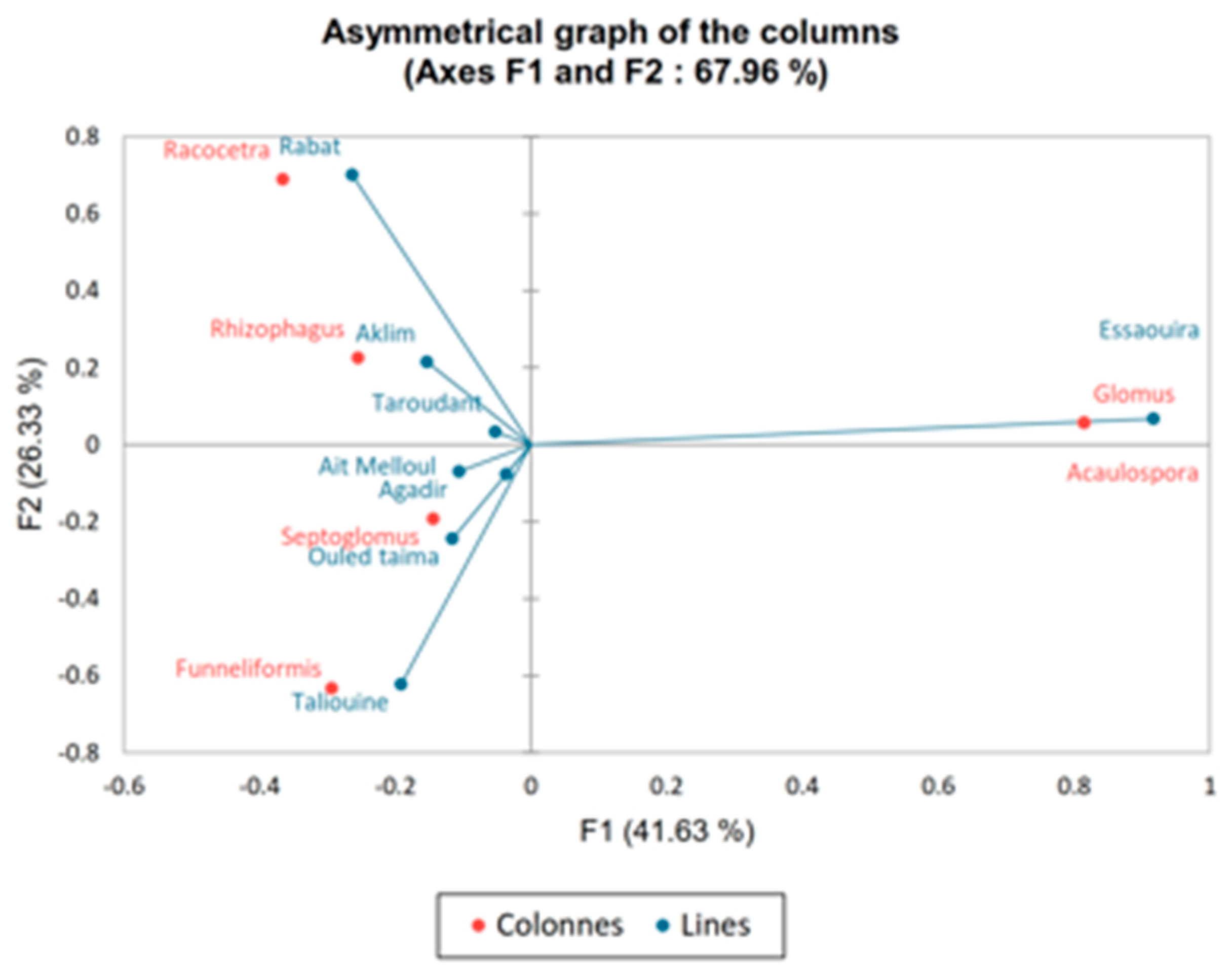
3.3. AMF Community Composition in Soils
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moukrim, S.; Lahssini, S.; Mharzi Alaoui, H.; Rifai, N.; Arahou, M.; Rhazi, L. Modélisation de la distribution spatiale des espèces endémiques pour leur conservation: Cas de l’Argania spinosa (L.) Skeels. Rev. DÉcologie Terre Vie 2018, 73, 153–166. [Google Scholar] [CrossRef]
- Mechqoq, H.; El Yaagoubi, M.; El Hamdaoui, A.; Momchilova, S.; Guedes da Silva Almeida, J.R.; Msanda, F.; El Aouad, N. Ethnobotany, Phytochemistry and Biological Properties of Argan Tree (Argania spinosa (L.) Skeels) (Sapotaceae)—A Review. J. Ethnopharmacol. 2021, 281, 114528. [Google Scholar] [CrossRef] [PubMed]
- Chaussod, R.; Adlouni, A.; et Christon, R. L’arganier et l’huile d’argane au Maroc: Vers la mutation d’un système agroforestier traditionnel? Enjeux et contribution de la recherche. Cah. Agric. 2005, 14, 351–356. [Google Scholar]
- Krajnc, B.; Bontempo, L.; Luis Araus, J.; Giovanetti, M.; Alegria, C.; Lauteri, M.; Augusti, A.; Atti, N.; Smeti, S.; Taous, F.; et al. Selective Methods to Investigate Authenticity and Geographical Origin of Mediterranean Food Products. Food Rev. Int. 2021, 37, 656–682. [Google Scholar] [CrossRef]
- Bousselmane, F.; Kenny, L.; et Achouri, M. Effet des mycorhizes à vésicules et arbuscules sur la croissance et la nutrition de l’arganier (Argania spinosa L.). Rev. Maroc. Des. Sci. Agron. Et. Vétérinaires 2022, 22, 193–198. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Del Mar Alguacil, M.; Torrecillas, E.; Kohler, J.; Roldán, A. A Molecular Approach to Ascertain the Success of “in Situ” AM Fungi Inoculation in the Revegetation of a Semiarid, Degraded Land. Sci. Total Environ. 2011, 409, 2874–2880. [Google Scholar] [CrossRef]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Chifetete, V.W.; Dames, J.F. Mycorrhizal Interventions for Sustainable Potato Production in Africa. Front. Sustain. Food Syst. 2020, 4, 593053. [Google Scholar] [CrossRef]
- Sellal, Z.; Ouazzani Touhami, A.; Chliyeh, M.; Dahmani, J.; Benkirane, R.; Douira, A. Arbuscular Mycorrhizal Fungi Species Associated with Rhizosphere of Argania spinosa (L.) Skeels in Morocco. Int. J. Pure Appl. Biosci. 2016, 4, 82–99. [Google Scholar] [CrossRef]
- Ganoudi, M.; Calonne-Salmon, M.; Ibriz, M.; Declerck, S. In Vitro Mycorrhization of Argania spinosa L. Using Germinated Seeds. Symbiosis 2021, 85, 57–68. [Google Scholar] [CrossRef]
- Requena, N.; Jimenez, I.; Toro, M.; Barea, J.M. Interactions between Plant-growth-promoting Rhizobacteria (PGPR), Arbuscular Mycorrhizal Fungi and Rhizobium Spp. in the Rhizosphere of Anthyllis Cytisoides, a Model Legume for Revegetation in Mediterranean Semi-arid Ecosystems. New Phytol. 1997, 136, 667–677. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis: Part 2; Page, A.L., Ed.; Chemical and Microbiological Properties; ASA and SSSA: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- El Mekkaoui, A.; Moussadek, R.; Mrabet, R.; Douaik, A.; El Haddadi, R.; Bouhlal, O.; Elomari, M.; Ganoudi, M.; Zouahri, A.; Chakiri, S. Effects of Tillage Systems on the Physical Properties of Soils in a Semi-Arid Region of Morocco. Agriculture 2023, 13, 683. [Google Scholar] [CrossRef]
- El Mekkaoui, A.; Moussadek, R.; Mrabet, R.; Chakiri, S.; Douaik, A.; Ghanimi, A.; Zouahri, A. The Conservation Agriculture in Northwest of Morocco (Merchouch Area): The Impact of No-till Systems on Physical Properties of Soils in Semi-Arid Climate. E3S Web Conf. 2021, 234, 37. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Trouvelot, A. Mesure Du Taux de Mycorhization VA d’un Système Radiculaire. Recherche de Méthodes d’estimation Ayant Une Signification Fonctionnelle. In Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; pp. 217–221. [Google Scholar]
- Gerdemann, J.W.; Nicolson, T.H. Spores of Mycorrhizal Endogone Species Extracted from Soil by Wet Sieving and Decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Cranenbrouck, S.; Voets, L.; Bivort, C.; Renard, L.; Strullu, D.-G.; Declerck, S. Methodologies for in Vitro Cultivation of Arbuscular Mycorrhizal Fungi with Root Organs. In In Vitro Culture of Mycorrhizas; Declerck, S., Fortin, J.A., Strullu, D.-G., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2005; Volume 4, pp. 341–375. ISBN 978-3-540-24027-3. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 44–53. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Bossou, L.-D.R.; Houngnandan, H.B.; Adandonon, A.; Zoundji, C.; Houngnandan, P. DiversitÉ des champignons mycorhiziens arbusculaires associés à la culture du maïs (Zea mays L.) au Bénin. Int. J. Biol. Chem. Sci. 2019, 13, 597. [Google Scholar] [CrossRef]
- El Mrabet, S.; Ouahmane, L.; El Mousadik, A.; Msanda, F.; Abbas, Y. The Effectiveness of bucular Mycorrhizal Inoculation and Bio-Compost Addition for Enhancing Reforestation with Argania Spinosa in Morocco Available. Open J. For. 2014, 4, 14–23. [Google Scholar]
- Ouallal, I.; Abbas, Y.; Ech-cheddadi, S.; Ouajdi, M.; Ouhadach, M.; El Yacoubi, H.; Kerdouh, B.; El Goumi, Y.; Rochdi, A. Diversité des champignons endomycorhiziens de l’arganier et potentiel mycorhizogène des sols rhizosphériques des arganeraies du Sud-Ouest marocain. Bois For. Trop. 2019, 338, 73. [Google Scholar] [CrossRef]
- Jensen, A.; Jakobsen, I. The Occrrence of Vesicular-Arbuscular Mycorrhiza in Barley and Wheat Grown in Some Danish Soils with Different Fertilizer Treatments. Plant Soil. 1980, 55, 403–414. [Google Scholar] [CrossRef]
- Khaddari, A.E.; Gabardi, S.E.; Touhami, A.O.; Aoujdad, J.; Ouajdi, M.; Dahmani, J. diversity of endomycorrhizal fungi in 456 the thuya rhizosphere, sefrou region (middle eastern atlas, morocco). Plant Cell. Biotechnol. Mol. Biol. 2019, 20, 1143–1159. [Google Scholar]
- Walker, C.; Mize, C.W. Populations of Endogonaceous Fungi at Two Locations in Central Iowa. Can. J. Bot. 1982, 60, 2518–2529. Available online: https://cdnsciencepub.com/doi/abs/10.1139/b82-305 (accessed on 16 March 2023). [CrossRef]
- Diagne, O.; Ingleby, K. Ácologie des Champignons Mycorhiziens Arbusculaires Infectant Acacia Raddiana. In Un Arbre au Désert; Grouzis, M., Le Floc’h, É., Eds.; IRD Éditions: Marseille, France, 2003; pp. 205–228. ISBN 978-2-7099-1522-9. [Google Scholar]
- Heikham, E.; Rupam, K.; Bhoopander, G. Arbuscular mycorrhizal fungi in alleviation of salt stress: A review. Ann. Bot. 2009, 104, 1263–1280. [Google Scholar]
- Garbaye, J. La Symbiose Mycorhizienne, une Association Entre les Plantes et les Champignons; Editions Quae: Versailles, France, 2013; 251 Synthèses; ISBN 9782759219643. Available online: Http://Digital.Casalini.It/9782759219643 (accessed on 12 August 2022).
- Mohammad, M.J.; Malkawi, H.I.; Shibli, R. Effects of Arbuscular Mycorrhizal Fungi and Phosphorus Fertilization on Growth and Nutrient Uptake of Barley Grown on Soils with Different Levels of Salts. J. Plant Nutr. 2003, 26, 125–137. [Google Scholar] [CrossRef]
- Kachkouch, W.; Touati, J.; Ouazzani Touhami, A.; Filali-Maltouf, A.; El Modafar, C.; Moukhli, A.; Oukabli, A.; Benkirane, R.; Douira, A. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of Olea europaea in three regions of Morocco (Tafilalt, Zagora and Taounate). Int. J. Pure App. Biosci. 2014, 2, 178–195. [Google Scholar]
- Elmaati, Y.; Msanda, F.; Mousadik, A.E.; Elhamdaoui, A.; El, S.; Ouahmane, L. contribution to the characterization of mycorrhizae in the south west of morocco and their effect on growth parameters of Argania spinosa. Am. J. Innov. Res. Appl. Sci. 2015, 1, 235–243. [Google Scholar]
- Kenny, L.; Galiana, A.; Bellefontaine, R. Projet UE/MEDA/ADS. Appui a L’amelioration de la Situation de L’emploi de la Femme Rurale et Gestion Durable de L’arganeraie dans le Sud-ouest du Maroc—Theme 2: Multiplication Vegetative et Symbioses Racinaires de L’arganier: Optimisation des Agrosystemes a Base D’arganier; Agence du Developpement Social (Maroc): Rabat, Maroc; Agropolis (France): Montpellier, France, 2009; Rapport final; pp. 1–71. [Google Scholar]
- Abbas, Y.; Ducousso, M.; Abourouh, M.; Azcón, R.; Duponnois, R. Diversity of Arbuscular Mycorrhizal Fungi in Tetraclinis Articulata (Vahl) Masters Woodlands in Morocco. Ann. For. Sci. 2006, 63, 285–291. [Google Scholar] [CrossRef]
- Ouahmane, L.; Hafidi, M.; Thioulouse, J.; Ducousso, M.; Kisa, M.; Prin, M.; Galiana, A.; Boumezzough, A.; Duponnois, R. Improvement of Cupressus Atlantica Gaussen Growth by Inoculation with Native Arbuscular Mycorrhizal Fungi. J. Appl. Microbiol. 2006, 103, 1364–5072. [Google Scholar] [CrossRef] [PubMed]
- Sghir, F.; Chliyeh, M.; Kachkouch, W.; Khouader, M.; Touhami, A.O.; Benkirane, R.; Douira, A. Mycorrhizal status of Olea europaea spp. oleaster in Morocco. J. Appl. Biosci. 2013, 61, 4478–4489. [Google Scholar] [CrossRef]
- Khirani, S.; Boutaj, H.; Modafar, C.E.; Khelil, A.O.E. Arbuscular mycorrhizal fungi associated with date palm in Ouargla region (southeastern algeria). Plant Cell. Biotechnol. Mol. Biol. 2020, 21, 15–28. [Google Scholar]
- Gosling, P.; Hodge, A.; Goodlass, G.; Bending, G.D. Arbuscular Mycorrhizal Fungi and Organic Farming. Agric. Ecosyst. Environ. 2006, 113, 17–35. [Google Scholar] [CrossRef]
- Houngnandan, P.; Yemadje, R.G.H.; Kane, A.; Boeckx, P.; et Van Cleemput, O. Les glomales indigènes de la forêt claire à Isoberlinia doka (Craibet Stapf) à Wari-Maro au centre du Bénin. Tropicultura 2009, 27, 83–87. [Google Scholar]
- Dalpeé, Y. Inventaire de la flore endomycorhizienne des rivages et dunes maritimes du Québec, du Nouveau-Brunswick et de la Nouvelle-Écosse. Le Nat. Can. 1989, 116, 219–236. [Google Scholar]
- Gemma, J.N.; Koske, R.E.; Carreiro, M. Seasonal dynamics of selected species of VA mycorrhizal fungi in a sand dune. Mycol. Res. 1989, 92, 317–321. [Google Scholar] [CrossRef]
- Smith, T.F. The effect of season and crop rotation on the abundance of spores of vesicular-arbuscular (VA) mycorrhizal endophytes. Plant Soil 1980, 57, 475–479. [Google Scholar] [CrossRef]
- Zangaro, W.; Moreira, M. Micorrizas arbusculares nos biomas Floresta Atlân- tica e Floresta de Araucária. In Micorrizas 30 anos de Pesquisa no Brasil; Siqueira, J.O., de Souza, F.A., Cardoso, E.J.B.N., Tsai, S.M., Eds.; UFLA: Lavras, Brazil, 2010; pp. 279–310. [Google Scholar]
- Chagnon, P.L.; Bradley, R.L.; Maherali, H.; Klironomos, J.N. A trait-based framework to understand life history of mycorrhizal fungi. Trends Plant. Sci. 2013, 18, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Loján, P.; Senés-Guerrero, C.; Suárez, J.P.; Kromann, P.; Schüßler, A.; Declerck, S. Potato field-inoculation in Ecuador with Rhizophagus irregularis: No impact on growth performance and associated arbuscular mycorrhizal fungal communities. Symbiosis 2018, 73, 45–56. [Google Scholar] [CrossRef]
- Da Silva, I.R.; de Mello CM, A.; Neto RA, F.; da Silva DK, A.; de Melo, A.L.; Oehl, F.; Maia, L.C. Diversity of arbuscular mycorrhizal fungi along an environmental gradient in the Brazilian semi-arid. Appl. Soil. Ecol. 2014, 84, 166–175. [Google Scholar] [CrossRef]
- Prreira, C.M.R.; Goto, B.T.; Silva DKA, D.A.; Ferreira, A.C.A.; de Souza, F.A.; de Silva, G.A.; da Maia, L.C.; Oehl, F. Acaulospora Reducta Sp. Nov. and A. Excavata Two Glomeromycota Fungi with Pitted Spores from Brazil. Mycotaxon 2016, 130, 983–995. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | % Clay | % Silt | % Sand | pH Water | pH KCl | P2O5 ppm | K2O ppm | % SOM | % SOC |
|---|---|---|---|---|---|---|---|---|---|
| Agadir | 20.6 ± 0.4 d | 36.4 ± 0.2 d | 43.6 ± 0.3 d | 7.5 ± 0.1 bc | 7.2 ± 0.1 b | 21.1 ± 15.9 c | 588.8 ± 0.4 b | 4.3 ± 0.1 a | 1.8 ± 0.0 b |
| Ait Melloul | 4.9 ± 0.1 f | 15.3 ± 0.2 h | 78.9 ± 0.1 a | 7.3 ± 0.2 c | 7.3 ± 0.1 ab | 47.2 ± 0.0 b | 360.2 ± 0.1 f | 2.1 ± 0.1 bc | 1.3 ± 0.0 d |
| Aklim | 5.7 ± 0.1 f | 74.8 ± 0.2 a | 19.2 ± 0.2 g | 7.6 ± 0.1 bc | 7.5 ± 0.a | 14.1 ± 0.1 cd | 1971.2 ± 0.1 a | 4.4 ± 0.1 a | 2 ± 0.1 b |
| Essaouira | 9.0 ± 0.1 e | 44.7 ± 0.4 b | 44.4 ± 0.4 d | 7.8 ± 0.0 ab | 7.1 ± 0.1 b | 4.2 ± 0.0 d | 161.5 ± 0.1 g | 4.2 ± 0.1 a | 0.8 ± 0.0 e |
| Ouled Taima | 21.6 ± 0.2 c | 29.5 ± 0.3 | 48.1 ± 0.1 c | 7.4 ± 0.1 bc | 7.2 ± 0.1 b | 30.4 ± 0.1 cb | 404.1 ± 0.1 e | 0.9 ± 0.0 d | 0.6 ± 0.0 f |
| Rabat | 20.1 ± 0.2 d | 24.7 ± 0.1 f | 36.8 ± 0.1 e | 7.2 ± 0.1 bc | 7.3 ± 0.1 ab | 29.1 ± 0.1 c | 134.4 ± 0.1 h | 1.9 ± 0.1 c | 2.8 ± 0.0 a |
| Taliouin | 32 ± 0.5 a | 43.1 ± 0.5 c | 24.4 ± 0.3 f | 7.7 ± 0.1 bc | 7.2 ± 0.1 b | 69.0 ± 0.1 a | 555.7 ± 0.1 c | 2.03 ± 0.1 c | 1.2 ± 0.0 d |
| Taroudant | 20.9 ± 0.2 b | 23.2 ± 0.2 g | 53.8 ± 0.5 b | 8.2 ± 0.0 a | 7.1 ± 0.1 b | 47.2 ± 0.1 b | 495.1 ± 0.1 d | 2.3 ± 0.0 b | 1.3 ± 0.1 c |
| Sites | % F | % I | % V | % A | SD/100 g of Soil |
|---|---|---|---|---|---|
| Agadir | 100.0 ± 0.0 a | 22.3 ± 2.3 b | 35.3 ± 4.2 ab | 52.2 ± 6.4 a | 101 a |
| Ait Melloul | 72.0 ± 3.0 a | 26.1 ± 4.5 ab | 42.7 ± 4.0 a | 50.2 ± 9.9 a | 60 b |
| Aklim | 98.0 ± 1.6 ab | 36.6 ± 6.2 a | 34.5 ± 3.8 abc | 52.0 ± 5.7 a | 98 ab |
| Essaouira | 99.6 ± 0.3 bc | 30.8 ± 2.7 a | 31.5 ± 2.7 abc | 51.8 ± 6.1 a | 120 a |
| Ouled Taima | 84.0 ± 5.5 c | 25.7 ± 2.4 ab | 41.1 ± 2.9 a | 49.8 ± 3.6 a | 57 bc |
| Rabat | 84.0 ± 3.2 c | 22.9 ± 2.0 b | 21.5 ± 3.7 bcd | 52.4 ± 1.6 a | 46 bc |
| Taliouin | 80.0 ± 4.4 cd | 27.3 ± 10.3 ab | 52.8 ± 10.9 a | 38.2 ± 12.2 a | 50 bc |
| Taroudant | 88.0 ± 6.0 d | 19.5 ± 4.0 b | 48.7 ± 7.6 a | 40.3 ± 6.3 a | 40 c |
| %Clay | %Silt | %Sand | pH Water | pH KCl | P2O5 ppm | K2O ppm | %SOM | %SOC | |
| SD | −0.58754 | 0.54890 | −0.12613 | −0.05867 | −0.10489 | −0.86885 | 0.29033 | 0.88022 | −0.11987 |
| 0.1256 | 0.1589 | 0.7660 | 0.8902 | 0.8048 | 0.0051 ** | 0.4855 | 0.0039 ** | 0.7774 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganoudi, M.; Ouallal, I.; El Mekkaoui, A.; Mounir, M.; Ibriz, M.; Iraqi, D. Diversity of Endomycorrhizal Fungi in Argan Forest Stands: Implications for the Success of Reforestation Programs. Forests 2023, 14, 1649. https://doi.org/10.3390/f14081649
Ganoudi M, Ouallal I, El Mekkaoui A, Mounir M, Ibriz M, Iraqi D. Diversity of Endomycorrhizal Fungi in Argan Forest Stands: Implications for the Success of Reforestation Programs. Forests. 2023; 14(8):1649. https://doi.org/10.3390/f14081649
Chicago/Turabian StyleGanoudi, Matike, Imane Ouallal, Abdelali El Mekkaoui, Majid Mounir, Mohammed Ibriz, and Driss Iraqi. 2023. "Diversity of Endomycorrhizal Fungi in Argan Forest Stands: Implications for the Success of Reforestation Programs" Forests 14, no. 8: 1649. https://doi.org/10.3390/f14081649