

The Use of Compost Tea in a Containerized Urban Tree Nursery Shows No Evident Benefits to Tree Growth or Mycorrhizal Colonization

Abstract
:1. Introduction
- Compost tea application will have a positive effect on containerized tree growth rates;
- Compost tea application will have a positive effect on root mycorrhizal colonization percentages;
- Percentages of root mycorrhizal colonization will positively correlate with rates of tree growth.
2. Materials and Methods
2.1. Compost Tea Formula
2.2. Focal Species
2.3. Experimental Design and Maintenance
2.4. Plant Growth Measurements
2.5. Root Sample Collection and Processing
2.6. Assessment of Mycorrhizal Colonization
2.7. Microbial Community Profiling of Compost Tea
2.8. Data Analysis
3. Results
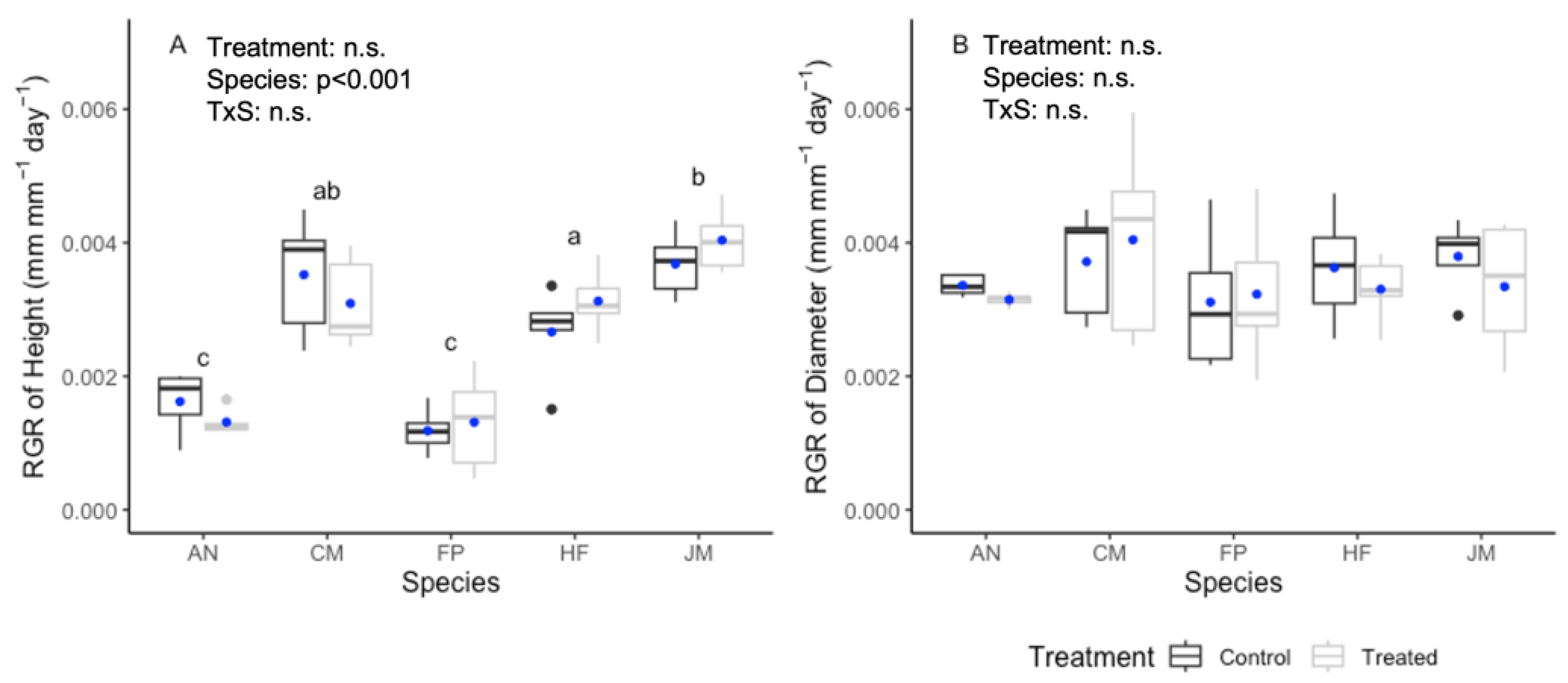
3.1. Plant Growth Response to Compost Tea Treatment
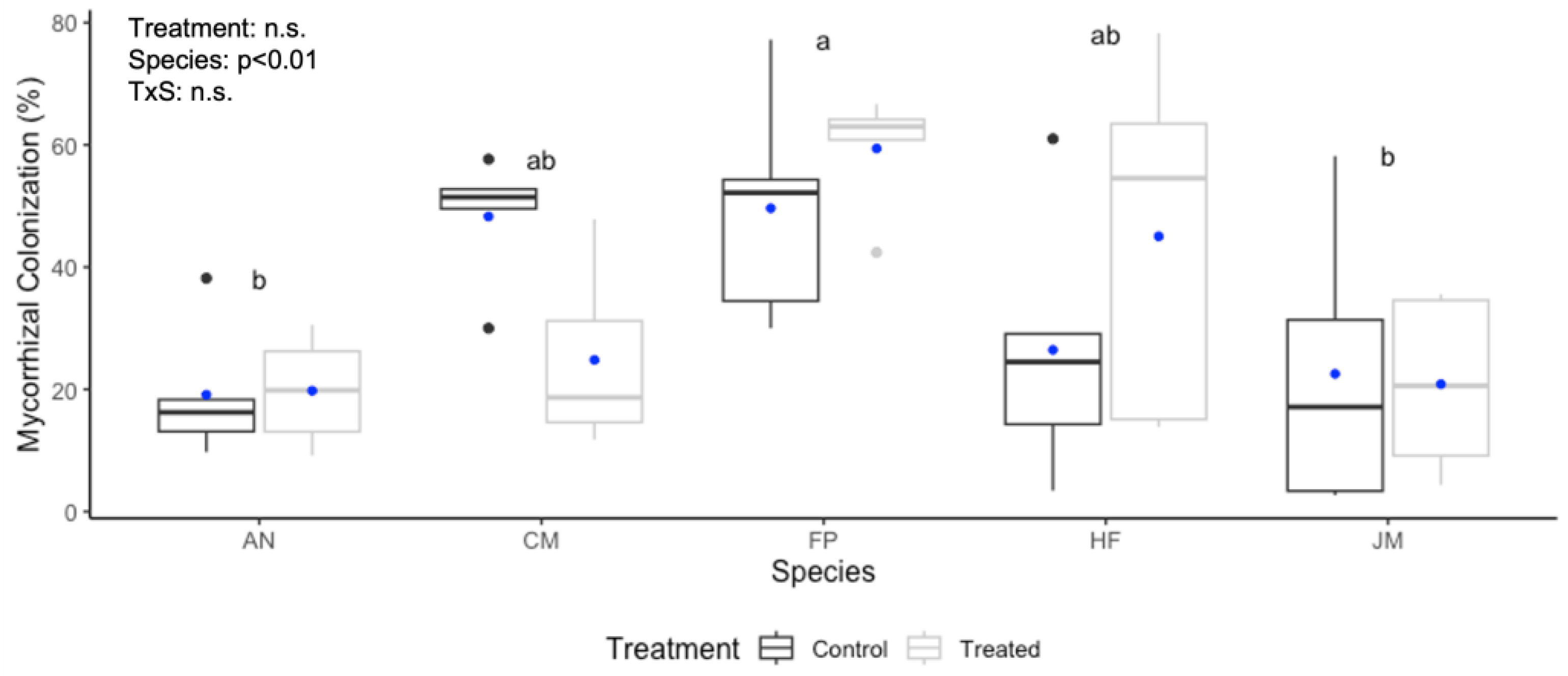
3.2. Root Mycorrhizal Colonization Response to Compost Tea Treatment
3.3. Mycorrhizal Colonization Types
3.4. The Bacterial and Fungal Composition of the Compost Tea
4. Discussion
4.1. Plant Growth Response to Compost Tea
4.2. Mycorrhizal Colonization Response to Compost Tea Treatment
4.3. Mycorrhizal Colonization and Plant Growth in a Container Production System
4.4. Other Effects of Compost Tea and Future Research
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quilty, J.R.; Cattle, S.R. Use and Understanding of Organic Amendments in Australian Agriculture: A Review. Soil Res. 2011, 49, 1–26. [Google Scholar] [CrossRef]
- Kim, M.J.; Shim, C.K.; Kim, Y.K.; Hong, S.J.; Park, J.H.; Han, E.J.; Kim, J.H.; Kim, S.C. Effect of Aerated Compost Tea on the Growth Promotion of Lettuce, Soybean, and Sweet Corn in Organic Cultivation. Plant Pathol. J. 2015, 31, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.K.; Yaseen, T.; Traversa, A.; Ben Kheder, M.; Brunetti, G.; Cocozza, C. Effects of the Main Extraction Parameters on Chemical and Microbial Characteristics of Compost Tea. Waste Manag. 2016, 52, 62–68. [Google Scholar] [CrossRef]
- Pant, A.P.; Radovich, T.J.K.; Hue, N.V.; Paull, R.E. Biochemical Properties of Compost Tea Associated with Compost Quality and Effects on Pak Choi Growth. Sci. Hortic. 2012, 148, 138–146. [Google Scholar] [CrossRef]
- Shaban, H.; Fazeli-Nasab, B.; Alahyari, H.; Alizadeh, G.; Shahpesandi, S. An Overview of the Benefits of Compost Tea on Plant and Soil Structure. Adv. Biores. 2015, 6, 154–158. [Google Scholar]
- Otero, M.; Salcedo, I.; Txarterina, K.; González-Murua, C.; Duñabeitia, M.K. Quality Assessment of Pinus radiata Production under Sustainable Nursery Management Based on Compost Tea. J. Plant Nutr. Soil Sci. 2019, 182, 356–366. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of Organic Amendments Used in Containerized Plant Production: Part 1—Compost-Based Amendments. Sci. Hortic. 2020, 266, 108856. [Google Scholar] [CrossRef]
- Larkin, R.P. Relative Effects of Biological Amendments and Crop Rotations on Soil Microbial Communities and Soilborne Diseases of Potato. Soil Biol. Biochem. 2008, 40, 1341–1351. [Google Scholar] [CrossRef]
- Molineux, C.J.; Connop, S.P.; Gange, A.C. Manipulating Soil Microbial Communities in Extensive Green Roof Substrates. Sci. Total Environ. 2014, 493, 632–638. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.-A.; Sanders, I.R. Mycorrhizal Ecology and Evolution: The Past, the Present, and the Future. New Phytol 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Killham, K. Soil Ecology; Cambridge University Press: Cambridge, MA, USA; New York, NY, USA, 1994; ISBN 978-0-521-43517-8. [Google Scholar]
- Paul, E.A. (Ed.) Soil Microbiology, Ecology, and Biochemistry, 3rd ed.; Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2007; ISBN 978-0-12-546807-7. [Google Scholar]
- Litterick, A.M.; Harrier, L.; Wallace, P.; Watson, C.A.; Wood, M. The Role of Uncomposted Materials, Composts, Manures, and Compost Extracts in Reducing Pest and Disease Incidence and Severity in Sustainable Temperate Agricultural and Horticultural Crop Production—A Review. Crit. Rev. Plant Sci. 2004, 23, 453–479. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Tedersoo, L. Evolutionary History of Mycorrhizal Symbioses and Global Host Plant Diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [Green Version]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T.; Malajczuk, N. Working with Mycorrhizas in Forestry and Agriculture; ACIAR Monograph Series; Australian Centre for International Agricultural Research: Canberra, Australia, 1996; ISBN 978-1-86320-181-0. [Google Scholar]
- Wu, Q.-S.; Xia, R.-X. Arbuscular Mycorrhizal Fungi Influence Growth, Osmotic Adjustment and Photosynthesis of Citrus under Well-Watered and Water Stress Conditions. J. Plant Physiol. 2006, 163, 417–425. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy Metal Stress and Some Mechanisms of Plant Defense Response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [Green Version]
- Leyval, C.; Turnau, K.; Haselwandter, K. Effect of Heavy Metal Pollution on Mycorrhizal Colonization and Function: Physiological, Ecological and Applied Aspects. Mycorrhiza 1997, 7, 139–153. [Google Scholar] [CrossRef]
- Schützendübel, A.; Polie, A. Plant Responses to Abiotic Stresses: Heavy Metal-Induced Oxidative Stress and Protection by Mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef]
- Giri, B.; Kapoor, R.; Mukerji, K.G. Improved Tolerance of Acacia nilotica to Salt Stress by Arbuscular Mycorrhiza, Glomus fasciculatum May Be Partly Related to Elevated K/Na Ratios in Root and Shoot Tissues. Microb. Ecol. 2007, 54, 753–760. [Google Scholar] [CrossRef]
- Hajiboland, R.; Aliasgharzadeh, N.; Laiegh, S.F.; Poschenrieder, C. Colonization with Arbuscular Mycorrhizal Fungi Improves Salinity Tolerance of Tomato (Solanum lycopersicum L.) Plants. Plant Soil 2010, 331, 313–327. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J.L. Building Roots in a Changing Environment: Implications for Root Longevity: REVIEW Building Absorptive Roots and Root Life Span. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Smith, K.; May, P. The Challenge of Urban Soils: Evaluation and Implications for Tree Growth. Landsc. Aust. 1998, 20, 38–42. [Google Scholar]
- Stabler, L.B.; Martin, C.A.; Stutz, J.C. Effect of Urban Expansion on Arbuscular Mycorrhizal Fungal Mediation of Landscape Tree Growth. J. Arboric. 2001, 27, 11. [Google Scholar] [CrossRef]
- Wiseman, P.E.; Wells, C. Soil Inoculum Potential and Arbuscular Mycorrhizal Colonization of Acer rubrum in Forested and Developed Landscapes. J. Arboric. 2005, 31, 7. [Google Scholar]
- Bainard, L.D.; Klironomos, J.N.; Gordon, A.M. The Mycorrhizal Status and Colonization of 26 Tree Species Growing in Urban and Rural Environments. Mycorrhiza 2011, 21, 91–96. [Google Scholar] [CrossRef]
- Savarese, C.; Cozzolino, V.; Verrillo, M.; Vinci, G.; De Martino, A.; Scopa, A.; Piccolo, A. Combination of Humic Biostimulants with a Microbial Inoculum Improves Lettuce Productivity, Nutrient Uptake, and Primary and Secondary Metabolism. Plant Soil 2022, 481, 285–314. [Google Scholar] [CrossRef]
- On, A.; Wong, F.; Ko, Q.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Antifungal Effects of Compost Tea Microorganisms on Tomato Pathogens. Biol. Control 2015, 80, 63–69. [Google Scholar] [CrossRef]
- Edenborn, S.L.; Johnson, L.M.K.; Edenborn, H.M.; Albarran-Jack, M.R.; Demetrion, L.D. Amendment of a Hardwood Biochar with Compost Tea: Effects on Plant Growth, Insect Damage and the Functional Diversity of Soil Microbial Communities. Biol. Agric. Hortic. 2018, 34, 88–106. [Google Scholar] [CrossRef]
- Radin, A.M.; Warman, P.R. Effect of Municipal Solid Waste Compost and Compost Tea as Fertility Amendments on Growth and Tissue Element Concentration in Container-Grown Tomato. Commun. Soil Sci. Plant Anal. 2011, 42, 1349–1362. [Google Scholar] [CrossRef]
- Ou-Zine, M.; El Hilali, R.; Haggoud, A.; Achbani, E.H.; Bouamri, R. Effects and Relationships of Compost Dose and Organic Additives on Compost Tea Properties, Efficacy Against Fusarium Oxysporum and Potential Effect on Endomycorrhization and Growth of Zea Mays L. Waste Biomass Valor 2022, 13, 4431–4445. [Google Scholar] [CrossRef]
- Sax, M.S.; Scharenbroch, B.C. Assessing Alternative Organic Amendments as Horticultural Substrates for Growing Trees in Containers. J. Environ. Hortic. 2017, 35, 66–78. [Google Scholar] [CrossRef]
- Atlas of Living Australia. Available online: https://www.ala.org.au (accessed on 13 May 2023).
- Soudzilovskaia, N.A.; Vaessen, S.; Barcelo, M.; He, J.; Rahimlou, S.; Abarenkov, K.; Brundrett, M.C.; Gomes, S.I.F.; Merckx, V.; Tedersoo, L. FungalRoot: Global Online Database of Plant Mycorrhizal Associations. New Phytol. 2020, 227, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Lovelock, C.E.; Miller, R. Heterogeneity in Inoculum Potential and Effectiveness of Arbuscular Mycorrhizal Fungi. Ecology 2002, 83, 823–832. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and Vinegar, a Simple Staining Technique for Arbuscular-Mycorrhizal Fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Zinger, L.; Nilsson, R.H.; Kennedy, P.G.; Yang, T.; Anslan, S.; Mikryukov, V. Best Practices in Metabarcoding of Fungi: From Experimental Design to Results. Mol. Ecol. 2022, 31, 2769–2795. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.M.; Jusino, M.A.; Banik, M.T.; Lindner, D.L. Non-Biological Synthetic Spike-in Controls and the AMPtk Software Pipeline Improve Mycobiome Data. PeerJ 2018, 6, e4925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Põlme, S.; Abarenkov, K.; Henrik Nilsson, R.; Lindahl, B.D.; Clemmensen, K.E.; Kauserud, H.; Nguyen, N.; Kjøller, R.; Bates, S.T.; Baldrian, P.; et al. FungalTraits: A User-Friendly Traits Database of Fungi and Fungus-like Stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Fusco, V.; Quero, G.M.; Cho, G.-S.; Kabisch, J.; Meske, D.; Neve, H.; Bockelmann, W.; Franz, C.M.A.P. The Genus Weissella: Taxonomy, Ecology and Biotechnological Potential. Front. Microbiol. 2015, 6, 155. [Google Scholar] [CrossRef] [Green Version]
- Finley, R.L.; Collignon, P.; Larsson, D.G.J.; McEwen, S.A.; Li, X.-Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The Scourge of Antibiotic Resistance: The Important Role of the Environment. Clin. Infect. Dis. 2013, 57, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Yoon, E.-J.; Goussard, S.; Touchon, M.; Krizova, L.; Cerqueira, G.; Murphy, C.; Lambert, T.; Grillot-Courvalin, C.; Nemec, A.; Courvalin, P. Origin in Acinetobacter guillouiae and Dissemination of the Aminoglycoside-Modifying Enzyme Aph(3′)-VI. mBio 2014, 5, e01972-14. [Google Scholar] [CrossRef] [Green Version]
- de Lima-Morales, D.; Chaves-Moreno, D.; Wos-Oxley, M.L.; Jáuregui, R.; Vilchez-Vargas, R.; Pieper, D.H. Degradation of Benzene by Pseudomonas veronii 1YdBTEX2 and 1YB2 Is Catalyzed by Enzymes Encoded in Distinct Catabolism Gene Clusters. Appl. Environ. Microbiol. 2016, 82, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil Beneficial Bacteria and Their Role in Plant Growth Promotion: A Review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Domann, E.; Hong, G.; Imirzalioglu, C.; Turschner, S.; Kuhle, J.; Watzel, C.; Hain, T.; Hossain, H.; Chakraborty, T. Culture-Independent Identification of Pathogenic Bacteria and Polymicrobial Infections in the Genitourinary Tract of Renal Transplant Recipients. J. Clin. Microbiol. 2003, 41, 5500–5510. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S. Demonstration of Safety of Probiotics—A Review. Int. J. Food Microbiol. 1998, 44, 93–106. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.-A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of Forest Soil Respiration in Response to Nitrogen Deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Carballo, T.; Gil, M.V.; Calvo, L.F.; Morán, A. The Influence of Aeration System, Temperature and Compost Origin on the Phytotoxicity of Compost Tea. Compos. Sci. Util. 2009, 17, 127–139. [Google Scholar] [CrossRef]
- Mestre, M.C.; Pastorino, M.J.; Aparicio, A.G.; Fontenla, S.B. Natives Helping Foreigners?: The Effect of Inoculation of Poplar with Patagonian Beneficial Microorganisms. J. Soil Sci. Plant Nutr. 2017, 17, 1028–1039. [Google Scholar] [CrossRef] [Green Version]
- Zaouchi, Y.; Bahri, N.B.; Rezgui, S.; Bettaieb, T. Effects of Arbuscular Mycorrhizal Inoculation and Fertilization on Mycorrhizal Statute of Jacaranda mimosifolia D.Don Cultivated in Nurseries. Comptes Rendus Biol. 2013, 336, 493–499. [Google Scholar] [CrossRef]
- Tomao, A.; Antonio Bonet, J.; Castaño, C.; de-Miguel, S. How Does Forest Management Affect Fungal Diversity and Community Composition? Current Knowledge and Future Perspectives for the Conservation of Forest Fungi. For. Ecol. Manag. 2020, 457, 117678. [Google Scholar] [CrossRef]
- Adams, F.; Reddell, P.; Webb, M.J.; Shipton, W.A. Arbuscular Mycorrhizas and Ectomycorrhizas on Eucalyptus grandis (Myrtaceae) Trees and Seedlings in Native Forests of Tropical North-Eastern Australia. Aust. J. Bot. 2006, 54, 271. [Google Scholar] [CrossRef]
- Estaún, V.; Camprubí, A.; Calvet, C.; Pinochet, J. Nursery and Field Response of Olive Trees Inoculated with Two Arbuscular Mycorrhizal Fungi, Glomus Intraradices and Glomus Mosseae. J. Am. Soc. Hortic. Sci. 2003, 128, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.J.; Nicholl, P.A.; Hicklenton, P.R. VA Mycorrhizal Inoculation of Landscape Trees and Shrubs Growing under High Fertility Conditions. J. Environ. Hortic. 1993, 11, 64–71. [Google Scholar] [CrossRef]
- Graham, J.H.; Eissenstat, D.M. Field Evidence for the Carbon Cost of Citrus Mycorrhizas. New Phytol. 1998, 140, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Albornoz, F.E.; Dixon, K.W.; Lambers, H. Revisiting Mycorrhizal Dogmas: Are Mycorrhizas Really Functioning as They Are Widely Believed to Do? Soil Ecol. Lett. 2021, 3, 73–82. [Google Scholar] [CrossRef]
- Sylvia, D.; Alagely, A.; Kent, D.; Mecklenburg, R. Mycorrhizae of Landscape Trees Produced in Raised Beds and Containers. J. Arboric. 1998, 24, 308–315. [Google Scholar] [CrossRef]
- Corkidi, L.; Allen, E.B.; Merhaut, D.; Allen, M.F.; Downer, J.; Bohn, J.; Evans, M. Assessing the Infectivity of Commercial Mycorrhizal Inoculants in Plant Nursery Conditions. J. Environ. Hortic. 2004, 22, 149–154. [Google Scholar] [CrossRef]
- Fini, A.; Frangi, P.; Amoroso, G.; Piatti, R.; Faoro, M.; Bellasio, C.; Ferrini, F. Effect of Controlled Inoculation with Specific Mycorrhizal Fungi from the Urban Environment on Growth and Physiology of Containerized Shade Tree Species Growing under Different Water Regimes. Mycorrhiza 2011, 21, 703–719. [Google Scholar] [CrossRef]
- Wiseman, P.E.; Wells, C.E. Arbuscular Mycorrhizal Inoculation Affects Root Development of Acer and Magnolia Species. J. Environ. Hortic. 2009, 27, 70–79. [Google Scholar] [CrossRef]
- Janos, D.P. Plant Responsiveness to Mycorrhizas Differs from Dependence upon Mycorrhizas. Mycorrhiza 2007, 17, 75–91. [Google Scholar] [CrossRef]
- Prescott, C.E. Sinks for Plant Surplus Carbon Explain Several Ecological Phenomena. Plant Soil 2022, 476, 689–698. [Google Scholar] [CrossRef]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A Meta-Analysis of Context-Dependency in Plant Response to Inoculation with Mycorrhizal Fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ramos, J.C.; Cale, J.A.; Cahill, J.F.; Erbilgin, N.; Karst, J. Soil Transfers from Intact to Disturbed Boreal Forests Neither Alter Ectomycorrhizal Fungal Communities nor Improve Pine Seedling Performance. J. Appl. Ecol. 2022, 59, 2430–2439. [Google Scholar] [CrossRef]
- Wasyliw, J.; Fellrath, E.G.; Pec, G.J.; Cale, J.A.; Franklin, J.; Thomasson, C.; Erbilgin, N.; Karst, J. Soil Inoculation of Lodgepole Pine Seedlings Alters Root-associated Fungal Communities but Does Not Improve Seedling Performance in Beetle-killed Pine Stands. Restor. Ecol. 2023, 31, e13663. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredient | Amount (by Volume) | Additional Information |
|---|---|---|
| Leaf trimmings | 10%–15% | Green waste on site |
| Weeds | 20%–25% | Green waste on site |
| Grass clippings | 5%–10% | Green waste on site |
| Dynamic lifter | 5% | Commercial product |
| Horse manure | 5% | - |
| Seamungus | 5% | Commercial product |
| Rock dust | 2% | - |
| Wood chips | 25%–30% | - |
| Root balls | 10%–15% | From discarded mixed tree species on site |
| Mature compost | 5% | From an old compost pile made previously in the same way |
| Family | Species | Type | Origin | Mycorrhizal Type |
|---|---|---|---|---|
| Sapindaceae | Acer negundo L. | Deciduous | Exotic | VAM |
| Myrtaceae | Corymbia maculata (Hook.) K.D. Hill & L.A.S. Johnson | Evergreen | Native | Dual (VAM/ECM) |
| Moraceae | Ficus platypoda (Miq.) A. Cunn. ex Miq. | Evergreen | Native | VAM |
| Pittosporaceae | Hymenosporum flavum F. Muell. | Evergreen | Native | VAM |
| Bignoniaceae | Jacaranda mimosifolia D. Don | Deciduous | Exotic | VAM |
| ID | Primary Lifestyle | Secondary Lifestyle | Relative Abund. (%) | Additional Information |
|---|---|---|---|---|
| Barnettozyma californica | Saprotroph (various substrates) | 25.07 | Yeast, sugar-rich substrates | |
| Mortierella indohii | Soil saprotroph | Root-associated | 14.53 | Filamentous mycelium |
| Penicillium sp. | Saprotroph (various substrates) | Foliar endophyte | 11.82 | Mould, some species are toxin-producing, animal parasites or mycoparasites |
| Chaetomium piluliferum | Litter saprotroph | Foliar endophyte | 9.44 | Soft rot |
| Trichothecium roseum | Plant pathogen | Litter saprotroph | 8.60 | Filamentous mycelium |
| Thermomyces sp. | Soil saprotroph | Litter saprotroph | 5.64 | Mould, potential plant pathogenicity |
| Unknown Trichosporonaceae | – | – | 4.16 | – |
| Rhodotorula sp. | Saprotroph (various substrates) | Foliar endophyte | 3.89 | Yeast |
| Candida sp. | Saprotroph (various substrates) | 3.62 | Yeast, sugar-rich substrates | |
| Byssochlamys sp. | Saprotroph (various substrates) | 3.13 | Mould, some species are food spoilage agents | |
| Mortierella reticulata | Soil saprotroph | Root-associated | 2.52 | Filamentous mycelium |
| Unknown Fungi | – | – | 2.22 | – |
| Fusarium oxysporum | Plant pathogen | Litter saprotroph | 1.24 | Soft rot |
| Mortierella sp. | Soil saprotroph | Root-associated | 1.00 | Filamentous mycelium |
| Unknown Nectriaceae | – | – | 0.76 | – |
| Aspergillus fumigatus | Saprotroph (various substrates) | Foliar endophyte | 0.67 | Mould |
| Cutaneotrichosporon sp. | Animal parasite | Animal decomposer | 0.58 | Yeast |
| Wickerhamomyces anomalus | Litter saprotroph | Saprotroph | 0.57 | Yeast, sugar-rich substrates |
| Acremonium sp. | Saprotroph (various substrates) | Foliar endophyte | 0.56 | Soft rot, potential plant pathogenicity |
| ID | Function | Relative Abund. (%) | Additional Information |
|---|---|---|---|
| Weissella sp. | Probiotic/pathogen | 25.60 | Prolific in environment, probiotic or pathogenic [44] |
| Acinetobacter sp. | Pathogen/degrader | 22.27 | Can be pathogenic to humans [45] |
| Acinetobacter guillouiae | Unknown | 11.05 | An environmental species [46] |
| Pseudomonas sp. | Pathogen/degrader | 10.30 | Commonly exist in soil and can be plant pathogens [11] |
| Unknown Enterobacteriaceae | Unknown | 4.15 | |
| Pseudomonas veronii | Degrader | 3.52 | A bioremediation of contaminated soils [47] |
| Acinetobacter sp. | Pathogen/degrader | 3.50 | Can be pathogenic to human [45] |
| Enterobacter sp. | Probiotic | 3.36 | Might be nitrogen fixing bacteria [48] |
| Acetinobacter sp. | Pathogen/degrader | 3.32 | Can be pathogenic to human [45] |
| Pseudomonas sp. | Pathogen/degrader | 3.22 | Commonly exist in soil and can be plant pathogens [11] |
| Hafnia sp. | Pathogen | 2.14 | Opportunistic pathogen |
| Rahnella aquatilis | Pathogen | 1.67 | Opportunistic pathogen [49] |
| Serratia sp. | Pathogen | 1.67 | Opportunistic pathogen |
| Bacillus sp. | Unclear | 1.47 | Commonly exist in soil and might fix nitrogen [11] |
| Lactococcus sp. | Lactic acid bacteria | 1.04 | They are generally safe and produce lactic acid [50] |
| Arthrobacter sp. | Opportunistic pathogen | 0.87 | Commonly exists in soil [11] |
| Acetinobacter sp. | Pathogen/degrader | 0.86 | Can be pathogenic to human [45] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, D.; Livesley, S.J.; Arndt, S.K.; Truong, C.; Miller, R.E. The Use of Compost Tea in a Containerized Urban Tree Nursery Shows No Evident Benefits to Tree Growth or Mycorrhizal Colonization. Forests 2023, 14, 1195. https://doi.org/10.3390/f14061195
Du D, Livesley SJ, Arndt SK, Truong C, Miller RE. The Use of Compost Tea in a Containerized Urban Tree Nursery Shows No Evident Benefits to Tree Growth or Mycorrhizal Colonization. Forests. 2023; 14(6):1195. https://doi.org/10.3390/f14061195
Chicago/Turabian StyleDu, Dan, Stephen J. Livesley, Stefan K. Arndt, Camille Truong, and Rebecca E. Miller. 2023. "The Use of Compost Tea in a Containerized Urban Tree Nursery Shows No Evident Benefits to Tree Growth or Mycorrhizal Colonization" Forests 14, no. 6: 1195. https://doi.org/10.3390/f14061195