


Plant–Soil Feedback of Companion Species during Grassland Community Succession
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.1.1. Plant, Seed, and Soil Collection
2.1.2. Plant–Soil Feedback (PSF) Experiment
2.1.3. Plant–Soil Sampling
2.2. Library Analysis
2.3. Statistical Analysis
3. Results
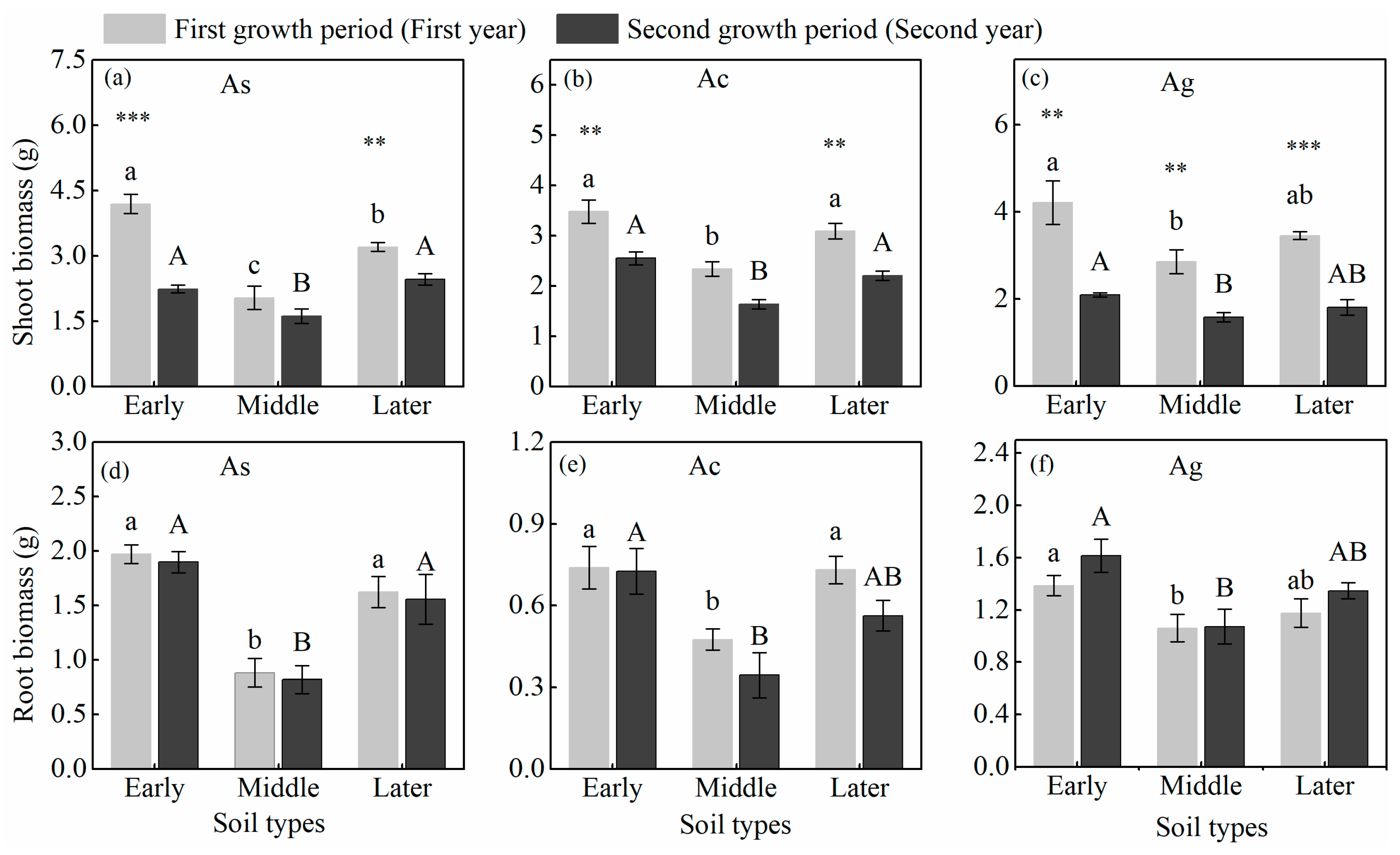
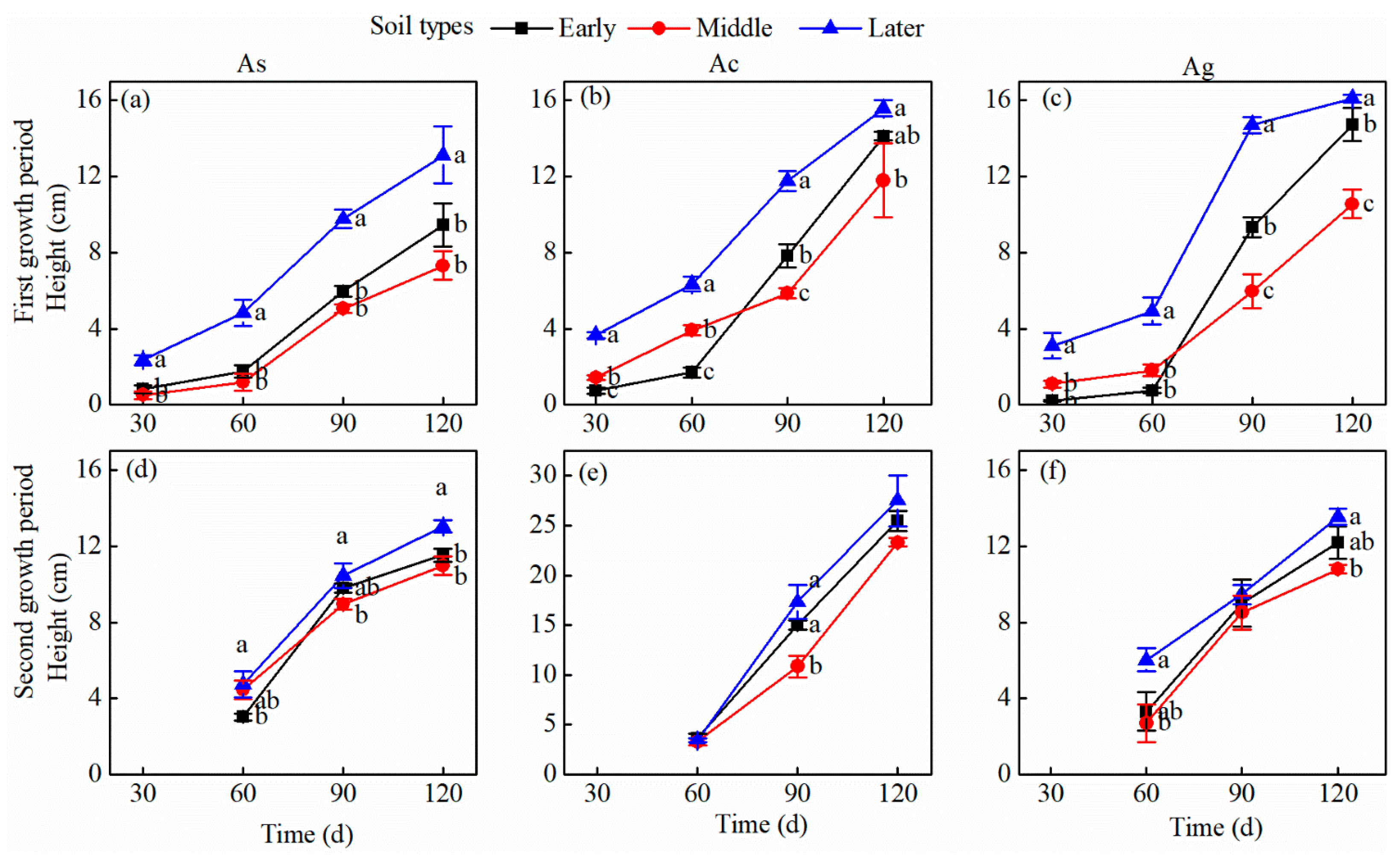
3.1. Plant Biomass, PSF Index, and Plant Height
3.2. Carbon, Nitrogen, and Phosphorus Characteristics of Plants
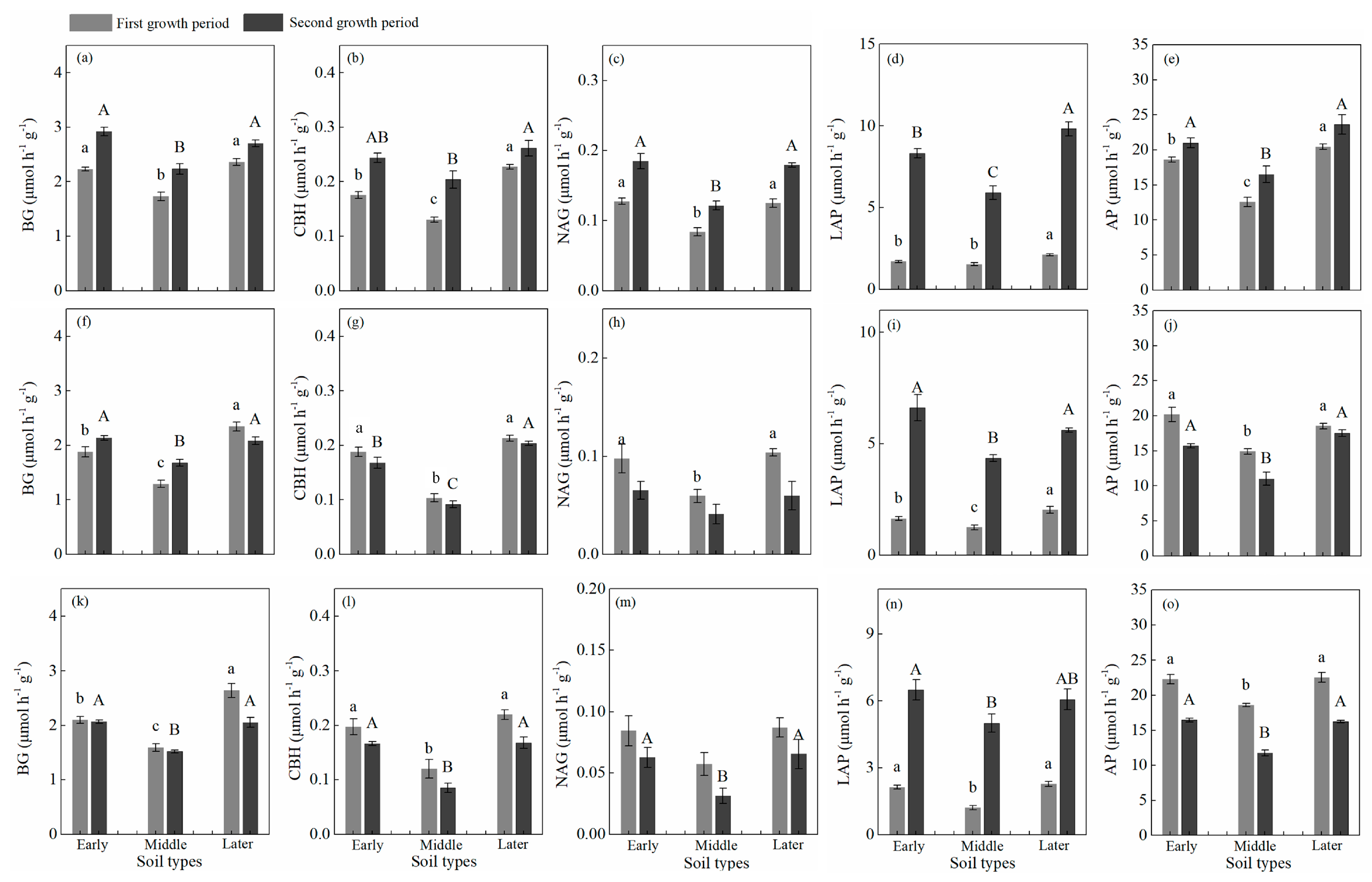
3.3. Soil Carbon, Nitrogen, and Phosphorus, Enzyme Activity, and Microbial Biomass
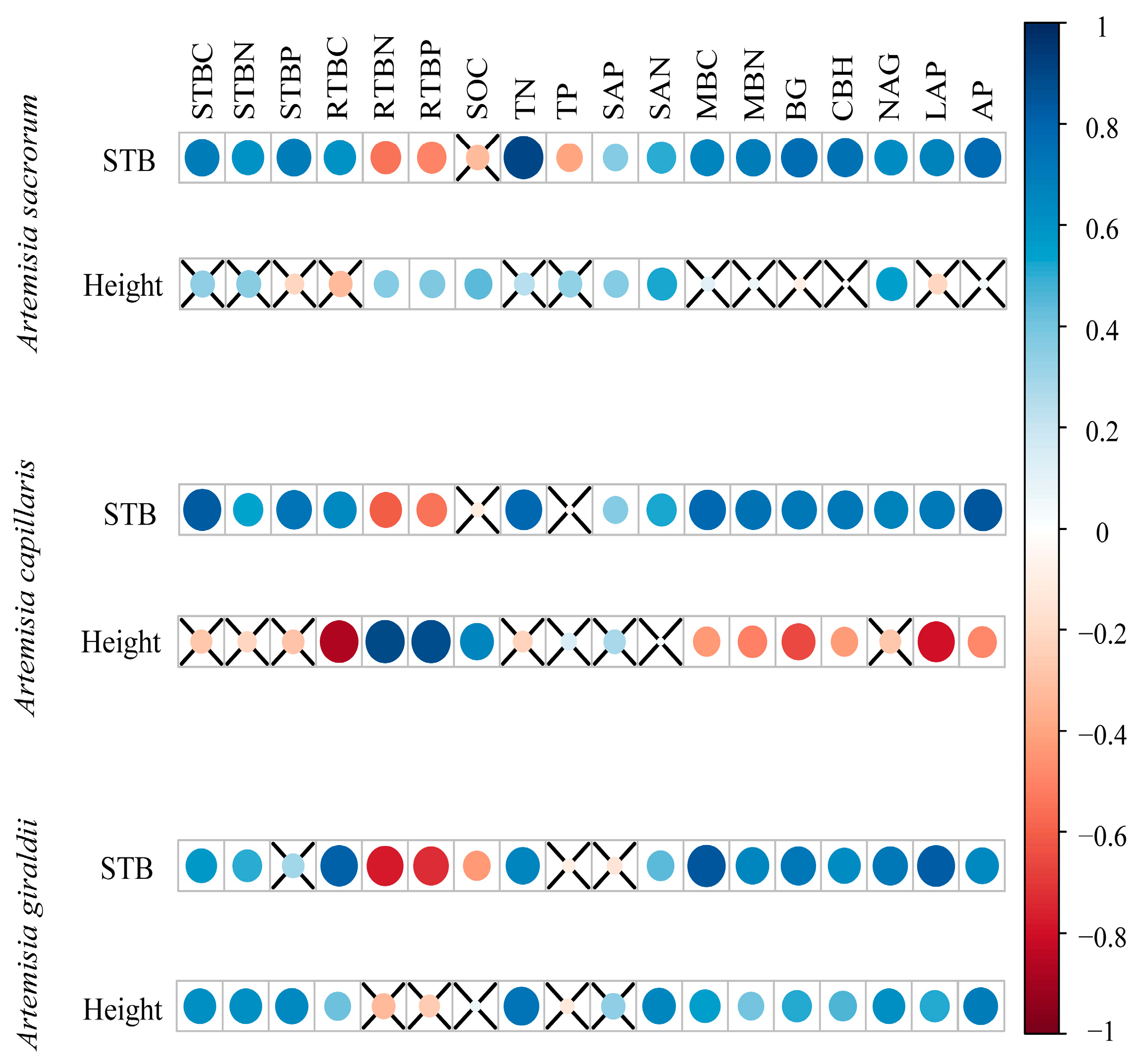
3.4. Correlation Analysis between Plant–Soil–Enzyme Activities and Plant Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marasco, A.; Giannino, F.; Iuorio, A. Modelling competitive interactions and plant-soil feedback in vegetation dynamics. Ric. Mat. 2020, 69, 553–577. [Google Scholar] [CrossRef]
- Zhang, J.; Ai, Z.; Xu, H.; Liu, H.; Wang, G.; Deng, L.; Liu, G.; Xue, S. Plant-microbial feedback in secondary succession of semiarid grasslands. Sci. Total Environ. 2021, 760, 143389. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ai, Z.; Qu, Q.; Wang, M.; Liu, G.; Xue, S. Invasibility and recoverability of a plant community following invasion depend on its successional stages. Soil Ecol. Lett. 2022, 4, 171–185. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Li, P.; Guo, Z.; Wulan, E.; Xue, S. Stocks and Stoichiometry of Soil Organic Carbon, Total Nitrogen, and Total Phosphorus after Vegetation Restoration in the Loess Hilly Region, China. Forests 2019, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dang, Z.; Tian, F.; Wang, D.; Wu, G. Soil organic carbon and inorganic carbon accumulation along a 30-year grassland restoration chronosequence in semi-arid regions (China). Land Degrad. Dev. 2017, 28, 189–198. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Chen, Y.; Liu, G.; Xue, S. Responses of soil enzyme activity and soil organic carbon stability over time after cropland abandonment in different vegetation zones of the Loess Plateau of China. Catena 2021, 196, 104812. [Google Scholar] [CrossRef]
- Deng, L.; Liu, G.; Shangguan, Z. Land-use conversion and changing soil carbon stocks in China’s ‘Grain-for-Green’ Program: A synthesis. Glob. Chang. Biol. 2014, 20, 3544–3556. [Google Scholar] [CrossRef]
- Frouz, J.; Toyota, A.; Mudrak, O.; Jilkova, V.; Filipova, A.; Cajthaml, T. Effects of soil substrate quality, microbial diversity and community composition on the plant community during primary succession. Soil Biol. Biochem. 2016, 99, 75–84. [Google Scholar] [CrossRef]
- Bever, J.D.; Westover, K.M.; Antonovics, J. Incorporating the soil community into plant population dynamics: The utility of the feedback approach. J. Ecol. 1997, 85, 561–573. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Ravit, B.; Elgersma, K. Feedback in the plant-soil system. Annu. Rev. Env. Resour. 2005, 30, 75–115. [Google Scholar] [CrossRef] [Green Version]
- Bezemer, T.M.; van der Putten, W.H.; Martens, H.; van de Voorde, T.F.J.; Mulder, P.P.J.; Kostenko, O. Above- and below-ground herbivory effects on below-ground plant-fungus interactions and plant-soil feedback responses. J. Ecol. 2013, 101, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Heinze, J.; Wacker, A.; Kulmatiski, A. Plant-soil feedback effects altered by aboveground herbivory explain plant species abundance in the landscape. Ecology 2020, 101, e03023. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Van Dijk, C.; Peters, B.A.M. Plant-specific soil-borne diseases contribute to succession in foredune vegetation. Nature 1993, 362, 53–56. [Google Scholar] [CrossRef]
- Jing, J.; Bezemer, T.M.; van der Putten, W.H. Complementarity and selection effects in early and mid-successional plant communities are differentially affected by plant-soil feedback. J. Ecol. 2015, 103, 641–647. [Google Scholar] [CrossRef]
- Kardol, P.; Bezemer, T.M.; van der Putten, W.H. Temporal variation in plant-soil feedback controls succession. Ecol. Lett. 2006, 9, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; van Kleunen, M. Common alien plants are more competitive than rare natives but not than common natives. Ecol. Lett. 2019, 22, 1378–1386. [Google Scholar] [CrossRef]
- Crawford, K.M.; Bauer, J.T.; Comita, L.S.; Eppinga, M.B.; Johnson, D.J.; Mangan, S.A.; Queenborough, S.A.; Strand, A.E.; Suding, K.N.; Umbanhowar, J.; et al. When and where plant-soil feedback may promote plant coexistence: A meta-analysis. Ecol. Lett. 2019, 22, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, X.P.; Qu, Y.B.; Wu, M.; Wang, J.L.; Wang, Z.L.; Wang, S.Q.; Zhao, N.X.; Gao, Y.B. Comparing the effects of companion species diversity and the dominant species (Stipa grandis) genotypic diversity on the biomass explained by plant functional trait. Ecol. Eng. 2019, 136, 17–22. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen-Total. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; ASA, SSSA: Madison, WI, USA, 1983; pp. 595–624. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 2. Chemical and Microbial Properties, 2nd ed.; Page, A.L., Ed.; ASA, SSSA: Madison, WI, USA, 1983; pp. 539–579. [Google Scholar]
- Schade, J.D.; Kyle, M.; Hobbie, S.E.; Fagan, W.F.; Elser, J.J. Stoichiometric tracking of soil nutrients by a desert insect herbivore. Ecol. Lett. 2003, 6, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis, Part 2. Chemical and Microbial Properties, 2nd ed.; Page, A.L., Ed.; ASA, SSSA: Madison, WI, USA, 1983; pp. 403–430. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Baxendal, C.; Orwin, K.H.; Poly, F.; Pommier, T.; Bardgett, R.D. Are plant-soil feedback responses explained by plant traits. New Phytol. 2014, 204, 408–423. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Xu, H.; Wang, M.; Liu, G.; Xue, S. Plant-soil feedback plays an important role in the progression of plant community succession. J. Plant Nutr. Soil Sci. 2023, 186, 169–176. [Google Scholar] [CrossRef]
- Martinez-Luscher, J.; Kurtural, S.K. Same season and carry-over effects of source-sink adjustments on grapevine yields and non-structural carbohydrates. Front. Plant Sci. 2021, 12, 695319. [Google Scholar] [CrossRef] [PubMed]
- Perkovich, C.; Ward, D. Aboveground herbivory causes belowground changes in twelve oak Quercus species: A phylogenetic analysis of root biomass and non-structural carbohydrate storage. Oikos 2021, 130, 1797–1812. [Google Scholar] [CrossRef]
- Mahaming, A.R.; Mills, A.A.S.; Adl, S.M. Soil community changes during secondary succession to naturalized grasslands. Appl. Soil Ecol. 2009, 41, 137–147. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.; Li, P.; Li, Q.; Xue, S. Ecoenzymatic stoichiometry and microbial nutrient limitation during secondary succession of natural grassland on the Loess Plateau, China. Soil Tillage Res. 2020, 200, 104605. [Google Scholar] [CrossRef]
- Harrison, K.A.; Bardgett, R.D. Influence of plant species and soil conditions on plant-soil feedback in mixed grassland communities. J. Ecol. 2010, 98, 384–395. [Google Scholar] [CrossRef]
- Bennett, J.A.; Klironomos, J. Mechanisms of plant-soil feedback: Interactions among biotic and abiotic drivers. New Phytol. 2019, 222, 91–96. [Google Scholar] [CrossRef] [Green Version]
- van de Voorde, T.F.J.; van der Putten, W.H.; Bezemer, T.M. Intra- and interspecific plant-soil interactions, soil legacies and priority effects during old-field succession. J. Ecol. 2011, 99, 945–953. [Google Scholar] [CrossRef]
- De Long, J.R.; Heinen, R.; Jongen, R.; Hannula, S.E.; Huberty, M.; Kielak, A.M.; Steinauer, K.; Martijn Bezemer, T. How plant-soil feedbacks influence the next generation of plants. Ecol. Res. 2021, 36, 32–44. [Google Scholar] [CrossRef]
- Bennett, J.A.; Maherali, H.; Reinhart, K.O.; Lekberg, Y.; Hart, M.M.; Klironomos, J. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 2017, 355, 181–184. [Google Scholar] [CrossRef]
- Fujii, K.; Shibata, M.; Kitajima, K.; Ichie, T.; Kitayama, K.; Turner, B.L. Plant-soil interactions maintain biodiversity and functions of tropical forest ecosystems. Ecol. Res. 2018, 33, 149–160. [Google Scholar] [CrossRef]
- González-López, O.; Ruano-Rosa, D. Root exudates, a key factor in the plant-bacteria interaction mechanisms. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 111–121. [Google Scholar]
- Bray, S.R.; Kitajima, K.; Sylvia, D.M. Mycorrhizae differentially alter growth, physiology, and competitive ability of an invasive shrub. Ecol. Appl. 2003, 13, 565–574. [Google Scholar] [CrossRef]
- Zheng, L.; Chen, H.; Wang, Y.; Mao, Q.; Zheng, M.; Su, Y.; Xiao, K.; Wang, K.; Li, D. Responses of soil microbial resource limitation to multiple fertilization strategies. Soil Tillage Res. 2020, 196, 104474. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, W.; Zhong, Z.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass, diversity, and compositions to altitudinal gradients depend on plant and soil characteristics. Sci. Total Environ. 2018, 610–611, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Ai, Z.; Li, J.; Li, X.; Zhang, J.; Liu, H.; Xu, H.; Liu, G.; Xue, S. Plant traits variably respond to plant–soil interactions during secondary succession on the Loess Plateau. Forests 2023, 14, 726. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Period | Species | Soil Types | STBC (g kg−1) | STBN (g kg−1) | STBP (g kg−1) |
|---|---|---|---|---|---|
| First | A. sacrorum | Early | 434.85 ± 13.02 a *** | 9.26 ± 0.35 a ** | 2.14 ± 0.07 b *** |
| Middle | 363.60 ± 5.90 b ** | 7.51 ± 0.44 b | 1.81 ± 0.05 c *** | ||
| Late | 428.20 ± 3.91 a *** | 9.54 ± 0.26 a ** | 2.45 ± 0.05 a *** | ||
| A. capillaris | Early | 438.20 ± 7.65 a *** | 9.80 ± 0.76 b ** | 1.74 ± 0.04 a *** | |
| Middle | 323.46 ± 5.55 c | 7.11 ± 0.07 c * | 0.66 ± 0.22 b | ||
| Late | 391.98 ± 6.10 b *** | 11.72 ± 0.28 a ** | 1.67 ± 0.04 a *** | ||
| A. giraldii | Early | 433.92 ± 3.67 a ** | 9.80 ± 0.31 ab ** | 1.12 ± 0.09 b | |
| Middle | 403.79 ± 7.15 c * | 8.34 ± 0.40 b * | 0.49 ± 0.04 c ** | ||
| Late | 452.30 ± 3.01 a *** | 10.15 ± 0.48 a | 1.81 ± 0.15 a ** | ||
| Second | A. sacrorum | Early | 407.65 ± 4.46 A | 8.70 ± 0.25 A | 1.10 ± 0.1 A |
| Middle | 325.11 ± 8.87 C | 7.01 ± 0.49 B | 0.76 ± 0.06 B | ||
| Late | 381.38 ± 4.53 B | 8.34 ± 0.10 A | 1.24 ± 0.05 A | ||
| A. capillaris | Early | 377.59 ± 3.80 A | 8.02 ± 0.23 B | 1.08 ± 0.04 A | |
| Middle | 305.00 ± 6.97 C | 6.57 ± 0.21 C | 0.67 ± 0.02 C | ||
| Late | 329.32 ± 4.56 B | 9.86 ± 0.23 A | 0.94 ± 0.03 B | ||
| A. giraldii | Early | 416.95 ± 2.53 A | 8.52 ± 0.13 B | 1.11 ± 0.02 A | |
| Middle | 376.67 ± 7.16 B | 7.34 ± 0.12 C | 0.72 ± 0.01 B | ||
| Late | 411.87 ± 6.53 A | 9.21 ± 0.08 A | 1.05 ± 0.04 A |
| Growth Period | Species | Soil Types | RTBC (g kg−1) | RTBN (g kg−1) | RTBP (g kg−1) |
|---|---|---|---|---|---|
| First | A. sacrorum | Early | 367.99 ± 1.08 b *** | 8.78 ± 0.25 a *** | 0.57 ± 0.04 a *** |
| Middle | 323.81 ± 5.24 c *** | 5.54 ± 0.46 b *** | 0.31 ± 0.05 b *** | ||
| Late | 401.89 ± 9.25 a *** | 7.79 ± 0.26 a *** | 0.58 ± 0.00 a *** | ||
| A. capillaris | Early | 370.16 ± 1.66 a *** | 7.80 ± 0.06 a *** | 0.55 ± 0.00 a *** | |
| Middle | 345.42 ± 7.07 b *** | 5.94 ± 0.4 b *** | 0.45 ± 0.00 b *** | ||
| Late | 376.50 ± 8.64 a *** | 7.96 ± 0.2 a *** | 0.59 ± 0.01 a *** | ||
| A. giraldii | Early | 366.62 ± 7.18 a *** | 9.02 ± 0.77 a *** | 0.61 ± 0.08 *** | |
| Middle | 318.49 ± 0.94 b *** | 6.05 ± 0.08 b *** | 0.43 ± 0.01 *** | ||
| Late | 362.53 ± 10.43 a *** | 8.06 ± 0.35 ab *** | 0.51 ± 0.02 *** | ||
| Second | A. sacrorum | Early | 348.67 ± 3.39 A | 7.10 ± 0.25 A | 0.46 ± 0.03 |
| Middle | 312.33 ± 4.58 B | 5.38 ± 0.15 B | 0.42 ± 0.03 | ||
| Late | 334.42 ± 5.01 A | 6.63 ± 0.17 A | 0.44 ± 0.01 | ||
| A. capillaris | Early | 354.78 ± 7.94 A | 6.74 ± 0.34 A | 1.07 ± 0.02 A | |
| Middle | 327.39 ± 2.95 B | 5.31 ± 0.13 B | 0.69 ± 0.01 C | ||
| Late | 351.72 ± 4.34 A | 6.69 ± 0.12 A | 0.89 ± 0.04 B | ||
| A. giraldii | Early | 334.69 ± 4.02 A | 8.01 ± 0.15 A | 1.12 ± 0.03 A | |
| Middle | 307.35 ± 7.27 B | 5.62 ± 0.17 C | 0.86 ± 0.01 B | ||
| Late | 338.28 ± 2.36 A | 6.95 ± 0.17 B | 0.86 ± 0.02 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, L.; Huang, L.; Zhou, H.; Xue, S.; Wang, M.; Xu, H. Plant–Soil Feedback of Companion Species during Grassland Community Succession. Forests 2023, 14, 1634. https://doi.org/10.3390/f14081634
Zhang L, Zhang L, Huang L, Zhou H, Xue S, Wang M, Xu H. Plant–Soil Feedback of Companion Species during Grassland Community Succession. Forests. 2023; 14(8):1634. https://doi.org/10.3390/f14081634
Chicago/Turabian StyleZhang, Li, Linhui Zhang, Lulu Huang, Huiling Zhou, Sha Xue, Minggang Wang, and Hongwei Xu. 2023. "Plant–Soil Feedback of Companion Species during Grassland Community Succession" Forests 14, no. 8: 1634. https://doi.org/10.3390/f14081634