
Ecoenzymatic Stoichiometry in the Rhizosphere and Bulk Soil of a Larix principis-rupprechtii Plantation in North China
Abstract
:1. Introduction
2. Methods
2.1. Study Sites
2.2. Soil Sampling
2.3. Measurements of Soil Properties
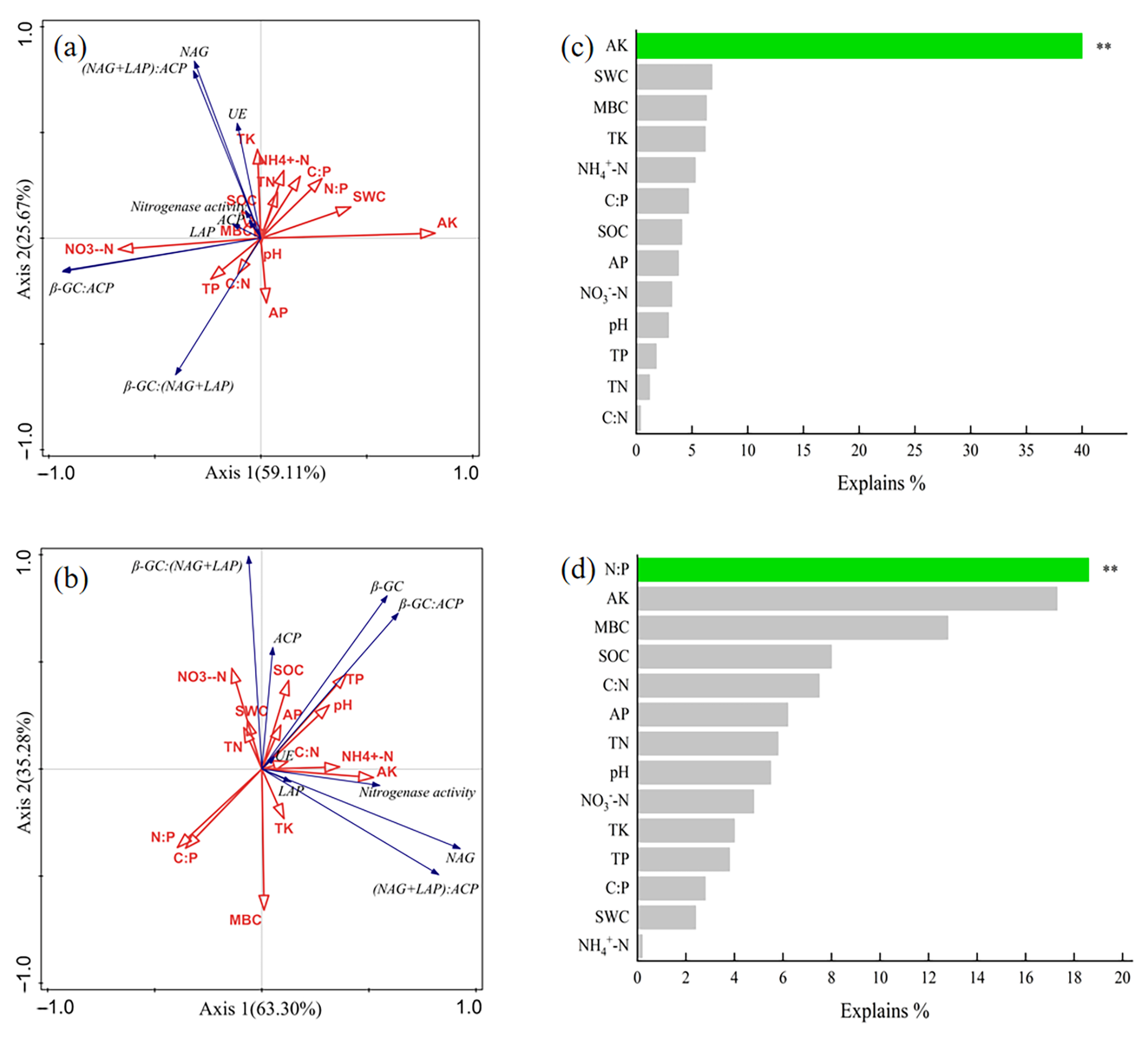
2.4. Data Processing and Analysis
3. Results
3.1. Chemical Properties of Rhizosphere and Bulk Soil
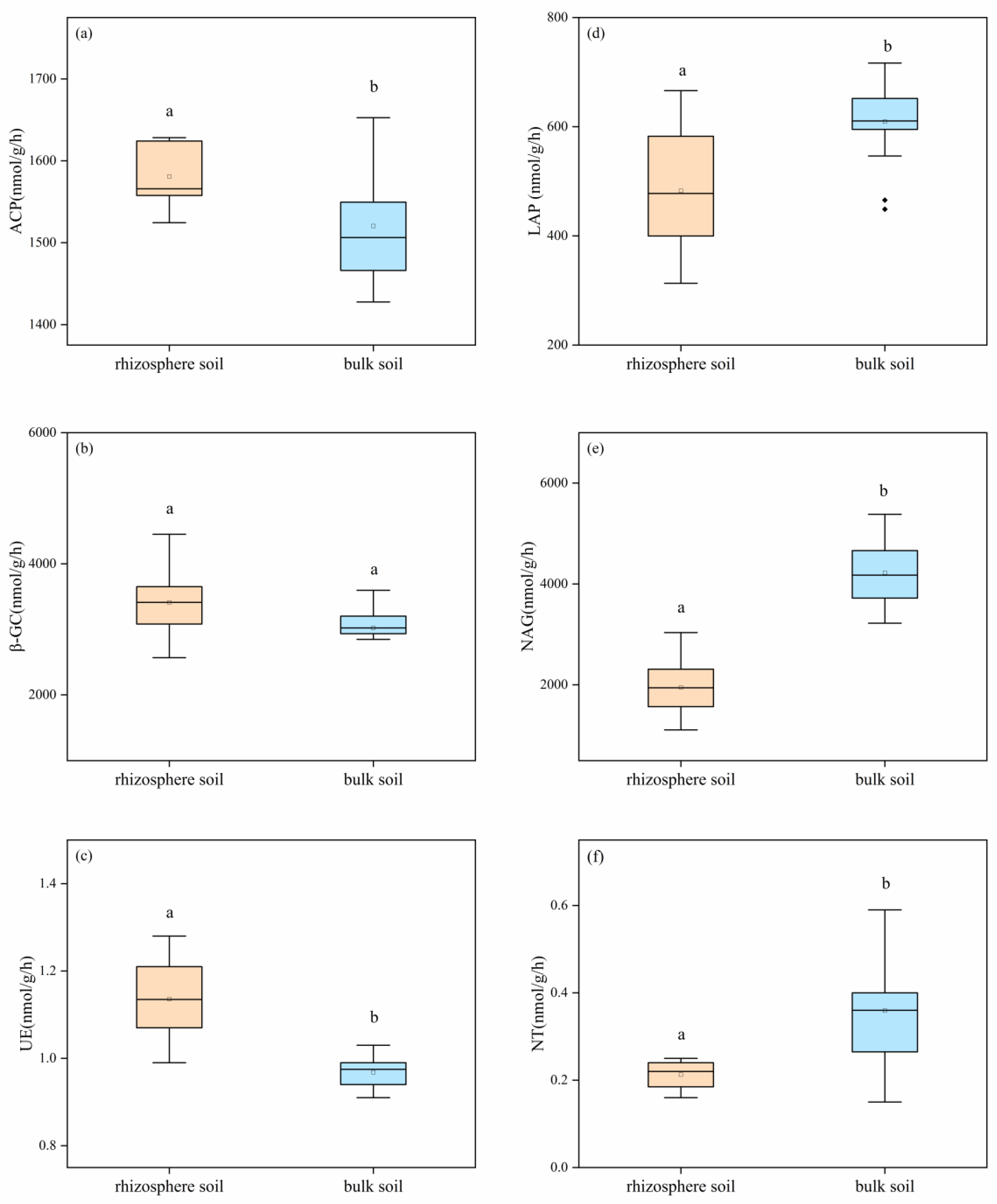
3.2. Enzyme Activities in Rhizosphere and Bulk Soils
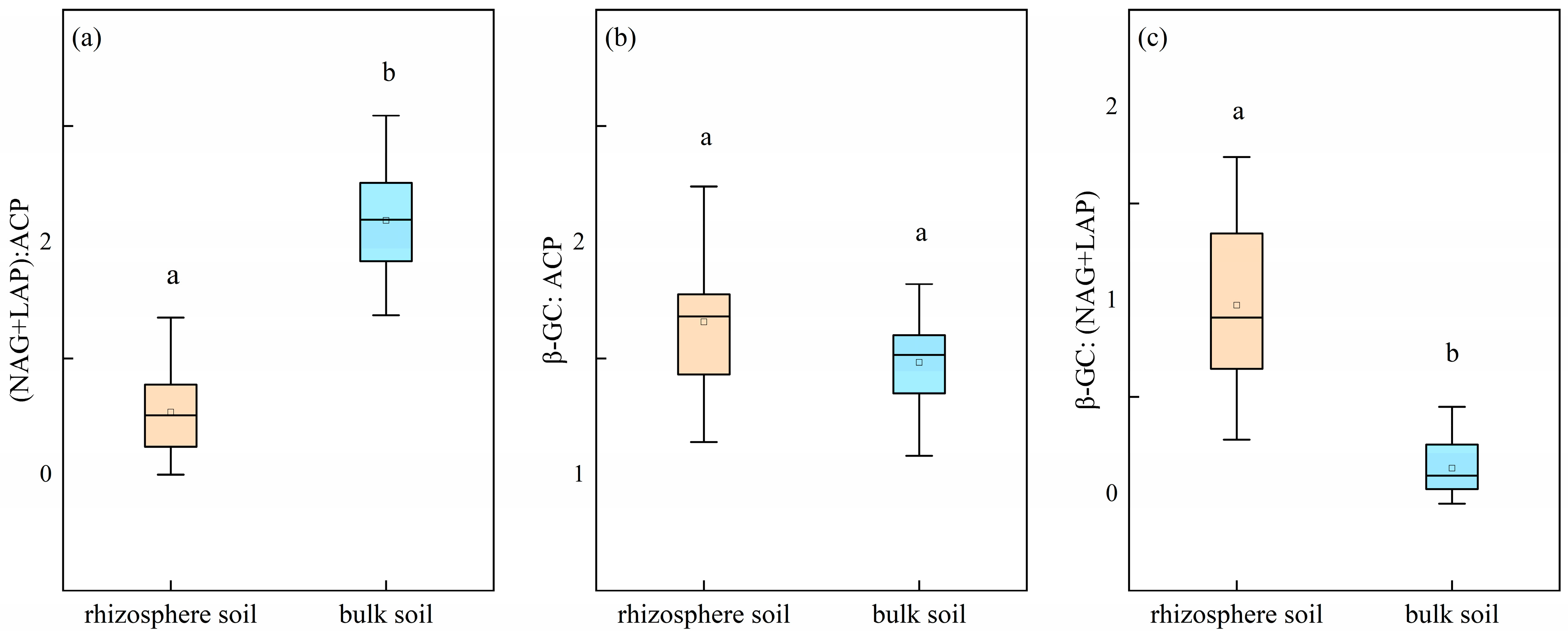
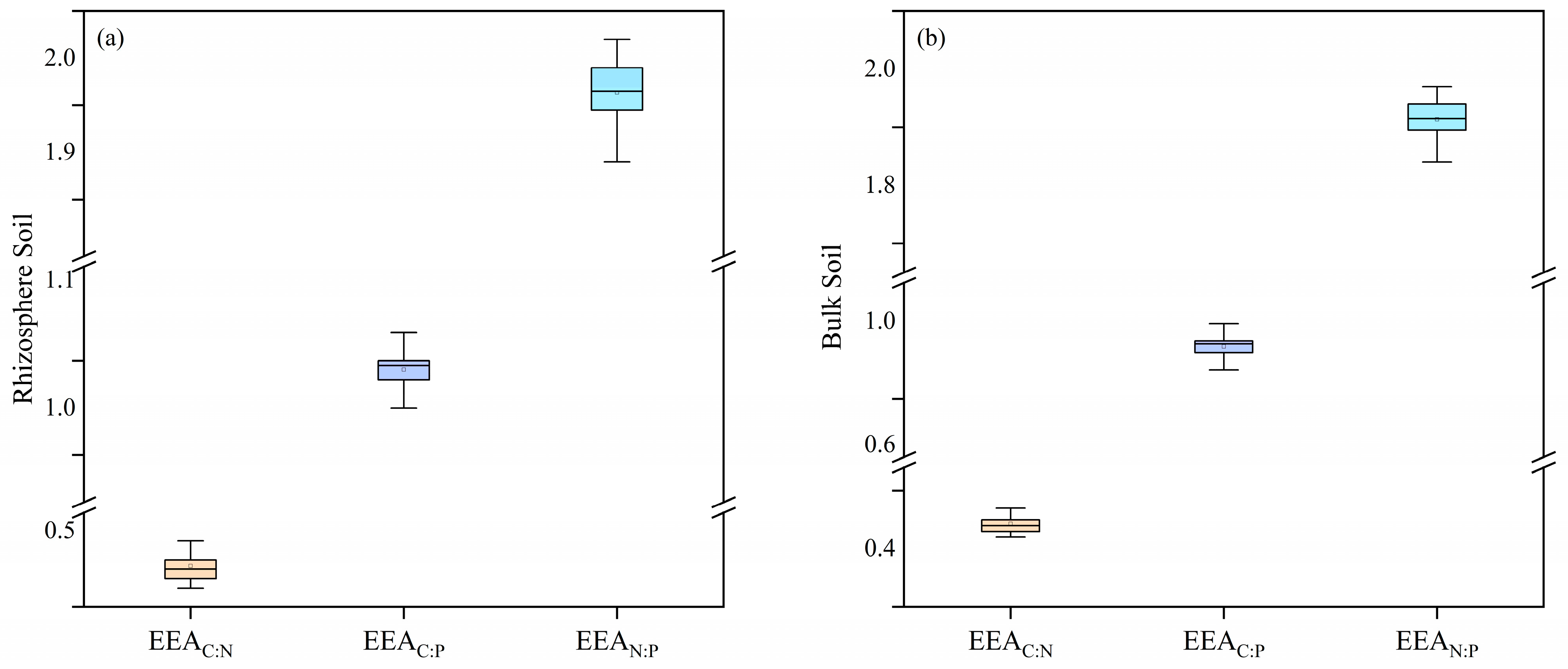
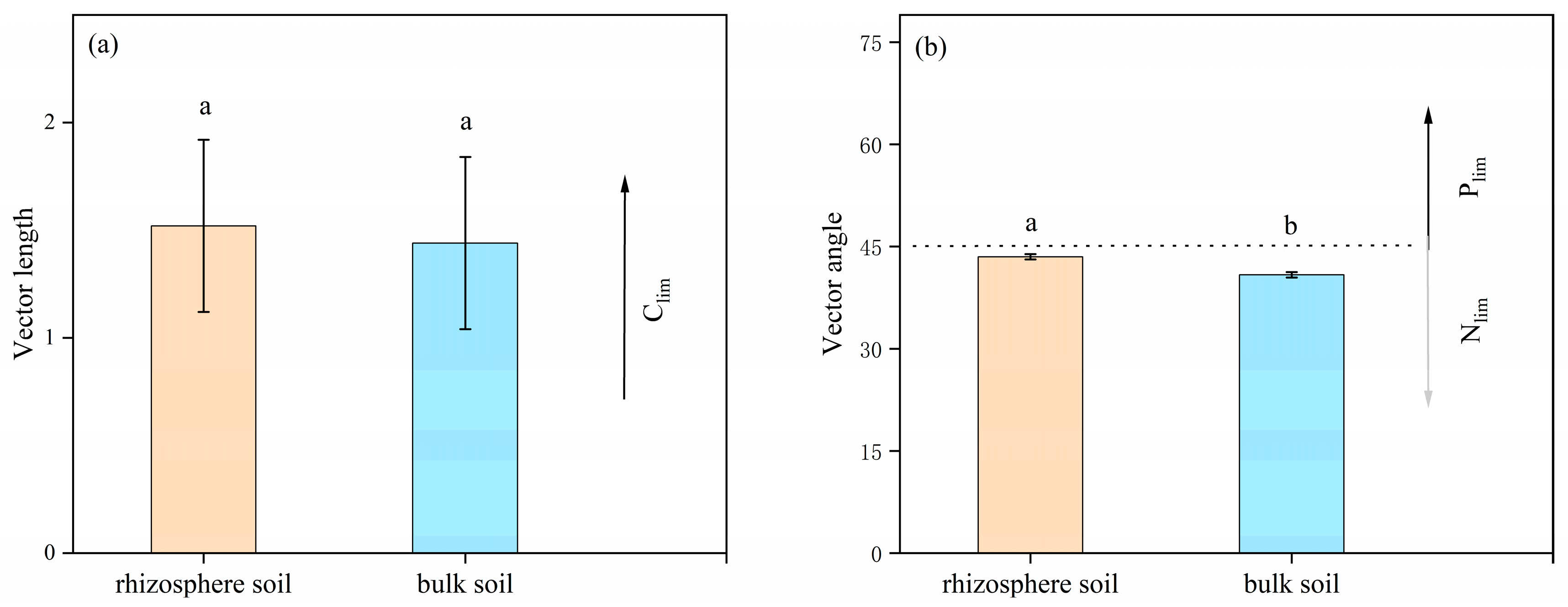
3.3. Stoichiometric Ratio and Vector Characteristics of Soil Enzymes
3.4. Factors Influencing Soil Enzyme Activity and Ecological Enzyme Stoichiometry
4. Discussion
4.1. Characteristics of Nutrients and Enzyme Activities in Rhizosphere and Bulk Soils
4.2. Ecoenzymatic Stoichiometry of Enzymes in Rhizosphere and Bulk Soils
4.3. Key Factors Affecting Enzyme Activity and Their Stoichiometric Ratio in Rhizosphere and Bulk Soils
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wu, X.D.; Zhao, L.; Fang, H.B. Soil Enzyme Activities in Permafrost Regions of the Western Qinghai-Tibetan Plateau. Soil Boil. Biochem. 2012, 76, 1280–1289. [Google Scholar] [CrossRef]
- Burns, R.G. Enzyme activity in soil: Location and A Possible Role in Microbial Ecology. Soil Boil. Biochem. 1982, 14, 423–427. [Google Scholar] [CrossRef]
- Huang, H.L.; Zong, N.; He, N.P.; Tian, J. Soil Enzyme Stoichiometric Characteristics at Different Altitudes in Alpine Meadow of Qinghai Tibet Plateau. J. Appl. Ecol. 2019, 30, 3689–3696. [Google Scholar]
- Liu, C.H.; Ma, J.Y.; Qu, T.T.; Xue, Z.J.; Li, X.Y.; Chen, Q.; Wang, N.; Zhou, Z.C.; An, S.S. Extracellular Enzyme Activity and Stoichiometry Reveal Nutrient Dynamics during Microbially-Mediated Plant Residue Transformation. Forests 2022, 14, 34. [Google Scholar] [CrossRef]
- Guan, S.Y. Soil Enzymes and Their Research Methods; Beijing Agricultural Press: Beijing, China, 1986; pp. 14–19. [Google Scholar]
- Ladd, J.N. Origin and Range of Enzymes in Soil. In Soil Enzymes; Burns, R.G., Ed.; Academic Press: London, UK, 1978; pp. 51–96. [Google Scholar]
- Tabatabai, M.A. Enzymes. In Methods of Soil Analysis; Weaver, R.W., Augle, S., Bottomly, P.J., Bezdicek, D., Smith, S., Tabatabai, A., Wollum, A., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Jones, D.L.; Hodge, A.; Kuzyakov, Y. Plant and Mycorrhizal Regulation of Rhizodeposition. New Phytol. 2004, 163, 459–480. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Simon, G.; Jorge, M.V. The Role of Root Exudates in Rhizosphere Interactions with Plants and other Organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.E.; Darby, B.A.; Giasson, M.A.; Kramer, M.A.; Phillips, R.P.; Finzi, A.C. Stoichiometry Constrains Microbial response to Root Exudation-insights from a Model and a field Experiment in a Temperate Forest. Biogeosciences 2013, 10, 821–838. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, W.B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Marcy, D.S.; Gallo, T.B.; Gartner, S.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [Green Version]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic Stoichiometry of Microbial Nutrient Acquisition in Tropical Soils. Biogeochemistry 2014, 117, 101–113. [Google Scholar] [CrossRef]
- Tapia-Torres, Y.; Elser, J.J.; Souza, V.; García-Oliva, F. Ecoenzymatic Stoichiometry at the Extremes: How Microbes Cope in an Ultra-oligotrophic Desert Soil. Soil Boil. Biochem. 2015, 87, 34–42. [Google Scholar] [CrossRef]
- He, Q.Q.; Wu, Y.H.; Bing, H.J.; Zhou, J.; Wang, J.P. Vegetation Type rather than Climate Modulates the variation in Soil Enzyme Activities and Stoichiometry in Subalpine Forests in the Eastern Tibetan Plateau Geoderma. Geoderma 2020, 374, 114424. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, G.B.; Xue, S.; Li, Q.; Xue, S. Ecoenzymatic Stoichiometry and Microbial Nutrient Limitation during Secondary Succession of Natural Grassland on the Loess Plateau, China. Soil Till Res. 2020, 200, 104605. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Zhao, J.; Zhang, W.; Xiao, K.; Wang, K. Nitrogen addition aggravates microbial carbon limitation: Ev-idence from ecoenzymatic stoichiometry. Geoderma 2018, 329, 61–64. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Boil. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Mori, T. Does ecoenzymatic stoichiometry really determine microbial nutrient limitations? Soil Boil. Biochem. 2020, 146, 107816. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Follstad, S.; Jennifer, J. Ecoenzymatic Stoichiometry of Microbial Organic Nutrient Acquisition in Soil and Sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Follstad, S.; Jennifer, J. Ecoenzymatic Stoichiometry and Ecological Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.Q.; Wang, W. Stoichiometry of Soil Extracellular Enzyme Activity along a Climatic Transect in Temperate Grasslands of Northern China. Soil Boil. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Cui, Y.X.; Bing, H.; Fang, L.; Mao, J.; Shen, G.T.; Yu, J.L.; Wang, X.; Zhu, H.; Wu, Y.H.; Zhang, X.C. Extracellular Enzyme Stoichiometry reveals the Carbon and Phosphorus Limitations of Microbial Metabolisms in the Rhizosphere and Bulk Soils in Alpine Ecosystems. Plant Soil 2019, 468, 7–20. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, Y.D.; Gao, D.X.; Wang, X.; Liu, W.C.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.A.; Ren, G.X. Ecoenzymatic Stoichiometry and Nutrient Dynamics along a Revegetation Chronosequence in the Soils of abandoned Land and Robinia Pseudoacacia Plantation on the Loess Plateau, China. Soil Boil. Biochem. 2019, 134, 1–14. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, C.; An, S.S.; Cheng, H.; An, S.S.; Chang, S.X. Soil Extracellular Enzyme Stoichiometry reflects the Shift from P-to N-limitation of Microorganisms with Grassland Restoration. Soil Boil. Biochem. 2020, 149, 107928. [Google Scholar] [CrossRef]
- Hartmann, A.; Rothballer, M.; Schmid, M. Lorenz Hiltner, A Pioneer in Rhizosphere Microbial Ecology and Soil Bacteriology Research. Plant Soil 2008, 312, 7–14. [Google Scholar] [CrossRef]
- Gartner, T.B.; Treseder, K.K.; Malcolm, G.M.; Sinsabaugh, R.L. Extracellular Enzyme Activity in the Mycorrhizospheres of a Boreal Fire Chronosequence. Pedobiologia 2012, 55, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G.; Römheld, V. Rhizosphere Chemistry in Relation to Plant Nutrition. In Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2012; pp. 347–368. [Google Scholar]
- Marschner, P. Rhizosphere biology. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Colac, Australia, 2012; pp. 369–388. [Google Scholar]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; Zhou, W. Responses of Extracellular Enzyme Activities and Microbial Community in Both the Rhizosphere and Bulk Soil to Long-term Fertilization Practices in a Fluvo-aquic Soil. Geoderma 2012, 173, 330–338. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, M.; Kou, Y.; Liu, D.Y.; Liu, Q.; Zhang, Z.L.; Jiang, Z.; Yin, H.J. Differential Effects of N addition on the Stoichiometry of Microbes and Extracellular Enzymes in the Rhizosphere and Bulk Soils of an Alpine Shrubland. Plant Soil 2020, 449, 285–301. [Google Scholar] [CrossRef]
- Cheng, W.; Kuzyakov, Y. Root Effects on Soil Organic Matter Decomposition. Roots Soil Manag. Interact. Roots Soil 2005, 48, 119–143. [Google Scholar]
- Finzi, A.C.; Abramoff, R.Z.; Spiller, S.K.; Brzostek, E.R.; Darby, B.A.; Kramer, M.A.; Phillips, R.P. Rhizosphere Processes are Quantitatively Important Components of Terrestrial Carbon and Nutrient Cycles. Gcb Bioenergy 2015, 21, 2082–2094. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Putten, V.D.; Wim, H. Going back to the Roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, T.; Shao, P.; Sun, J.; Xu, W.; Zhang, Z. Variation in bacterial community structure in rhizosphere and bulk soils of different halophytes in the yellow River Delta. Front. Ecol. Evol. 2022, 9, 816918. [Google Scholar] [CrossRef]
- Phillips, R.P.; Finzi, A.C.; Bernhardt, E.S. Enhanced Root Exudation induces Microbial Feedbacks to N Cycling in a Pine Forest Under long-term CO2 Fumigation. Ecol. Lett. 2011, 14, 187–194. [Google Scholar] [CrossRef]
- Avazpoor, Z.; Moradi, M.; Basiri, R.; Mirzaei, J.; Taghizadeh-Mehrjardi, R.; Kerry, R. Soil Enzyme Activity variations in Riparian Forests in relation to Plant Species and Soil Depth. Arab. J. Geosci. 2019, 12, 708. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Greco, A.; Drake, J.E.; Finzi, A.C. Root Carbon inputs to the Rhizosphere stimulate Extracellular Enzyme Activity and increase Nitrogen Availability in Temperate Forest Soils. Biogeochemistry 2013, 115, 65–76. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Z.J.; Tang, M.; Zhou, B. Greater Variations of Rhizosphere Effects within Mycorrhizal Group than between Mycorrhizal Group in a Temperate Forest. Soil Boil. Biochem. 2018, 126, 237–246. [Google Scholar] [CrossRef]
- Xu, Z.W.; Yu, G.R.; Zhang, X.Y.; He, N.P.; Wang, Q.F.; Wang, S.Z.; Wang, R.L.; Zhao, N.; Jia, Y.L.; Wang, C.Y. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China. Soil Boil. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Cui, Y.X.; Fang, L.C.; Guo, X.B.; Wang, X.; Zhang, Y.J.; Lia, P.F.; Zhang, X.C. Ecoenzymatic Stoichiometry and Microbial Nutrient Limitation in Rhizosphere Soil in the Arid Area of the Northern Loess Plateau, China. Soil Boil. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Sun, X.; Ye, Y.; Liao, J.; Soromotin, A.V.; Smirnov, P.V.; Kuzyakov, Y. Organic Mulching Increase Microbial Activity in Urban Forest Soil. Forests 2022, 13, 1352. [Google Scholar] [CrossRef]
- Phillips, R.P.; Fahey, T.J. The Influence of Soil Fertility on Rhizosphere Effects in Northern Hardwood Forest Soils. Soil Sci. Soc. Am. J. 2008, 72, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Peng, D.; Tian, H.; Wang, H.; Liu, X.; Cao, L.; Li, Z.; Cheng, S. Soil ecoenzymatic stoichiometry and microbial resource limitation driven by thinning practices and season types in Larix principis-rupprechtii plantations in North China. For. Ecol. Manag. 2021, 482, 118880. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agricultural Press: Beijing, China, 2000; pp. 26–114. [Google Scholar]
- Chen, H.; Li, D.J.; Xiao, K.C.; Wang, K.L. Soil microbial processes and resource limitation in karst and non-karst forests. Funct. Ecol. 2018, 32, 1400–1409. [Google Scholar] [CrossRef]
- Patra, A.K.; Abbadie, L.; Clays-Josserand, A.; Degrange, V.; Grayston, S.J.; Guillaumaud, N.; Loiseau, P.; Louault, F.; Mahmood, E.; Nazaret, S.; et al. Effects of Management Regime and Plant Species on the Enzyme Activity and Genetic Structure of N-fixing, Denitrifying and Nitrifying Bacterial Communities in Grassland Soguils. Environ. Microbiol. 2006, 8, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Keeler, B.L.; Hobbie, S.E.; Kellogg, L.E. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: Implications for litter and soil organic matter decomposition. Ecosystems 2009, 12, 1–15. [Google Scholar] [CrossRef]
- Fanin, N.; Moorhead, D.; Bertrand, I. Eco-enzymatic stoichiometry and enzymatic vectors reveal differential C, N, P dynamics in decaying litter along a land-use gradient. Biogeochemistry 2016, 129, 21–36. [Google Scholar] [CrossRef]
- Dai, X.Q.; Fu, X.L.; Kou, L.; Wang, H.M.; Shock, C.C. C:N:P Stoichiometry of Rhizosphere Soils Differed Significantly among Overstory Trees and Understory Shrubs in Plantations in Subtropical China. Can. J. For. Res. 2018, 48, 1398–1405. [Google Scholar] [CrossRef]
- Meier, I.C.; Pritchard, S.G.; Brzostek, E.R.; McCormack, M.L.; Phillips, R.P. The Rhizosphere and Hyphosphere Differ in their Impacts on Carbon and Nitrogen Cycling in Forests exposed to elevated CO2. New Phytol. 2015, 205, 1164–1174. [Google Scholar] [CrossRef]
- Gao, D.; Bai, E.; Wang, S.; Zong, S.; Liu, Z.; Fan, X.; Zhao, C.; Hagedorn, F. Three-dimensional mapping of carbon, nitrogen, and phosphorus in soil microbial biomass and their stoichiometry at the global scale. Glob. Change Biol. 2022, 28, 6728–6740. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P Stoichiometry in Soil: Is there a “Redfield ratio” for the Microbial Biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Wojciech, P.; Ewa, B.; Jaroslaw, L.; Lukac, M. A Comparison of C:N:P Stoichiometry in Soil and Deadwood at An advanced Decomposition Stage. Catena 2019, 179, 1–5. [Google Scholar]
- Wardle, D.A.; Walker, L.R.; Bardgett, R.D. Ecosystem Properties and Forest decline in Contrasting long-term Chronosequences. Science 2004, 305, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.X.; Parton, W.J.; Gonzalez-Meler, M.A.; Phillips, R.P.; Asao, S.; McNickle, G.G.; Brzostek, E.; Jastrow, J.D. Synthesis and modeling perspectives of rhizosphere priming. New Phys. 2014, 201, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools. Soil Boil. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Moorhead, D.L. Resource Allocation to Extracellular Enzyme Production: A Model for Nitrogen and Phosphorus Control of Litter Decomposition. Soil Boil. Biochem. 1994, 26, 1305–1311. [Google Scholar] [CrossRef]
- Treseder, K.K.; Vitousek, P.M. Effects of Soil Nutrient Availability on Investment in Acquisition of N and P in Hawaiian rain Forests. Ecology 2001, 82, 946–954. [Google Scholar] [CrossRef]
- Zhao, Q.; Classen, A.T.; Wang, W.W.; Zhao, X.R.; Mao, B.; Zeng, D.H. Asymmetric effects of litter removal and litter addition on the structure and function of soil microbial communities in a managed pine forest. Plant Soil 2017, 414, 81–93. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.; Hu, X.F.; Wan, S.; Wang, H.; Liang, C.; Chen, F.S. Litter manipulation effects on microbial communities and enzymatic activities vary with soil depth in a subtropical Chinese fir plantation. For. Ecol. Manag. 2021, 480, 118641. [Google Scholar] [CrossRef]
- Wang, L.; Deng, D.; Feng, Q.; Xu, Z.; Pan, H.; Li, H. Changes in litter input exert divergent effects on the soil microbial community and function in stands of different densities. Sci. Total Environ. 2022, 845, 157297. [Google Scholar] [CrossRef]
- Ai, L.; Wu, F.; Fan, X.; Yang, Y.; Zhang, Y.; Zheng, X.; Zhu, J.; Ni, X. Different effects of litter and root inputs on soil enzyme activities in terrestrial ecosystems. Appl. Soil Ecol. 2023, 183, 104764. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Ding, C.; Liu, Y.; Wu, K.; Jiang, F.; Su, D. Water addition promotes vegetation recovery of degraded alpine meadows by regulating soil enzyme activity and nutrients in the Qinghai–Tibetan Plateau. Ecol. Eng. 2020, 158, 106047. [Google Scholar] [CrossRef]
- Shi, L.J.; Wang, H.M.; Fu, X.L.; Kou, L.; Meng, S.W.; Dai, X.Q. Soil Enzyme Activities and their Stoichiometric Characteristics of Typical Plantations in the Middle Subtropical Zone. J. Appl. Ecol. 2020, 31, 1980–1988. [Google Scholar]
- Montagnoli, A.; Di Iorio, A.; Terzaghi, M.; Trupiano, D.; Scippa, G.S.; Chiatante, D. Influence of soil temperature and water content on fine-root seasonal growth of European beech natural forest in Southern Alps, Italy. Eur. J. For. Res. 2014, 133, 957–968. [Google Scholar] [CrossRef] [Green Version]
- Coners, H.; Leuschner, C. In situ measurement of fine root water absorption in three temperate tree species-temporal variability and control by soil and atmospheric factors. Basic Appl. Ecol. 2005, 6, 395–405. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Factors affecting rhizosphere priming effects. J. Plant Nutr. Soil Sci. 2002, 165, 382–396. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Name | EC Number | Abbreviation | Function |
|---|---|---|---|
| β-glucosidase | 3.2.1.2.1 | β-GC | Hydrolyzes glucose from cellobiose |
| Leucine aminopeptidase | 3.4.11.1 | LAP | Hydrolyzes leucine and other hydrophobic amino acids from the N terminus of polypeptides |
| N-acetyl-β-D-glucosidase | 3.4.11.1 | NAG | Degrades chitin and other β-1,4 glucosamine polymers |
| Acid phosphatase | 3.1.3.2 | ACP | Hydrolyzes phosphosaccarides and phospholipids to release phosphate |
| Urease | 3.5.1.5 | UE | Hydrolyzes NH3-N in urea |
| Nitrogenase | — | NT | Reduces nitrogen molecules to ammonia |
| Soil Type | TN (g/kg) | TP (g/kg) | TK (g/kg) | AP (mg/kg) | AK (mg/kg) | SOC (g/kg) | NH4+-N (mg/kg) | NO3−-N (mg/kg) | MBC (mg/L) | SWC (%) | pH | C:N | C:P | N:P |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rhizosphere soil | 10.49 ± 1.04 a | 1.02 ± 0.15 a | 7.02 ± 0.33 a | 26.82 ± 3.82 a | 496.98 ± 68.63 a | 172.99 ± 21.07 a | 13.96 ± 6.61 a | 87.48 ± 21.80 a | 1591.11 ± 249.10 a | 58.21 ± 13.49 a | 5.79 ± 0.27 a | 16.38 ± 1.53 a | 172.22 ± 14.51 a | 10.65 ± 1.14 a |
| Bulksoil | 5.38 ± 0.52 b | 0.89 ± 0.20 a | 8.66 ± 0.28 b | 16.01 ± 2.49 b | 308.12 ± 23.36 b | 69.61 ± 5.17 b | 9.15 ± 3.05 b | 27.23 ± 7.24 b | 697.04 ± 28.47 b | 39.4 ± 4.91 b | 6.01 ± 0.12 b | 13.06 ± 1.23 b | 81.03 ± 10.02 b | 6.26 ± 0.87 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Jia, Y.; Li, Q.; Cui, H.; Lu, J.; Ma, J.; Xu, Z. Ecoenzymatic Stoichiometry in the Rhizosphere and Bulk Soil of a Larix principis-rupprechtii Plantation in North China. Forests 2023, 14, 1315. https://doi.org/10.3390/f14071315
Yang L, Jia Y, Li Q, Cui H, Lu J, Ma J, Xu Z. Ecoenzymatic Stoichiometry in the Rhizosphere and Bulk Soil of a Larix principis-rupprechtii Plantation in North China. Forests. 2023; 14(7):1315. https://doi.org/10.3390/f14071315
Chicago/Turabian StyleYang, Liu, Yanlong Jia, Qianru Li, Hongna Cui, Jinping Lu, Jiaojiao Ma, and Zhongqi Xu. 2023. "Ecoenzymatic Stoichiometry in the Rhizosphere and Bulk Soil of a Larix principis-rupprechtii Plantation in North China" Forests 14, no. 7: 1315. https://doi.org/10.3390/f14071315