1. Introduction
Eucalyptus sp. plantations cover almost 1 million hectares in Uruguay, being the main source of wood production in the country [
1]. Of these, around 40,000 ha are harvested annually, expecting a significative increase in the short term. The sustainable management of these plantations is based on practices that seek to conserve the stocks of soil organic matter (SOM) and nutrients, both key components of soil fertility, and are mainly supported via the contributions of mulch (litter) and root replacement during the plantation cycle and through the harvest residue decomposition in the replanting of the site. Therefore, keeping these residues in the field has a great impact on soil nutrients [
2] and SOM stocks [
3], further helping to sustain the initial growth of trees [
4].
Maintaining soil with residues improves the different chemical, physical and bio-logical properties of the soil, resulting in an improvement in soil quality, while constituting a proportionately important nutrient reserve [
5].
Harvest residues, although they represent a smaller proportion of the total aerial biomass produced (about 30%), contain most of the nutrients absorbed [
6], which, through their decomposition, can be used by the next plantation. Understanding the dynamics and the factors that affect the decomposition and their effects on soil fertility is relevant to the management of forest plantations [
7].
Spangenberg et al. [
8] established that both the quantification of the nutrient content in the different forest plantations and the knowledge of the relationship between the export of nutrients and those that are available in the soil for subsequent reuse are essential for defining management strategies with the aim of maintaining the sustainability of the ecosystem.
The decomposition of the different components of the aerial biomass that remain at the site (bark, branches and leaves) on the soil occurs at variable rates and depends on the intrinsic characteristics of the residue itself in terms of its physical and chemical structure [
9], specific climatic conditions such as humidity and temperature [
10,
11], and the period of time during which the processes occur. Regarding the effect of these factors on the process, [
12] indicates that there is still little knowledge about the decomposition of lignified woody residues, particularly in ecosystems of
Eucalyptus sp., recognizing that the climate has an important control on a regional scale but not so when evaluated in narrower ranges. Ferreira et al. [
13] reported that the decomposition of residues and the nutrient release were more controlled by the chemical properties of the residues than by climatic or soil characteristics. Similarly, De Souza [
14] identified that rainfall increases did not accelerate the decomposition times of surface residues, even though it did for residues that were buried.
N contents, as well as the carbon (C):N ratio, are very important in the decomposition rate of plant materials [
15], since microorganisms, although they base their activity on the availability of carbon, need certain amounts of the other nutrients, and given the scarcity of the latter, decomposition is slowed. In residues of large physical size and little contact with the soil—such as branches— their slow decomposition acquires even more relevance, because, when residues are incorporated, the mineral N of the soil is immobilized by the microbial biomass and, in this way, the soil provides the N necessary for growth [
16]. The lignin (L) and phenolic compounds content in plant residues also affect the N mineralization [
17]. Higher lignin content makes the materials more resistant to decomposition, so residues with high L:N ratios tend to decompose slowly [
10,
18]. In turn, phenolic compounds act as microbial inhibitors affecting the decomposition processes and nutrient cycles of the soil through multiple mechanisms [
19].
Since E. grandis and E. globulus are widely planted species of the genus Eucalyptus (E. grandis is found in Brazil, Argentina, South Africa, Sri Lanka, India or New Zealand and E. globulus in Spain, Portugal and Chile), experimental information for both regarding the decomposition of residues is widely available, although it is mostly derived from climates different from that of Uruguay. However, experimental information on the recycling of nutrients from the harvest residues is limited, despite its importance for developing a fertilization program for the future replanting of the site. On the other hand, for the species E. dunnii, there is little information available worldwide regarding the rates of residue decomposition or the process of nutrient recycling, since this species has only rarely been planted across the globe—although today it shows a clear expansion in temperate regions—despite being the species most commonly planted in Uruguay. Thus, there is a need for comparative experimental information about the decomposition rates of residues and recycling of nutrients to the soil by these species, which will allow researchers to obtain specific parameters that can be used in models of decomposition and cycling of nutrients from regions climatically such as the one corresponding to this research.
The hypothesis of this work was that the decomposition rate of the different harvest components of Eucalyptus sp. is variable and highly dependent on particle size and chemical constitution, as well as on the species itself.
The main objectives of this work were:
- (a)
To identify and quantify the characteristics of each species (E. dunnii, E. grandis and E. globulus) that affect the decomposition rates of the different harvest residues, as well as the indicators that can explain the process.
- (b)
To quantify the potential recycling of N, P, K, Ca and Mg to the soil from the decomposition of these residues and assess the quantitative and qualitative differences between the species evaluated.
2. Materials and Methods
The studies of the residue decomposition “in situ” were carried out after harvest of commercial plantations of the genus
Eucalyptus (
E. dunnii,
E. grandis and
E. globulus), destined for the production of cellulose, between 9 and 10 years old and located in the northeast and west of the country. In 2 of the 5 cases, previously published information was used [
20,
21]. At all the sites, the methodology regarding the quantification of harvest residue, soils and plant sampling was similar. The harvests were carried out between 2007 and 2011, between the months of June and September.
- -
Location of experimental sites
The coordinates of the different experimental sites as well as some of the properties of the soils (A horizon) are presented in
Table 1.
- -
Climatic characteristics of the study areas
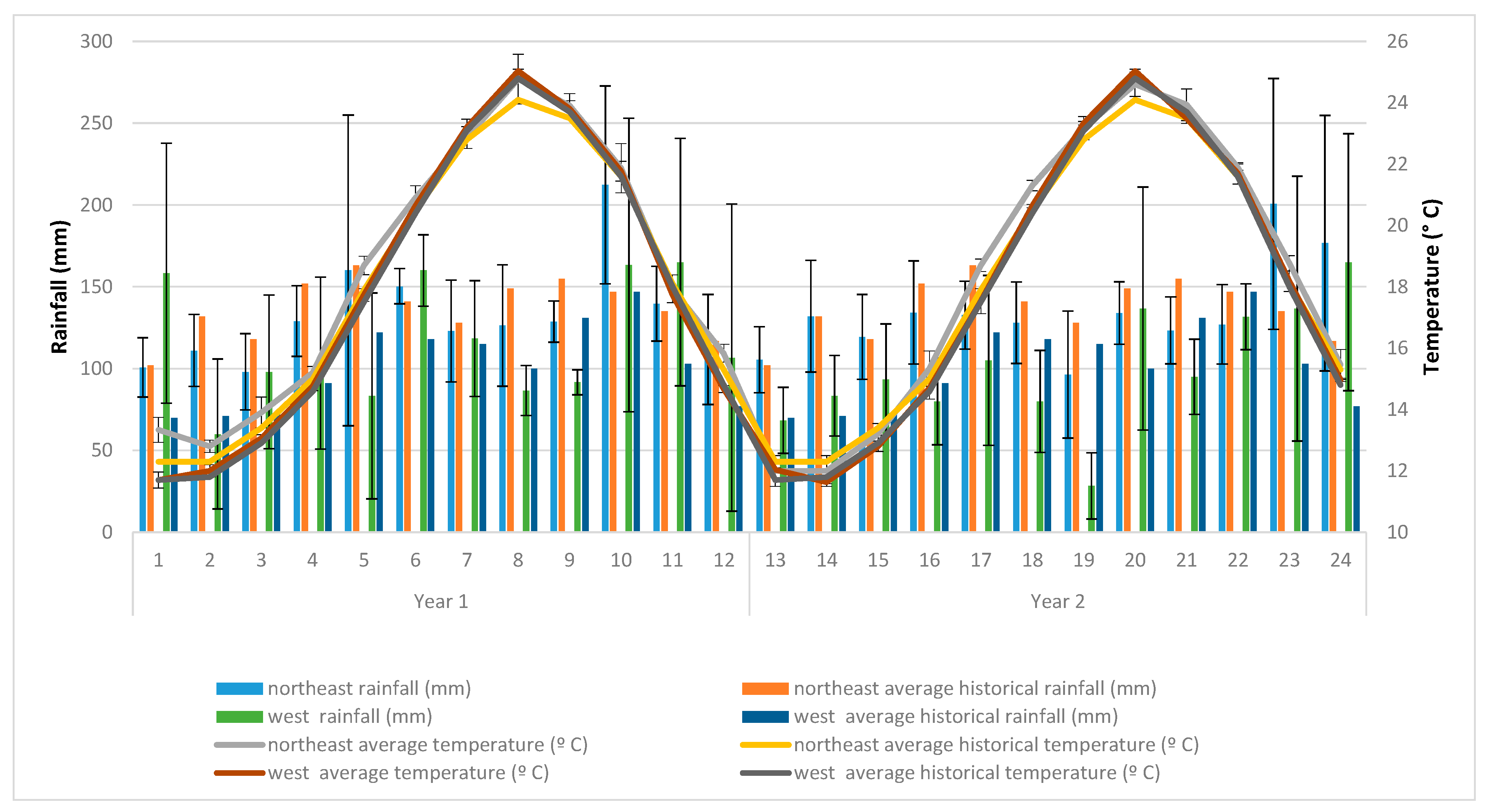
According to the Köppen–Geiger climate classification, the territory of Uruguay corresponds to the temperate climate zone (Cf), with an average temperature of 17.5 °C (16 °C and 19.5 °C range, in the south and northwest, respectively), with hot summers, similar monthly average rainfall throughout the year and four clearly differentiated seasons. The highest temperatures occur in January and February, and the lowest in June and July, with greater thermal amplitude in the north than in the south. The relative humidity is high, ranging from 70% to 75% throughout the country.
The precipitation and average monthly mean temperature data recorded at the sites and the respective historical averages [
22] are presented in
Figure 1.
- -
Plant sampling and chemical analysis
Samples of known leaf weight (100 g) and thin branches (diameter < 1 cm, 150 g) were placed in mesh bags (1 mm2), while the pieces of thick branches (diameter > 1 cm and 15 cm long) and bark (15 cm long and 3–4 cm wide) were weighed and individually identified. As many samples of each component of the residues were prepared, sampling dates were planned, and they were placed on the ground in three zones (replications) according to topographical position. Each group of samples was protected with a wire mesh fabric (100 mm2). At pre-established sampling dates (1, 2, 4, 6, 9, 12, 18 and 24 months post-harvest), samples were taken from each residue (one by replication). At the time of harvest, samples were also taken from each component.
The samples of the different residues were dried at 65 °C to constant weight and subsequently milled to particle sizes of less than 0.5 mm for subsequent chemical analysis.
The concentrations of P, Ca, Mg and K of the samples were determined after dry combustion at 550 °C and ash dissolution through 10% HCl. In the extracts, P was deter-mined via colorimetry [
23], Ca and Mg via atomic absorption spectrophotometry and K via emission spectrophotometry. The determination of the N concentration in the sample was performed through the wet digestion of the sample (H
2SO
4 and catalyst mixture) and the subsequent distillation of N, according to the method by Kjeldahl [
24]. The total carbon and soluble carbon contents of the residues were measured via oxidation with K
2Cr
2O
7 at 150 °C followed by colorimetric determination [
24]. The determination of lignin was performed using acid hydrolysis with H
2SO
4 then diluted to an acid concentration of 3%, with subsequent boiling for 4 h and the filtering and determination of the remaining materials via gravimetry [
25]. For the analysis of soluble polyphenols (Pol), the extraction was performed using H
2O and determination was carried out via colorimetry using the Folin–Ciocalteu method [
26].
- -
Calculations and statistical analysis of the information
The biomass data of harvest residues were annualized (dividing them by the age of the plantation) to facilitate their analysis and comparison.
The biomass decomposition was calculated as: biomass at time t (P)/initial biomass (Pinic) × 100. From this, the rate of decomposition was calculated by adjusting an exponential decay model type P/Pinic = e
−kt, where P/Pinic is the proportion of the remaining material at a time t measured in years and k is the decomposition constant [
27]. This model also allowed the calculation of the half-life in the years of the different crop residues (t
1/2).
For the three species studied: (1) analysis of variance was performed for (a) the amount of biomass residue according to species and (b) the half-life according to species and type of residue; (2) multivariate analysis was performed through the analysis of main components (PCA) of decomposition after 24 months, considering the different parts and species; and (3) the regression models for the decomposition of the different components according to different ratios (C:N, Pol:N and L:N) and the soluble carbon concentration.
As a first step, the normality of the variables under study (half-life) was verified via the Shapiro–Wilk test (p > 0.05 for normal data). The differences were considered statistically significant when p < 0.05 (Tukey’s test).
The correlations between the half-life of the different components were analyzed with respect to the ratios C:N, Pol:N and L:N, as well as in relation to the concentration of soluble carbon. For the correlation analysis, Spearman’s coefficient [
28] was used since some of the variables did not have a normal distribution.
The nutrient contents of the residues were calculated considering the biomass of each component and the concentration of the nutrients at each date. In turn, the total nutrients released during the first 12 months and the entire period was calculated as the difference between the nutrient present in the residues at the end of the decomposition period (12 and 24 months) and the amount determined immediately after harvest.
4. Discussion
The little experimental information that exists worldwide on
E. dunnii has shown that this species is characterized by a higher proportion of harvest residues in relation to the total aerial biomass [
29,
30], compared to other widely planted and studied species, such as
E. grandis and
E. globulus [
7,
31].
In this study, the proportion of harvest residues reached 35% in
E. dunnii and 24% in the other two species (
E. grandis and
E. globulus). Furthermore, the growth of this species and
E. grandis was higher than that of
E. globulus [
32]. Shammas et al. [
31], in a 7-year-old
E. globulus plantation in the SW of Australia, in relation to the aerial biomass of residues, found a similar proportion for thick branches, a lower proportion for bark, and a higher proportion for thin branches and leaves compared to the same species in this research. In the present investigation, although the bark was the part with the highest proportion in all species (
Table 2), its proportion was higher in
E. globulus compared to
E. dunnii and
E. grandis. Likewise, differences were also registered in the minority parts, i.e., the leaves in
E. dunnii and the thin branches in
E. grandis and
E. globulus, which can be explained by the differences in the structure of the crown, with more branches in
E. dunnii.
Although the decomposition studies analyzed in this work were not carried out in the same period, differences in the climatic conditions between them were not of great magnitude (
Figure 1). On the other hand, it has been observed that in long evaluation periods (24 months), the significance of extreme climate events is mitigated [
12,
33]. Likewise, in studies of forest litter decomposition in northern Uruguay, Baietto et al. [
11] tested the effect of the starting season of the decomposition period on the pattern of biomass loss, finding that this effect was not significant. Similarly, Ferreira et al. [
13] reported that the release of nutrients was more controlled via the management and the physical and chemical properties of the residues than by the climate or the soil properties.
The highest decomposition constants (k) were reported in the leaves part. Rezende et al. [
9] for
E. grandis in incubation experiments under controlled conditions in Brazil, reported an annual decomposition constant for leaves (k) of 0.59, slightly higher than that found in the present work for the same species.
Rocha et al. [
7], for 12-year-old
E. grandis in Sao Paulo, Brazil, found higher values of k in leaves than in bark and in branches, which resulted in the same order as in the present research for this species, although with higher values in all cases (3.6, 1.2 and 0.5 for leaves, bark and branches, respectively) when compared with the same sample types in our research. Beyond the climatic differences between the sites, the higher k values reported by these authors in a humid sub-tropical climate [
34] could also be explained by the addition of various nutrients as fertilizers at the time of reforestation (130, 44, 125, 480 and 120 kg ha
−1 of N, P, K, Ca and Mg, respectively), which stimulates decomposition through microbial biomass.
Shammas et al. [
31], for 7-year-old
E. globulus trees in Australia, found the highest values of k in leaves, then in bark and, finally, in branches, that is, a similar order to that of the same species in the present work. The values reported by these authors were slightly higher than those of this study for bark and thin and thick branches (0.22, 0.21 and 0.16, respectively) and higher for leaves (1.54). Considering similarities in the climate (rainfall and temperature), the slight variations could be explained by differences in age (7 and 10 years, for Australia and Uruguay, respectively), with younger tissues in the case of Australia. From the decomposition constant (k) the half-life of each of the remains can be estimated. In this sense, the half-life showed significant differences in the interaction between species and part, with the
E. dunnii bark being the component with the highest durability (t
½ = 5.62 years). This species’ bark thickness, which was significantly higher than the others, could help explain the above. In the other species (
E. grandis and
E. globulus), the branches (thick and thin) were the parts with the highest half-life, similar to the result reported by different authors for the same species [
7,
31,
33,
35]. In general, and except for the leaves, the harvest residues decomposed slowly, which is positive considering the protection they exert against possible erosive events on the soil. In Entre Ríos, Argentina, with similar climatic conditions, the authors of [
36], in a field decomposition experiment with
E. globulus, found a half-life for leaves similar to that of this study for this species (1.5 years).
Jones et al. [
33], for plantations of 11-year-old
E. globulus in Monte Jarrio, Spain, with similar rainfall (1,119 mm year
−1) and lower average annual temperatures (13.1 °C), reported a similar half-life for thin and thick branches (4.0 and 5.0 years, respectively) compared to the same species in our study. The same authors, for the same species and age, in Furadouro and Vale Pequeno, Portugal, with a mean annual temperature of 15.6 °C and much lower rainfall (630 mm yr
−1), indicated a longer half-life for thin and thick branches (5.6 and 6.1 years, respectively) compared to the same species in our research. The low rainfall in these areas of Portugal (about 50% less) helps explain the lower decomposition found. In these studies, the soils differed from each other (ranges 6.5–53 g kg
−1 and 85–290 g kg
−1 for organic carbon and clay, respectively, in the first 20 cm of soil), although no differences were reported for this cause. Coincidentally, the authors of [
37] indicated that the parent material and the soils did not significantly influence the decomposition of the litter or the nutrient dynamics for the different forest species studied (
Eucalyptus,
Pinus and
Quercus). Similar results were obtained in the present research, being the relative homogeneity of the soils, all of them acidic and poor in nutrients, a possible explanation for this behavior (low variability in the population and activity of microorganisms).
In their study with
E. globulus, the authors of [
31] found slightly lower half-lives for bark, and thin and thick branches parts (3.1, 3.4 and 4.3 years, respectively), and lower half-lives for leaves (0.4 years). The younger age of the trees (7 year old) compared to those of the present work could explain these small differences. Likewise in Australia, similar values for the components thin and thick branches of
E. globulus were indicated by [
35]. Rocha et al. [
7] for
E. grandis in Sao Paulo, Brazil, reported lower half-life values compared to the same species in this study (t
½ of 0.2, 0.6 and 1.5 for leaves, bark and branches, respectively), although adding all the nutrients at the time of new planting, which could accelerate the decomposition process, in addition to the temperature differences mentioned above.
Several causes could explain differences in the rates of decomposition of the various residues. The specific surface of each part was an important factor, being smaller in those of greater size (bark, branches), which, in turn, were those of slower decomposition and, therefore, had a longer half-life.
The contents of more unstable components or a more labile chemical structure, such as soluble organic compounds (e.g., soluble carbon), also influence decomposition of the residues with longer half-lives and the most resistant constituents, as shown by [
31]. In the present investigation, the highest soluble carbon contents were found in the leaf part, which regardless of the species, presented the highest rates of decomposition (
Table 3 and
Table 4). The thick branches, meanwhile, showed the lowest concentrations. Data reported by [
9,
15] in
E. grandis residue mineralization studies in Brazil and Australia, respectively, also indicated a high concentration of leaf-soluble carbon.
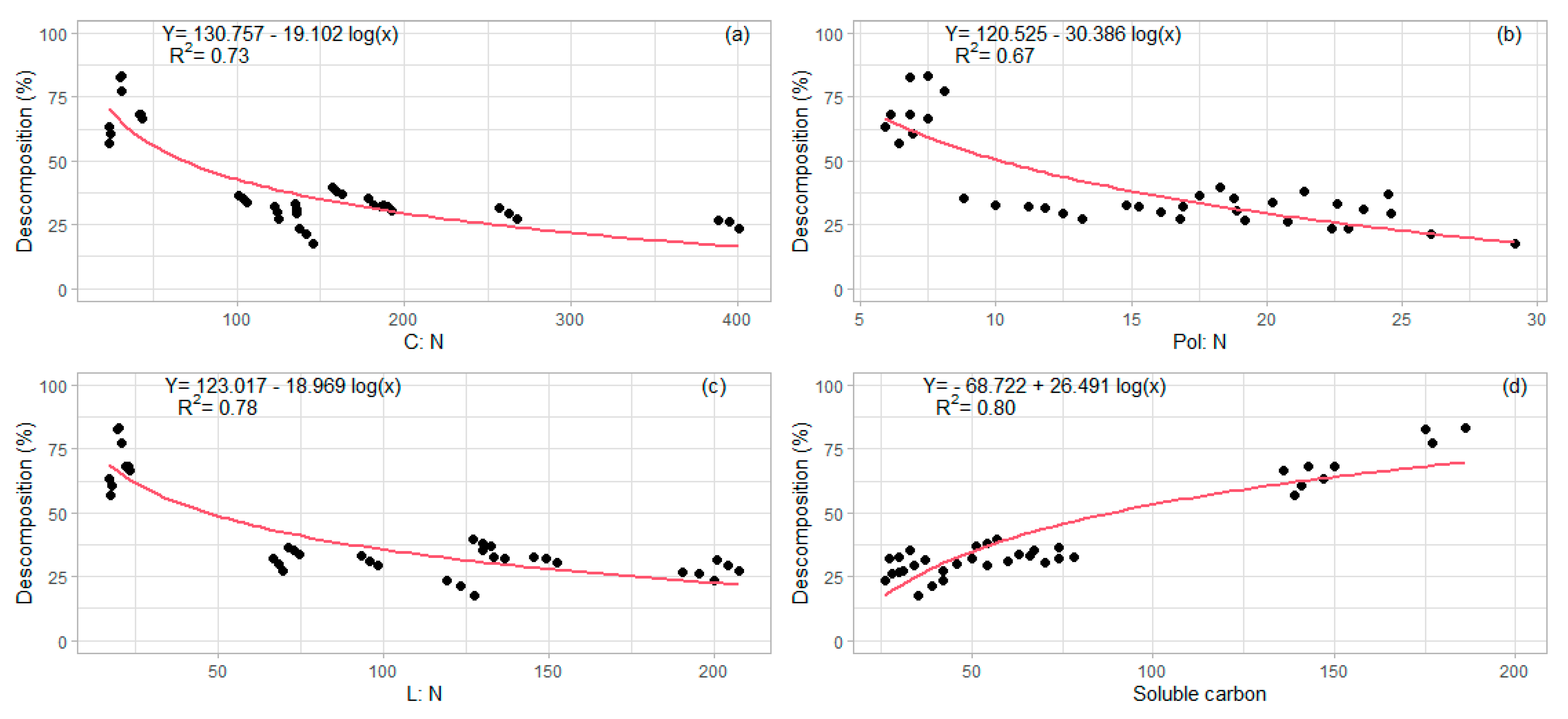
The N contents as well as the C:N ratio also differed between the residues. According to [
38], the incorporation to the soil of residues with a C:N < 25 ratio can cause net N mineralization, while C:N > 25 ratio tends to cause net immobilization of N. The present study showed that the highest concentrations of N were found in the leaves, as well as lower C:N ratios (
Table 4). The other parts presented very low N concentrations and high C:N ratios. In addition, the ratios of lignin and polyphenols with N were even more decisive than the concentrations of the former, as reported by [
10]. Higher Pol:N or L:N ratios resulted in lower rates of decomposition (
Figure 4).
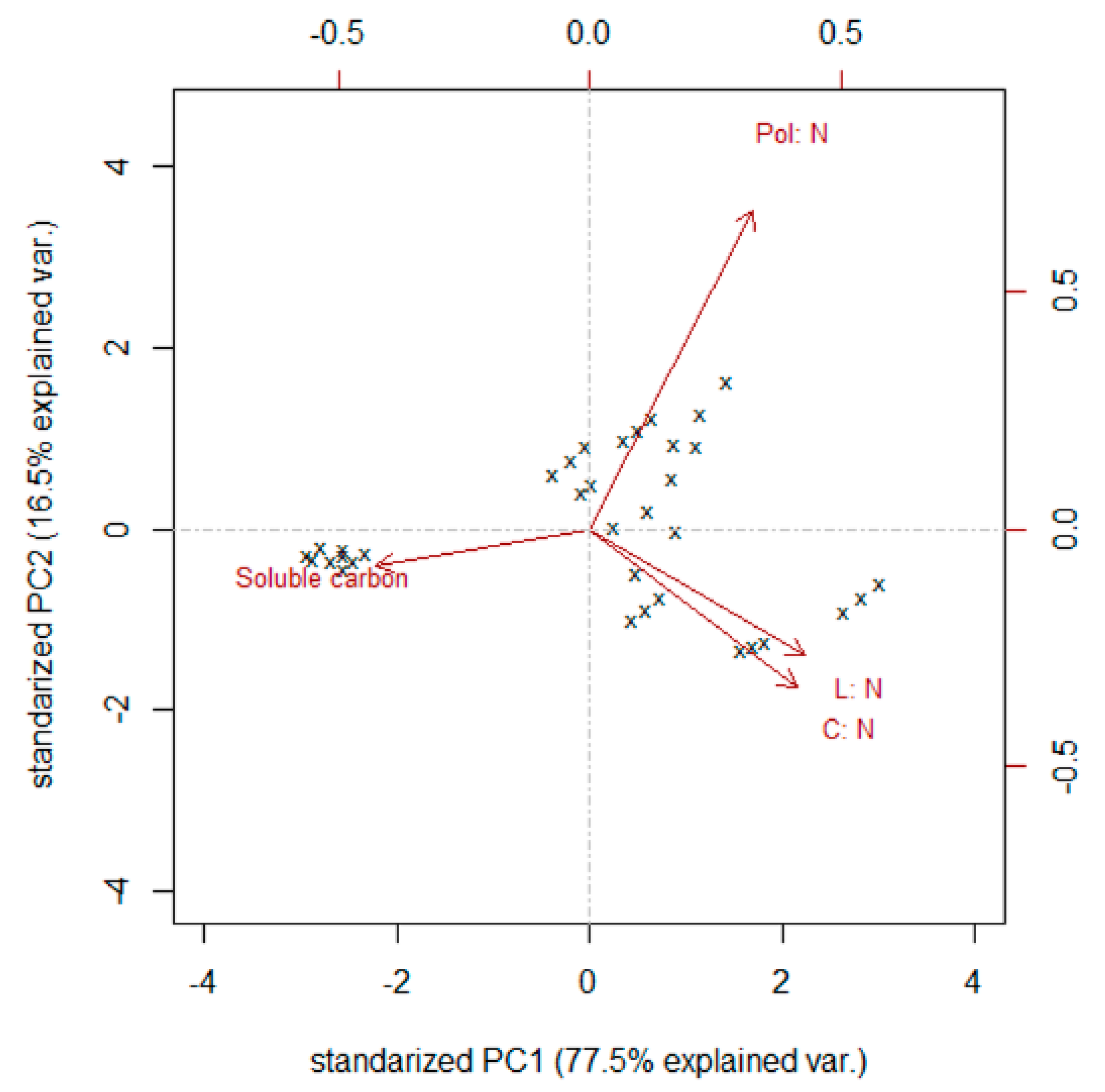
The results of the PCA showed that with the first two components, it was possible to explain almost the entire variation. The C:N, Pol:N and L:N ratios showed an acute angle that suggests a positive correlation, while soluble carbon presented a negative correlation with these variables (opposite vectors and flat angles). It can be inferred that the species under study had distinctive values of these variables since, in the biplot, the observations for the same species appeared grouped.
The high values of the correlations between the half-life of the residues and the chemical parameters indicate a high magnitude of the association. None of the chemical characteristics studied could individually explain the differences between the rates of decomposition, but all of them had a similar influence (
Table 5).
The greater total amounts of nutrients present in the residues of
E. dunnii, then in
E. grandis and, finally, in
E. globulus were associated, in part, with the differences in the amounts of biomass at the time of harvest (
Table 2), and, in addition, with the differences in the concentrations that for all nutrients occurred in residues, generally greater in
E. dunnii, intermediate in
E. grandis and lesser in
E. globulus [
32]. The soils, despite having some differences in depth and fertility, were relatively homogeneous (acid pH and low to medium natural fertility), with little influence on the absorption of nutrients.
The absolute amounts recycled appeared, due to the aforementioned (higher total amount in E. dunnii residues), in the same order for all the nutrients (E. dunnii > E. grandis > E. globulus); however, they varied when analyzed at the same percentage. This was explained because the release patterns depended on the nutrient and the parts, which, in turn, differed in their decomposition according to the species.
The high release of K (similar in the three species) can be explained because this nutrient is not part of the organic structures in plants but is found in ionic form (K
+) developing osmotic functions. This characteristic consequently determines its easiness of dissolution and removal via rainwater, regardless of biological factors [
39]. Similar results were obtained by [
7,
31,
37].
The Mg release frequently accompanies the loss of biomass of the residues [
39], although the amount released in the present investigation was proportionally greater, explained by the lower concentrations of Mg in the different parts towards the end of the study. Rocha et al. [
7] for
E. grandis (Sao Paulo, Brazil) reported that 65% of the Mg remained in the residues at the end of the evaluation (10 months post-harvest), a percentage similar to that found in the present investigation for the three species 12 months after harvest. As mentioned above, the addition of all nutrients at the time of the new planting in the study carried out in Brazil was able to accelerate the decomposition process and reach the same remaining percentage 2 months before our study.
In P, a slight decrease in concentration was observed at 2 years, associated with the loss of soluble compounds, resulting in a slightly higher recycled percentage 2 years after harvest compared to the loss of biomass. For this nutrient, the higher percentage release in
E. dunnii compared to
E. grandis and
E. globulus was explained by the greater biomass loss that occurred in the
E. dunnii leaf part (83%) and that, in this part, the highest concentrations was recorded P. Rocha et al. [
7] found in
E. grandis that 45% of the P remained in the residues at 10 months post-harvest, a percentage lower than in our research for the same species at 12 months (63%). The lower half-life for the leaf part reported by these authors compared to our research explains these differences.
Regarding the N, the low overall release from the residues (33% returned to the site in 2 years) was explained because some parts retained or even immobilized mineral N from the soil. For the three species, the net immobilization of N was recorded in the bark part. In addition, immobilization also occurred in the thick branches part of
E. grandis and
E. globulus, the latter having the highest C:N ratio (
Table 3). In all cases, it was observed that concentrations were higher after 2 years of harvest (unpublished data). Similar immobilization behavior in woody components has been reported by [
9,
10,
15].
As for P, the greater loss of biomass occurred in the leaf part of
E. dunnii, explaining the highest percentage of N release in this species, since it showed the highest concentrations of N. In turn, a slightly higher percentage of N was released in
E. grandis compared to
E. globulus, despite the lower loss of leaf biomass in this species (57% and 68% at 2 years for
E. grandis and
E. globulus, respectively), which could be explained because in
E. grandis the amount of this nutrient in the leaves was almost two thirds of the total present in the residues, while in the other two species it was slightly less than 50%. Rocha et al. [
7] indicated that 70% of the total N remained in the residues of
E. grandis at 10 months post-harvest, a percentage lower than that found for the same species at 12 months in this research (83%). The differences can be explained, as for the P, by the lower half-life of the leaf part found by these authors and the higher concentration of N present in this part in relation to the woody residues.
Regarding Ca, all parts, in the three species, maintained or increased their concentrations in the 2 years, so its loss was lower compared to that of biomass, and this explained the high contents in the residues after 24 months. This nutrient is found in vegetables as a constituent of the cell wall, a structural function [
40], which explains the behavior found, since soluble nutrients are released first. On average across the species, only 29% of the total returned to the soil in the evaluation period, decreasing significantly only in the leaf part. The lowest cycling occurred in
E. dunnii, which was explained by its bark being the part of greater durability of all those analyzed (
Table 3) and considering that this part presented the highest concentration of this nutrient in all species. Rocha et al. [
7] reported that 76% of the total Ca remained in the residues of
E. grandis at 10 months post-harvest, a similar percentage to that found for the same species at 12 months in the present study.
Although for P and cations (K, Ca and Mg) there are few loss mechanisms that occur during the decomposition process and, therefore, their reuse efficiency is very high, there are mechanisms of absolute N losses (passage to gaseous forms and leaching of soluble inorganic forms, such as nitrate anion) and relative losses (immobilization in organic forms). Therefore, for N, it is not possible to ensure that the recycled total will be fully available for the next planting [
41].
In Uruguay, commercial plantations only include N and P in the fertilization schemes at the time of planting; therefore, it is particularly interesting to estimate for the cations (K, Ca and Mg) the contribution from the cycling in comparison to their stocks in the A horizon of the soils. Considering the soil mass present in the A horizon (depending on its depth and apparent density) and the concentration of cations under soluble and exchangeable forms (without taking into account non-exchangeable forms or geochemical recycling), K stands out as the nutrient whose contribution via this route presented a greater proportion. Considering the differences observed between the species, as well as the different stocks according to the soil, at 2 years of harvest, the return from the residues in relation to the stocks present in the soil accounted for between 23 to 81% for K, 4 to 19% for Ca and 5 to 16% for Mg.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}