
Metabolome and Transcriptome Analysis Reveals the Effects of Host Shift on Dendrolimus houi Lajonquière Larvae

Abstract
:1. Introduction
2. Materials and Methods
2.1. The Tested Insects
2.2. Sample Preparation for HPLC-Mass
2.3. Metabolite Profiling
2.4. Qualitative and Quantitative Analyses of Metabolites
2.5. RNA Extraction and Transcriptome Sequencing Analysis
2.6. Combined Analysis of Metabolome and Transcriptome
2.7. Data Processing and Analysis
3. Results
3.1. The Effects of Host Shift on the Growth and Development of D. houi Larvae
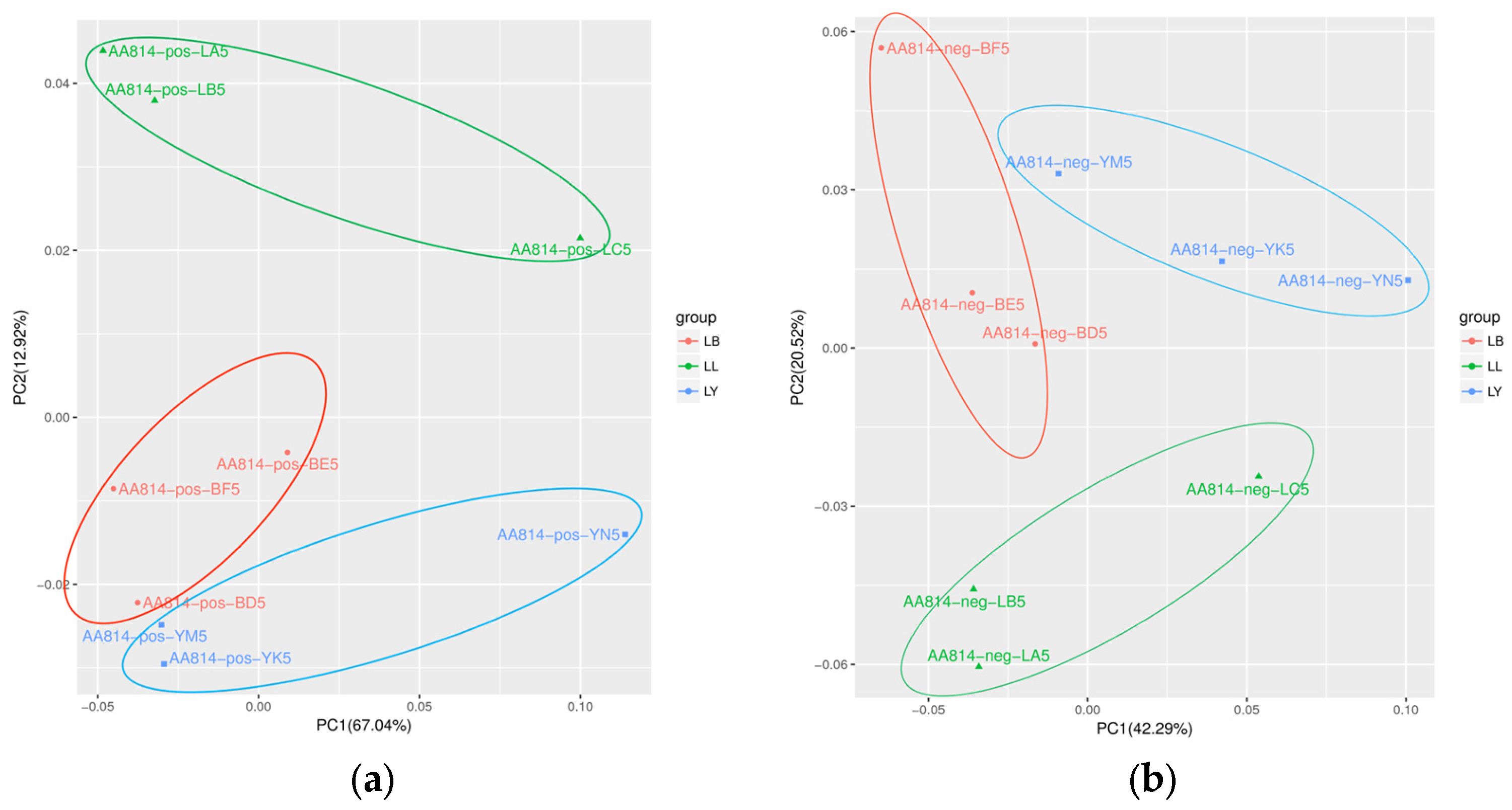
3.2. The Effects of Host Shift on the Metabolome of D. houi Larvae
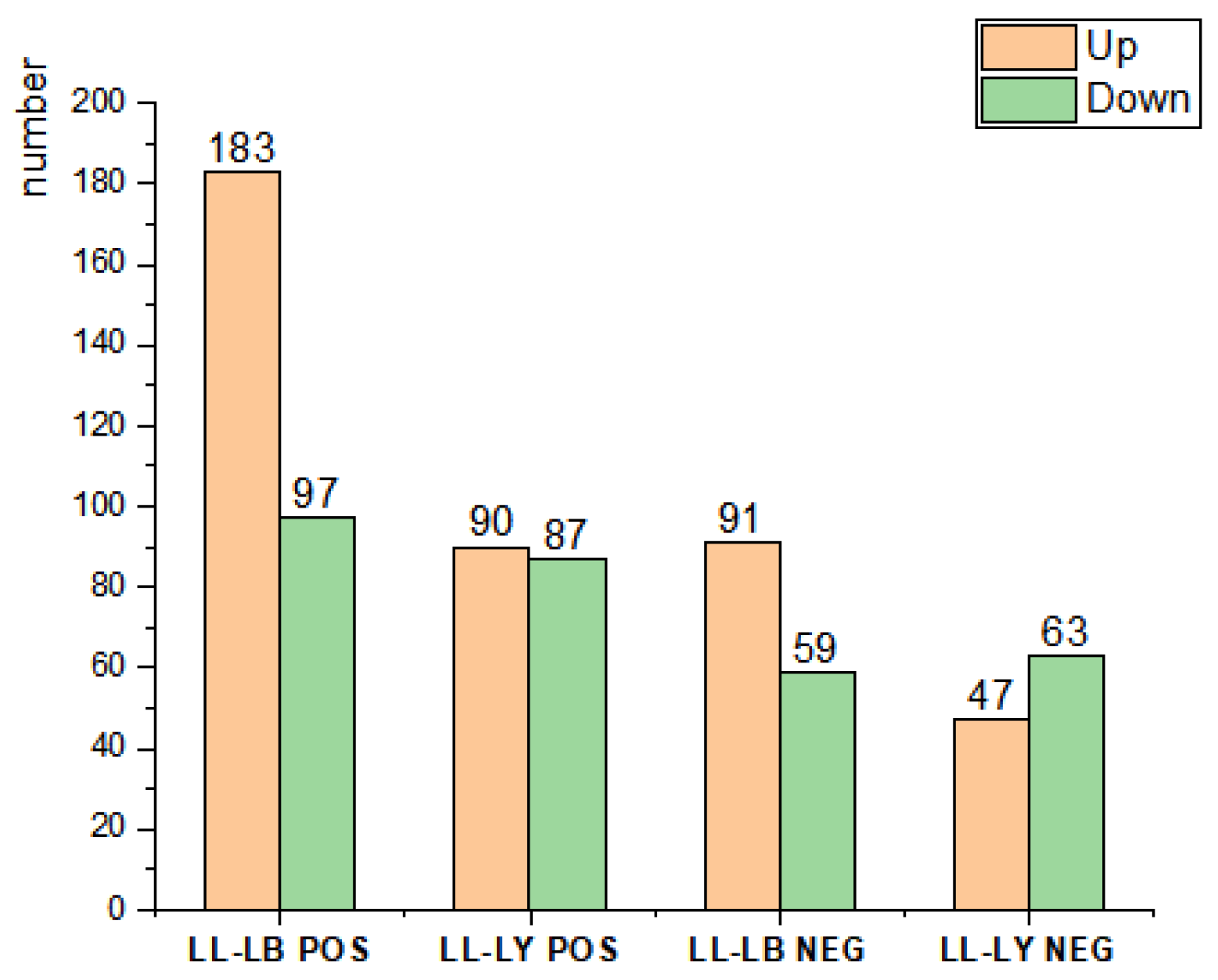
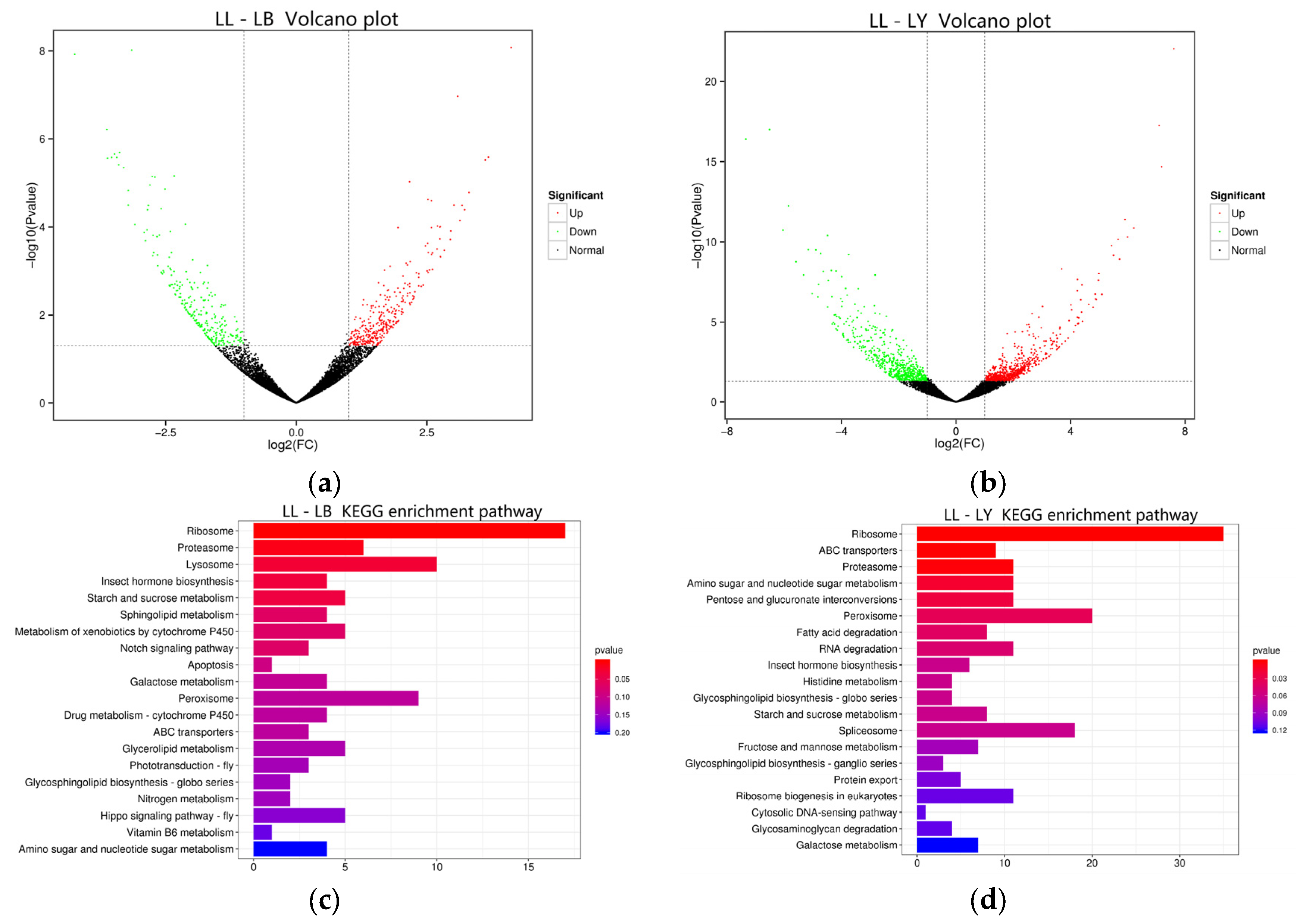
3.3. The Effects of Host Shift on the Transcriptome of D. houi Larvae
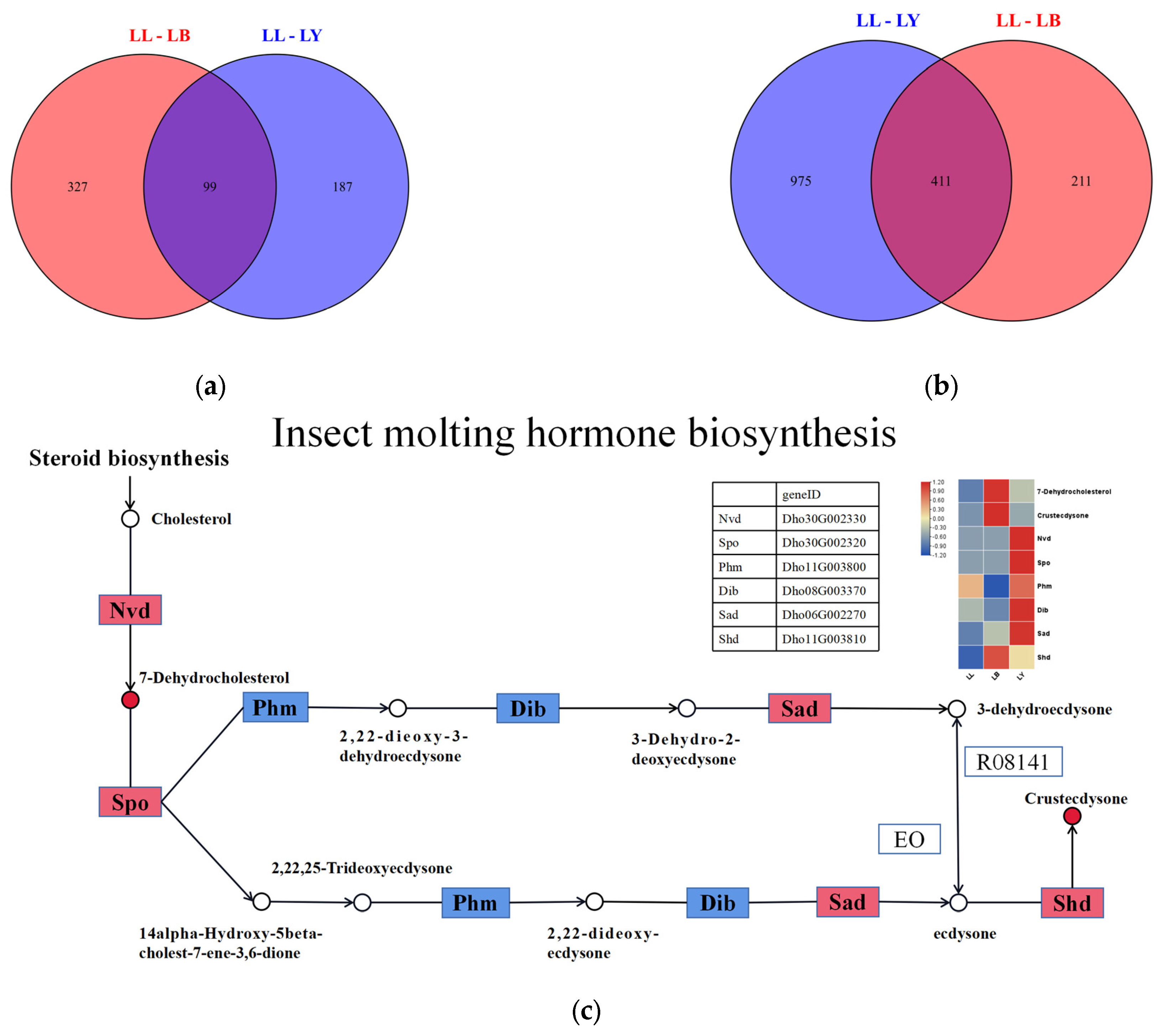
3.4. Integrated Metabolome and Transcriptome Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Hemati, S.; Naseri, B.; Ganbalani, G.N.; Dastjerdi, H.R.; Golizadeh, A. Effect of different host plants on nutritional indices of the pod borer, Helicoverpa armigera. J. Insect Sci. 2012, 12, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, M.D.; McNeil, J.N. Host-plant quality influences diapause and voltinism in a polyphagous insect herbivore. Ecology 1997, 78, 977–986. [Google Scholar] [CrossRef]
- Klemola, N.; Klemola, T.; Rantala, M.J.; Ruuhola, T. Natural host-plant quality affects immune defence of an insect herbivore. Entomol. Exp. Appl. 2007, 123, 167–176. [Google Scholar] [CrossRef]
- Vogelweith, F.; Thiéry, D.; Quaglietti, B.; Moret, Y.; Moreau, J. Host plant variation plastically impacts different traits of the immune system of a phytophagous insect. Funct. Ecol. 2011, 25, 1241–1247. [Google Scholar] [CrossRef]
- Zhang, D.W.; Xiao, Z.J.; Zeng, B.P.; Li, K.; Tang, Y.L. Insect behavior and physiological adaptation mechanisms under starvation stress. Front. Physiol. 2019, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; Yi, W.; Li, Q.; Hu, H.Y. Research progress on Monolepta hieroglyphica (Motschulsky). China Plant Prot. 2016, 36, 19–26. [Google Scholar]
- Wang, Z.H.; Wang, H.F.; Liu, X.; Wang, Y.W.; Zhang, H.Q. Population dynamics of Monolepta hieroglyphica (Motschulsky) in agro-ecosystem of adjacent crop fields. Plant Prot. 2018, 44, 161–165+178. [Google Scholar] [CrossRef]
- Wang, Y.L.; Wang, H.X.; Wang, X.P.; Wang, J.Y.; Li, C.R.; Wang, Z.L. Effect of Host Switching after Emergency on the Longevity and Egg Laying of Henosepilachna vigintioctopunctata Adults. J. Yangtze Univ. (Nat. Sci. Ed.) 2019, 16, 63–66+6. [Google Scholar] [CrossRef]
- Qiu, R.; Li, C.J.; Wang, H.T.; Chen, Y.G.; Li, X.J.; Zhang, Y.H.; Liu, D.S.; Li, H.J. Studies on Host Conversion Regularity of Myzus persicae (Sulzer) in Early Spring. J. Henan Agric. Sci. 2017, 46, 86–90. [Google Scholar] [CrossRef]
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.L.; Zhang, J.; Shen Tu, X.P.; Hao, P.Y.; Pang, K.; Yu, X.P. Review of how secondary metabolites defend plants against herbivorous insects. Chinese J. Appl. Entomol. 2022, 59, 969–978. [Google Scholar]
- Zhang, Y.B.; Lou, Y.G. Research progress in chemical interactions between plants and phytophagous. Chin. J. Appl. Ecol. 2020, 21, 2151–2160. [Google Scholar] [CrossRef]
- Smith, C.M. Plant Resistance to Insects, 1st ed.; CRC Press: Boca Raton, FL, USA, 1999; pp. 171–208. [Google Scholar]
- Gen, Y.S.; Zhao, X.D.; Han, Y.Y.; Qiao, H.; Hao, D.J. Effects of host switch on the development and digestive enzyme activities of Hyphantria cunea (Lepidoptera:Erebidae) larvae. Acta Entomol. Sin. 2022, 65, 312–321. [Google Scholar] [CrossRef]
- Lu, H.; Yang, P.; Xu, Y.; Luo, L.; Zhu, J.; Cui, N.; Kang, L.; Cui, F. Performances of survival, feeding behavior and gene expression in aphids reveal their different fitness to host alteration. Sci. Rep. 2016, 6, srep19344. [Google Scholar] [CrossRef] [Green Version]
- Pankoke, H.; Bowers, M.D.; Dobler, S. Influence of iridoid glycoside containing host plants on midgut β-glucosidase activity in a polyphagous caterpillar, Spilosoma virginica Fabricius (Arctiidae). J. Insect Physiol. 2010, 56, 1907–1912. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.L. Research Advances on Dendrolimus houi in Yunnan Province. J. Southwest For. Coll. 2008, 3, 42–44+50. [Google Scholar]
- Liu, X.Z. Study on Biological Characteristics and Its Integrated Control of Dendrolimus latipenis. J. Southwest For. Coll. 2006, 3, 52–54+58. [Google Scholar]
- Luo, S.G.; Xu, Z.W.; Zhao, R.Y.; Hua, B.S.; Lin, S.B. Biological Properties and Control of Dendrolimus houi. J. Zhejiang For. Sci. Technol. 1998, 1, 11–15. [Google Scholar]
- Yi, A.L.; Chuan, R.F.; Xu, G.L. A Study on the Biological Characteristics of Dendrolimus houi. J. Southwest For. Coll. 2002, 4, 53–55. [Google Scholar]
- Bi, Z.N. Major Pest Control of Pinus yunnanensis in Yunnan. Shaanxi For. Sci. Technol. 2014, 3, 111–114. [Google Scholar]
- Li, W.; Zhong, G.H. Occurrence and Prevention and Control of Dendrolmus houi lajonquere. Ningxia J. Agric. For. Sci. Technol. 2013, 54, 76–77. [Google Scholar]
- Hua, Y.; Lu, C.D.; Shi, Y.F.; He, H.; Han, X.H.; Lin, H.Y.; Zhang, F.P.; Liang, G.H. Research Advances in Dendrolimus houi Lajonquiere of China. World For. Res. 2019, 32, 62–68. [Google Scholar] [CrossRef]
- Valdes, A.M.; Glass, D.; Spector, T.D. Omics technologies and the study of human ageing. Nat. Rev. Genet. 2013, 14, 601–607. [Google Scholar] [CrossRef]
- Cavill, R.; Jennen, D.; Kleinjans, J.; Briedé, J.J. Transcriptomic and metabolomic data integration. Brief. Bioinform. 2016, 17, 891–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.Y.; Liang, Y.; Chen, Z.H.; Hu, Y.; Li, C.L.; Zhang, Z.H.; Weng, H.F.; Huang, B.; Zhang, F.P.; Liang, G.H. Effects of Different Host Plants on the Growth, Reproduction and Physiological Enzyme Activity of Dendrolimus houi Lajonquiere (Lepidoptera: Lasiocampidae). For. Res. 2022, 35, 63–70. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, D.; Meng, Z.J.; Yan, S.C. Adaptability of Larval Growth and Development in Hyphantria cunea to Different Host Plant Secondary Metabo. J. Northeast. For. Univ. 2020, 48, 100–104. [Google Scholar] [CrossRef]
- Zhu, Y.; Gao, H.; Han, S.; Li, J.; Wen, Q.; Dong, B. Antidiabetic activity and metabolite profiles of ascidian Halocynthia roretzi. J. Funct. Foods 2022, 93, 105095. [Google Scholar] [CrossRef]
- Li, S.L.; Song, J.Z.; Choi, F.F.; Qiao, C.F.; Zhou, Y.; Han, Q.B.; Xu, H.X. Chemical profiling of Radix Paeoniae evaluated by ultra-performance liquid chromatography/photo-diode-array/quadrupole time-of-flight mass spectrometry. J. Pharm. Biomed. Anal. 2009, 49, 253–266. [Google Scholar] [CrossRef]
- Bocca, C.; Kane, M.S.; Veyrat-Durebex, C.; Chupin, S.; Alban, J.; Nzoughet, J.K.; Le Mao, M.; de la Barca, J.M.C.; Amati-Bonneau, P.; Bonneau, D.; et al. The metabolomic bioenergetic signature of opa1-disrupted mouse embryonic fibroblasts highlights aspartate deficiency. Sci. Rep. 2018, 8, 11528. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Li, L.; Gan, J.; Peng, C.; Wang, X.; Chen, W.; Peng, D. Identification and functional analysis of circular RNAs induced in rats by middle cerebral artery occlusion. Gene 2019, 701, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Wang, B.; He, G.; Wang, Y.; Tang, X.; Wang, S.; Ma, Y.; Fu, C.; Chai, G.; Zhou, G. Metabolomics Integrated with Transcriptomics Reveals Redirection of the Phenylpropanoids Metabolic Flux in Ginkgo biloba. J. Agric. Food Chem. 2019, 67, 3284–3291. [Google Scholar] [CrossRef] [PubMed]
- Rohfritsch, O. A “defense” mechanism of Picea excelsa L. against the gall former Chermes abietis L. (Homoptera, Adelgidae). Z. Für Angew. Entomol. 1981, 92, 18–26. [Google Scholar] [CrossRef]
- Stotz, H.U.; Kroymann, J.; Mitchell-Olds, T. Plant-insect interactions. Curr. Opin. Plant Biol. 1999, 2, 268–272. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 1st ed.; Oxford University Press on Demand: New York, NY, USA, 2005. [Google Scholar]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hua, Y.; Fang, X.Y.; Zhang, Z.H.; Yan, J.W.; Liang, G.H. Selectivity and fitness of Dendrolimus houi on different host plants. J. For. Environ. 2023, 43, 217–223. [Google Scholar] [CrossRef]
- Montanaro, L.; Treré, D.; Derenzini, M. Nucleolus, ribosomes, and cancer. Am. J. Pathol. 2008, 173, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Bohnsack, K.E.; Bohnsack, M.T. Uncovering the assembly pathway of human ribosomes and its emerging links to disease. EMBO J. 2019, 38, e100278. [Google Scholar] [CrossRef]
- Ramakrishnan, V. Ribosome structure and the mechanism of translation. Cell 2002, 108, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Kapahi, P.; Zid, B.M.; Harper, T.; Koslover, D.; Sapin, V.; Benzer, S. Regulation of Lifespan in Drosophila by Modulation of Genes in the TOR Signaling Pathway. Curr. Biol. 2004, 14, 885–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, K.Z.; Palter, J.E.; Rogers, A.N.; Olsen, A.; Chen, D.; Lithgow, G.J.; Kapahi, P. Inhibition of mRNA translation extends lifespan in Caenorhabditis elegans. Aging Cell 2007, 6, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molenaars, M.; Janssens, G.E.; Williams, E.G.; Jongejan, A.; Lan, J.; Rabot, S.; Joly, F.; Moerland, P.D.; Schomakers, B.V.; Lezzerini, M.; et al. A conserved mito-cytosolic translational balance links two longevity pathways. Cell Metab. 2020, 31, 549–563.e7. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.E. A role for the ribosome in development. Trends Plant Sci. 2009, 14, 512–519. [Google Scholar] [CrossRef]
- Niwa, Y.S.; Niwa, R. Transcriptional regulation of insect steroid hormone biosynthesis and its role in controlling timing of molting and metamorphosis. Dev. Growth Differ. 2016, 58, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Connacher, R.P.; O’connor, M.B. Control of the insect metamorphic transition by ecdysteroid production and secretion. Curr. Opin. Insect Sci. 2020, 43, 11–20. [Google Scholar] [CrossRef]
- Meister, M.; Richards, G. Ecdysone and insect immunity: The maturation of the inducibility of the diptericin gene in Drosophila larvae. Insect Biochem. Mol. Biol. 1996, 26, 155–160. [Google Scholar] [CrossRef]
- Belles, X.; Piulachs, M.-D. Ecdysone signalling and ovarian development in insects: From stem cells to ovarian follicle formation. Biochim. Biophys. Acta (BBA)—Gene Regul. Mech. 2015, 1849, 181–186. [Google Scholar] [CrossRef]
- Swevers, L.; Iatrou, K. The ecdysone regulatory cascade and ovarian development in lepidopteran insects: Insights from the silkmoth paradigm. Insect Biochem. Mol. Biol. 2003, 33, 1285–1297. [Google Scholar] [CrossRef]
- Agui, N.; Hiruma, K. Ecdysone as a feedback regulator for the neurosecretory brain cells in Mamestra brassicae. J. Insect Physiol. 1977, 23, 1393–1396. [Google Scholar] [CrossRef]
- Mizoguchi, A.; Kamimura, M.; Kiuchi, M.; Kataoka, H. Positive feedback regulation of prothoracicotropic hormone secretion by ecdysteroid—A mechanism that determines the timing of metamorphosis. Insect Biochem. Mol. Biol. 2015, 58, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Robbins, W.E.; Kaplanis, J.N.; Thompson, M.J.; Shortino, T.J.; Cohen, C.F.; Joyner, S.C. Ecdysones and analogs: Effects on development and reproduction of insects. Science 1968, 161, 1158–1160. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.L.; Zhang, J.Y.; Wang, D.; Zhao, Y.M.; Han, X.L.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone binds to dopamine receptor to repress lepidopteran insect feeding and promote pupation. PLoS Genet. 2019, 15, e1008331. [Google Scholar] [CrossRef] [Green Version]
- Nijhout, H.F.; Riddiford, L.M.; Mirth, C.; Shingleton, A.W.; Suzuki, Y.; Callier, V. The developmental control of size in insects. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 113–134. [Google Scholar] [CrossRef]
- Shafiei, M.; Moczek, A.P.; Nijhout, H.F. Food availability controls the onset of metamorphosis in the dung beetle Onthophagus taurus (Coleoptera: Scarabaeidae). Physiol. Entomol. 2001, 26, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Kannangara, J.R.; Mirth, C.K.; Warr, C.G. Regulation of ecdysone production in Drosophila by neuropeptides and peptide hormones. Open Biol. 2021, 11, 200373. [Google Scholar] [CrossRef] [PubMed]
- Davidowitz, G.; D’Amico, L.J.; Nijhout, H.F. The effects of environmental variation on a mechanism that controls insect body size. Evol. Ecol. Res. 2004, 6, 49–62. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instars | C. fortunei–C. fortunei (LL) | C. fortunei–C. funebris (LB) | C. fortunei–P. yunnanensis (LY) | |
|---|---|---|---|---|
| body length/mm | 4th | 46.09 ± 2.88 a | 48.95 ± 4.10 a | 48.30 ± 2.89 a |
| 6th | 90.93 ± 5.18 a | 73.27 ± 13.18 b | 67.26 ± 10.60 b | |
| body weight/g | 4th | 0.77 ± 0.12 b | 0.80 ± 0.27 b | 0.94 ± 0.24 a |
| 6th | 6.28 ± 1.13 a | 3.54 ± 1.56 b | 2.84 ± 1.58 b | |
| head shell width/mm | 4th | 4.03 ± 0.19 a | 4.10 ± 0.21 a | 4.15 ± 0.21 a |
| 6th | 6.97 ± 0.11 a | 6.52 ± 0.61 b | 5.80 ± 0.79 b | |
| development duration/d | 4–6th | 32.00 ± 0.34 b | 37.78 ± 0.87 a | 38.11 ± 1.13 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, X.; Chen, Z.; Chen, Z.; Chen, J.; Zhao, Z.; Wu, P.; Wu, H.; Zhang, F.; Liang, G. Metabolome and Transcriptome Analysis Reveals the Effects of Host Shift on Dendrolimus houi Lajonquière Larvae. Forests 2023, 14, 1307. https://doi.org/10.3390/f14071307
Fang X, Chen Z, Chen Z, Chen J, Zhao Z, Wu P, Wu H, Zhang F, Liang G. Metabolome and Transcriptome Analysis Reveals the Effects of Host Shift on Dendrolimus houi Lajonquière Larvae. Forests. 2023; 14(7):1307. https://doi.org/10.3390/f14071307
Chicago/Turabian StyleFang, Xinyuan, Zhenghao Chen, Zhenhong Chen, Jian Chen, Zhenhui Zhao, Peilin Wu, Hongmin Wu, Feiping Zhang, and Guanghong Liang. 2023. "Metabolome and Transcriptome Analysis Reveals the Effects of Host Shift on Dendrolimus houi Lajonquière Larvae" Forests 14, no. 7: 1307. https://doi.org/10.3390/f14071307