

Identifying Plant Physiological and Climatic Drivers in the Woody Shrub Prosopis strombulifera: Effects of Spring Traits on Flower Sprouting and Fruit Production

Abstract
:1. Introduction
2. Materials and Methods
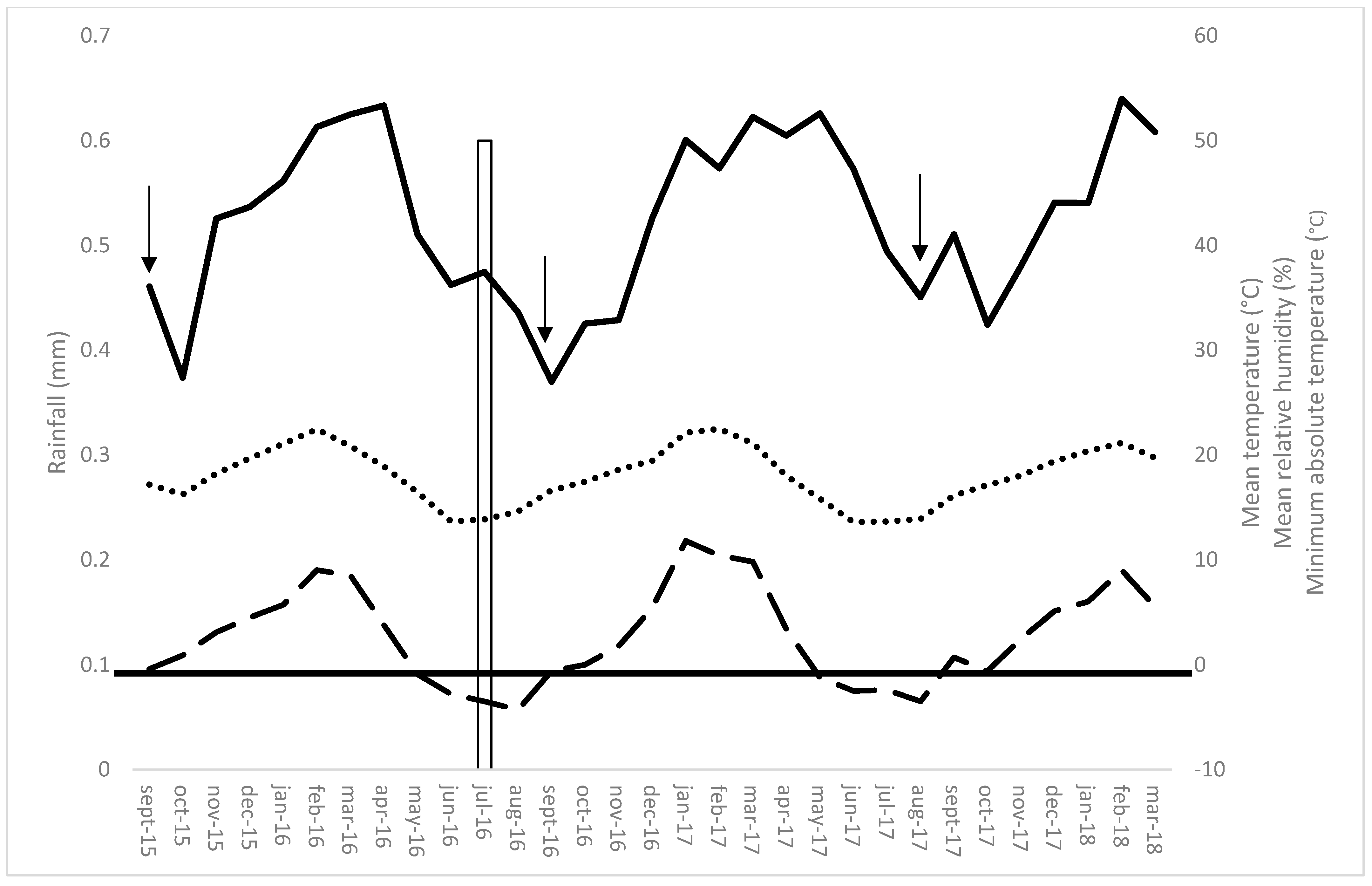
2.1. Study Area
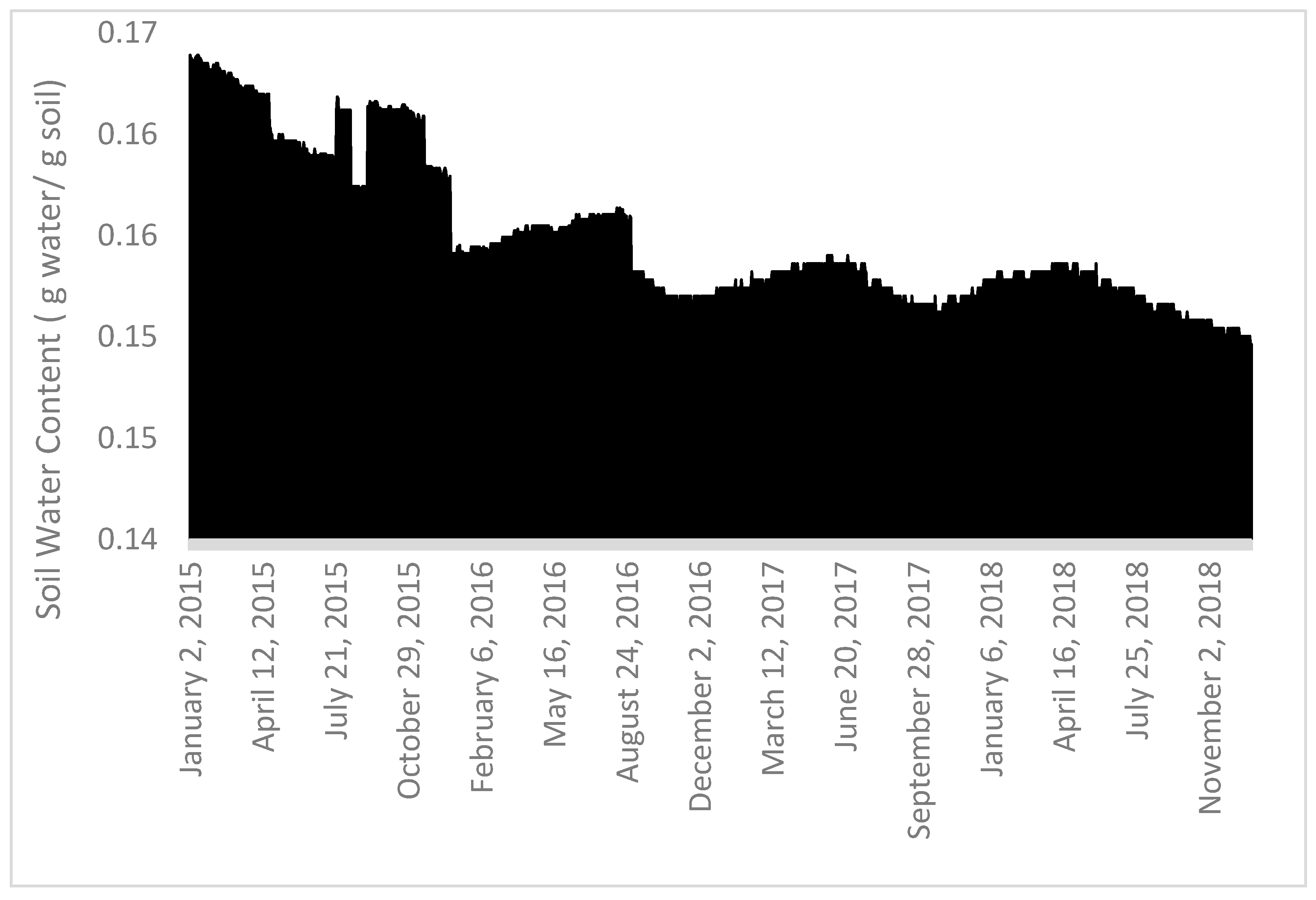
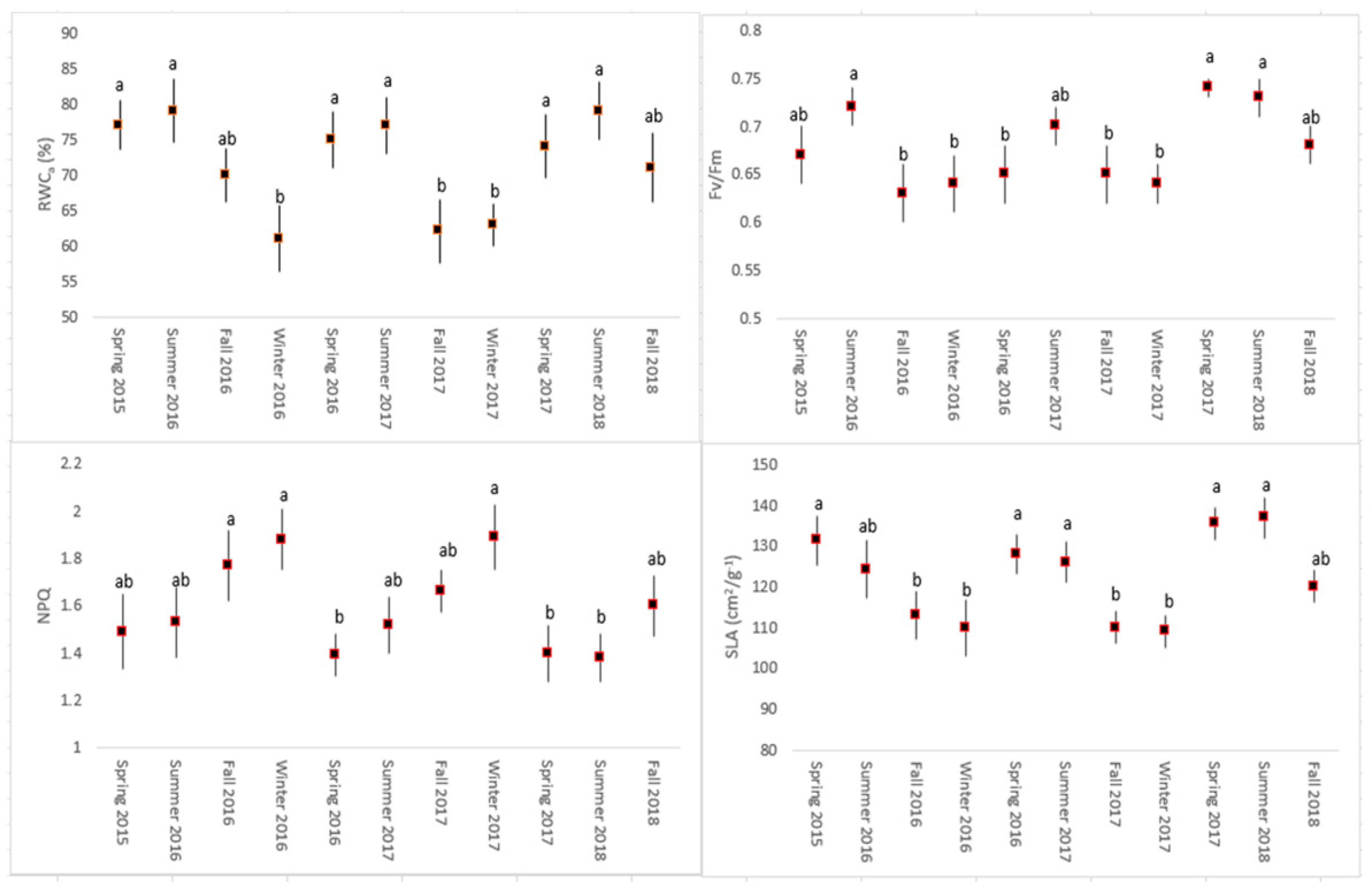
2.2. Water Parameters and Leaf Traits
2.3. Photosynthetic Efficiency
2.4. Flower Sprouting and Pod Tagging
2.5. Fruit Production
2.6. Data Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gutiérrez, A.; Barbosa, O.; Christie D del Val, E.; Ewing, H.; Jones, C.; Marquet, P.; Weathers, K.; Armesto, J. Regeneration patterns and persistence of the fog-dependent Fray Jorge forest in semiarid Chile during the past two centuries. Glob. Chang. Biol. 2008, 14, 161–176. [Google Scholar] [CrossRef]
- Squeo, F.; Rada, F.; García, C.; Ponce, M.; Rojas, A.; Azócar, A. Cold resistance mechanisms in high desert Andean plants. Oecologia 1996, 105, 552–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angert, A.L.; Horst, J.; Huxman, T.; Venable, D. Phenotypic plasticity and precipitation response in Sonoran Desert winter annuals. Am. J. Bot. 2010, 97, 405–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carevic, F.; Delatorre, J.; Arenas, A. Physiological strategies during frost periods for two populations of Prosopis burkartii, an endangered species endemic to the Atacama desert. J. Arid Environ. 2015, 114, 79–83. [Google Scholar] [CrossRef]
- Pesendorfer, M.; Bogdziewicz, M.; Szymkowiak, J.; Borowski, Z.; Kantorowicz, W.; Espelta, J.; Fernández-Martinez, M. Investigating the relationship between climate, stand age, and temporal trends in masting behavior of European forest trees. Glob. Chang. Biol. 2020, 26, 1654–1667. [Google Scholar] [CrossRef] [Green Version]
- Hacket-Pain, A.; Foest, J.J.; Pearse, I.S.; LaMontagne, J.M.; Koenig, W.D.; Vacchiano, G.; Bogdziewicz, M.; Caignard, T.; Celebias, P.; van Dormolen, J.; et al. MASTREE+: Time-series of plant reproductive effort from six continents. Global Chang. Biol, 2022; Epub ahead of print. [Google Scholar] [CrossRef]
- McRostie, V.B.; Gayo, E.; Santoro, C.; De Pol-Holz, R.; Latorre, C. The pre-Columbian introduction and dispersal of Algarrobo (Prosopis, Section Algarobia) in the Atacama Desert of northern Chile. PLoS ONE 2017, 12, e0181759. [Google Scholar] [CrossRef] [Green Version]
- Burkart, A. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). J. Arnold Arbor. 1976, 57, 450–525. [Google Scholar] [CrossRef]
- Rogers, K.E. The Magnificent Mesquite, 1st ed.; University of Texas Press: Austin, TX, USA, 2000; p. 167. [Google Scholar]
- Contreras, R.; Carevic, F.; Porcile, V.; Aburto, M. Development of SSR loci in Prosopis tamarugo Phillipi and assessment of their transferability to species of the Strombocarpa section. For. Syst. 2020, 29, 108–117. [Google Scholar] [CrossRef]
- Chávez, R.; Jan, G.; Clevers, W.; Herold, M.; Acevedo, E.; Ortiz, E. Assessing water stress of desert tamarugo trees using in situ data and very high spatial resolution remote sensing. Remote Sens. 2013, 5, 5064–5088. [Google Scholar] [CrossRef] [Green Version]
- Viguier, B.; Jourde, H.; Yáñez, G.; Lira, E.; Léonardi, V.; Moya, C.; Pérez, T.; Maringue, T.; Lictevout, E. Multidisciplinary study for the assessment of the geometry, boundaries and preferential recharge zones of an overexploited aquifer in the Atacama Desert (Pampa del Tamarugal, Northern Chile). J. S. Am. Earth Sci. 2018, 86, 366–383. [Google Scholar] [CrossRef]
- Carevic, F.; Alarcón, E.; Villacorta, A. Is the visual survey method effective for measuring fruit production in Prosopis tree species? Rangel. Ecol. Manag. 2021, 74, 119–124. [Google Scholar] [CrossRef]
- Corcuera, L.; Camarero, J.; Gil-Pelegrin, E. Functional groups in Quercus species derived from the analysis of pressure volume curves. Trees 2002, 16, 465–472. [Google Scholar] [CrossRef]
- Robichaux, R.H. Variation in the tissue water relations of two sympatric Hawaiian Dubautia species and their natural hybrid. Oecologia 1984, 65, 75–81. [Google Scholar] [CrossRef]
- Guisande-Collazo, A.; González, L.; Souza-Alonso, P. Impact of an invasive nitrogen-fixing tree on arbuscular mycorrhizal fungi and the development of native species. AoB Plants 2016, 8, plw018. [Google Scholar] [CrossRef] [Green Version]
- Leport, L.; Turner, N.; Davies, S.; Siddique, K. Variation in pod production and abortion among chickpea cultivars under terminal drought. Eur. J. Agron. 2006, 24, 236–246. [Google Scholar] [CrossRef]
- Greenberg, C.H. Individual variation in acorn production by five species of Southern Appalachian oaks. For. Ecol. Manag. 2000, 132, 199–210. [Google Scholar] [CrossRef]
- Sokal, R.; Rohlf, F. Biometry, 4th ed.; W. H. Freeman and Co.: New York, NY, USA, 2012. [Google Scholar]
- Kachout, S.; Ben Mansour, A.; Hamza, K.; Leclerc, J.; Rejeb, M.; Ouerghi, Z. Leaf–water relations and ion concentrations of the halophyte Atriplex hortensis in response to salinity and water stress. Acta Physiol. Plant. 2011, 33, 335–342. [Google Scholar] [CrossRef]
- Garrido, M.; Bown, H.; Ayamante, J.; Orell, M.; Sánchez, A.; Acevedo, E. The adjustment of Prosopis tamarugo hydraulic architecture traits has a homeostatic effect over its performance under descent of phreatic level in the Atacama Desert. Trees 2020, 34, 89–99. [Google Scholar] [CrossRef]
- Carevic, F.; Delatorrre, J.; Carrasco, A. Plant water variable and reproductive traits are influenced by seasonal climatic variables in Prosopis burkartii (Fabaceae) at Northern Chile. Flora 2017, 233, 7–11. [Google Scholar] [CrossRef]
- Traversari, S.; Neri, A.; Traversi, M.L.; Giovannelli, A.; Francini, A.; Sebastiani, L. Daily Osmotic Adjustments in Stem May Be Good Predictors of Water Stress Intensity in Poplar. Plant Physiol. Biochem. 2020, 146, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Rundel, O.; Dillon, O.; Palma, B.; Mooney, A.; Gulmon, S.; Ehleringer, J. The phytogeography and ecology of the coastal Atacama and Peruvian Desert. Aliso 1991, 13, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Cereceda, P.; Larraín, H.; Osses, P.; Farías, M.; Egaña, I. The climate of the coast and fog zone in the Tarapacá Region, Atacama Desert, Chile. Atmos. Res. 2008, 87, 301–311. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Ehleringer, J.; Rundel, P.; Palma, B.; Mooney, H. Carbon isotope ratios of Atacama Desert plants reflect hyperaridity of region in northern Chile. Rev. Chil. Hist. Nat. 1998, 71, 79–86. [Google Scholar]
- Gong, X. Tradeoffs between nitrogen- and water-use efficiency in dominant species of the semiarid steppe of Inner Mongolia. Plant Soil 2011, 340, 227–238. [Google Scholar] [CrossRef]
- Carvalho, M.; Castro, P.; Ferraz, J.; Mendes, A. Are plant growth retardants a strategy to decrease lodging and increase yield of sunflower? Comun. Sci. 2016, 7, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Hamerlynck, E.P.; Huxman, T. Ecophysiology of two Sonoran Desert evergreen shrubs during extreme drought. J. Arid Environ. 2019, 73, 582–585. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 13, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.H.; Adams, W.; Demmig-Adams, B.; Logan, B.; Verhoven, A.; Smith, S. Nocturnally retained zeaxanthin does not remain engaged in a state primed for energy dissipation during the summer in two Yucca species growing in the Mojave Desert. Plant Cell Environ. 2002, 25, 95–103. [Google Scholar] [CrossRef]
- Aravena, R.; Acevedo, E. Estudio de la relación hídrica de Prosopis tamarugo Phil. mediante isótopos estables, oxígeno-18 y deuterio. (Study of the water relations of Prosopis Tamarugo Phil using stable isotopes, oxigen-18 and deuterium). In Estado Actual Sobre el Conocimiento de Prosopis Tamarugo, Arica, Chile, 11–15 de Junio de 1984; Mario, H., Ed.; FAO: Rome, Italy, 1985; 483p. [Google Scholar]
- McKay, C.P.; Friedmann, E.; Gomez-Silva, B.; Caceres, L.; Andersen, D.; Landheim, R. Temperature and moisture conditions in the extreme arid regions of the Atacama Desert: Four years of observations including the El Nino of 1997–1998. Astrobiology 2003, 3, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Calderón, G.; Garrido, M.; Acevedo, E. Prosopis tamarugo Phil.: A native tree from the Atacama Desert. Groundwater table depth thresholds for conservation. Rev. Chil. Hist. Nat. 2015, 88, 18. [Google Scholar] [CrossRef] [Green Version]
- García-Mozo, H.; Gómez-Casero, M.; Domínguez, E.; Galán, C. Influence of pollen emission and weather-related factors on variations in holm-oak (Quercus ilex subsp. ballota) acorn production. Environ. Exp. Bot. 2007, 61, 35–40. [Google Scholar] [CrossRef]
- Laouali, A.; Dan Guimbo, I.; Chaibou, I.; Mahamane, A. Fruit production of Prosopis africana (G. et Perr.) Taub., an overexploited species in the Southeastern Niger. Int. J. Curr. Microb. Appl. Sci. 2015, 4, 50–56. [Google Scholar]
- Risio, L.; Calama, R.; Bogino, S.; Bravo, F. Inter-annual variability in Prosopis caldenia pod production in the Argentinean semiarid pampas: A modelling approach. J. Arid Environ. 2016, 131, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Joseau, M.J.; Verga, A.; Díaz, M.; Julio, N. Morphological diversity of populations of the Genus Prosopis in the Semiarid Chaco of Northern Cordoba and Southern Santiago Del Estero. Am. J. Plant Sci. 2013, 4, 2092–2111. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fruit Production | Flower Sprout | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | Intrapopulation | Interannual | Intrapopulation | Interannual | ||||
| p-Value | F | Variation Coefficient | Total Fruit Biomass [g m−2] | p-Value | F | Variation Coefficient | Mean Number of Flower Sprouts | |
| 2016 | 0.23 | 0.18 | 0.65 | 169.8 a ± 6.1 | 0.12 | 1.08 | 0.55 | 26 a ± 5.3 |
| 2017 | 0.03 | 0.11 | 0.74 | 142.9 b ± 5.8 | 0.04 | 1.87 | 0.58 | 12 b ± 4.1 |
| 2018 | 0.22 | 0.19 | 0.75 | 178.1 a ± 3.4 | 0.11 | 2.99 | 0.60 | 24 a ± 5.2 |
| Parameter [n = 75] | Fruit Production | Flower Sprout | ||
|---|---|---|---|---|
| p Value | r | p Value | r | |
| Ψpredawn | 0.43 | <0.01 | 0.54 | <0.01 |
| Ψπ100 | 0.93 | <0.001 | 0.41 | <0.01 |
| Ψπ0 | 0.13 | 0.10 | 0.29 | 0.09 |
| RWC0 | 0.19 | 0.17 | 0.45 | 0.02 |
| RWCa | 0.02 | 0.55 | >0.01 | 0.58 |
| Fv/Fm | 0.09 | 0.32 | 0.09 | 0.29 |
| NPQ | 0.03 | −0.52 | 0.01 | −0.50 |
| Pq | 0.55 | <0.01 | 0.36 | <0.01 |
| SPAD | 0.88 | <0.001 | 0.43 | <0.01 |
| SLA | 0.04 | 0.48 | 0.02 | 0.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carevic, F.S.; Contreras, R.; Carmona, E.; Barrientos, E. Identifying Plant Physiological and Climatic Drivers in the Woody Shrub Prosopis strombulifera: Effects of Spring Traits on Flower Sprouting and Fruit Production. Forests 2023, 14, 1167. https://doi.org/10.3390/f14061167
Carevic FS, Contreras R, Carmona E, Barrientos E. Identifying Plant Physiological and Climatic Drivers in the Woody Shrub Prosopis strombulifera: Effects of Spring Traits on Flower Sprouting and Fruit Production. Forests. 2023; 14(6):1167. https://doi.org/10.3390/f14061167
Chicago/Turabian StyleCarevic, Felipe S., Roberto Contreras, Erico Carmona, and Ermindo Barrientos. 2023. "Identifying Plant Physiological and Climatic Drivers in the Woody Shrub Prosopis strombulifera: Effects of Spring Traits on Flower Sprouting and Fruit Production" Forests 14, no. 6: 1167. https://doi.org/10.3390/f14061167