
Fine-Root Soil Stoichiometry of Picea schrenkiana Fisch. et Mey. and Its Correlation with Soil Environmental Factors under Different Nitrogen Input Levels in the Tianshan Mountains, Xinjiang

Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Experimental Design
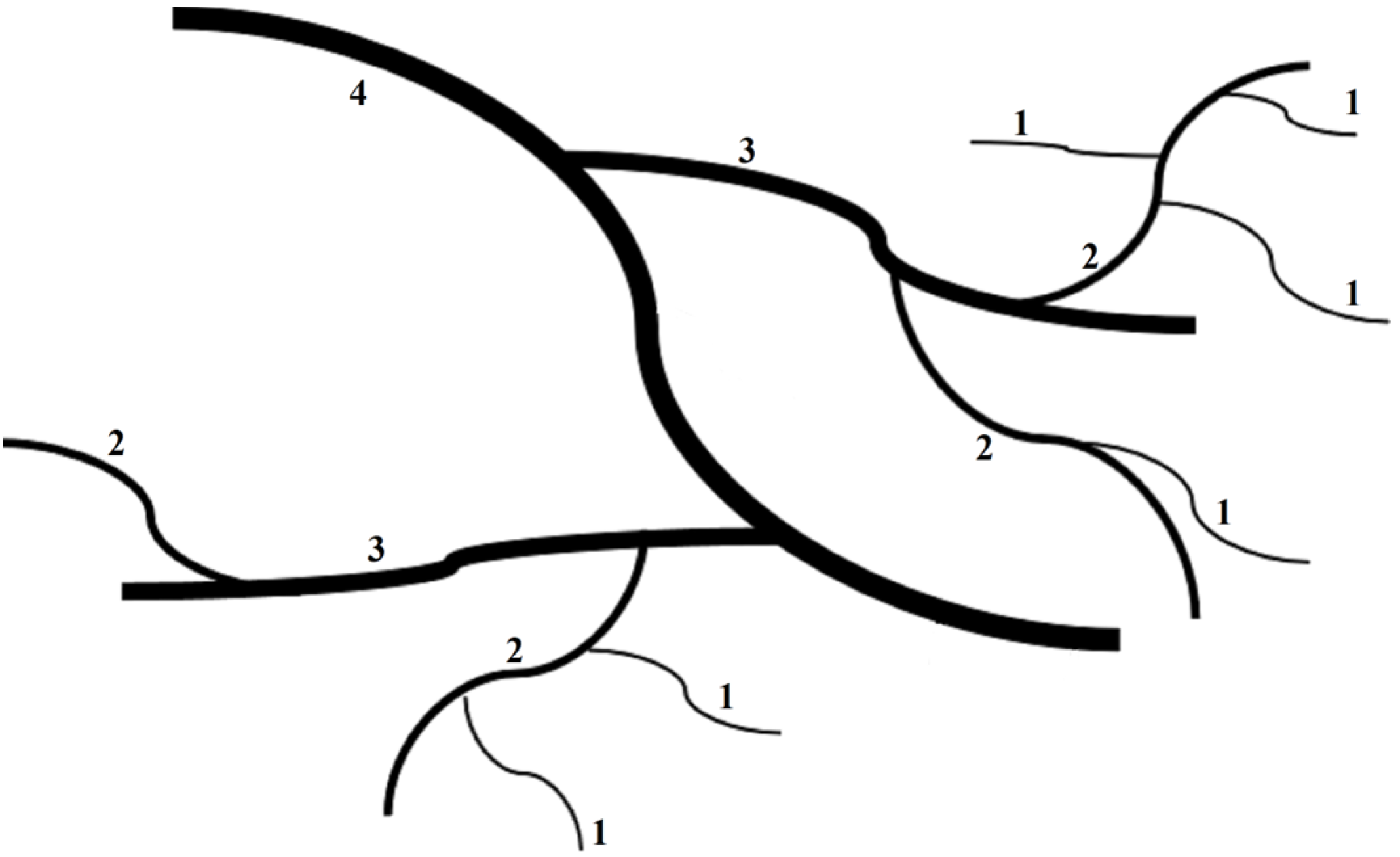
2.3. Field Survey and Sampling
2.4. Determination of the Ecological Stoichiometry Characteristics of Fine Roots
2.5. Determination of Soil Environmental Factors and Ecological Stoichiometry Characteristics
2.6. Mathematical Statistical Methods
3. Results
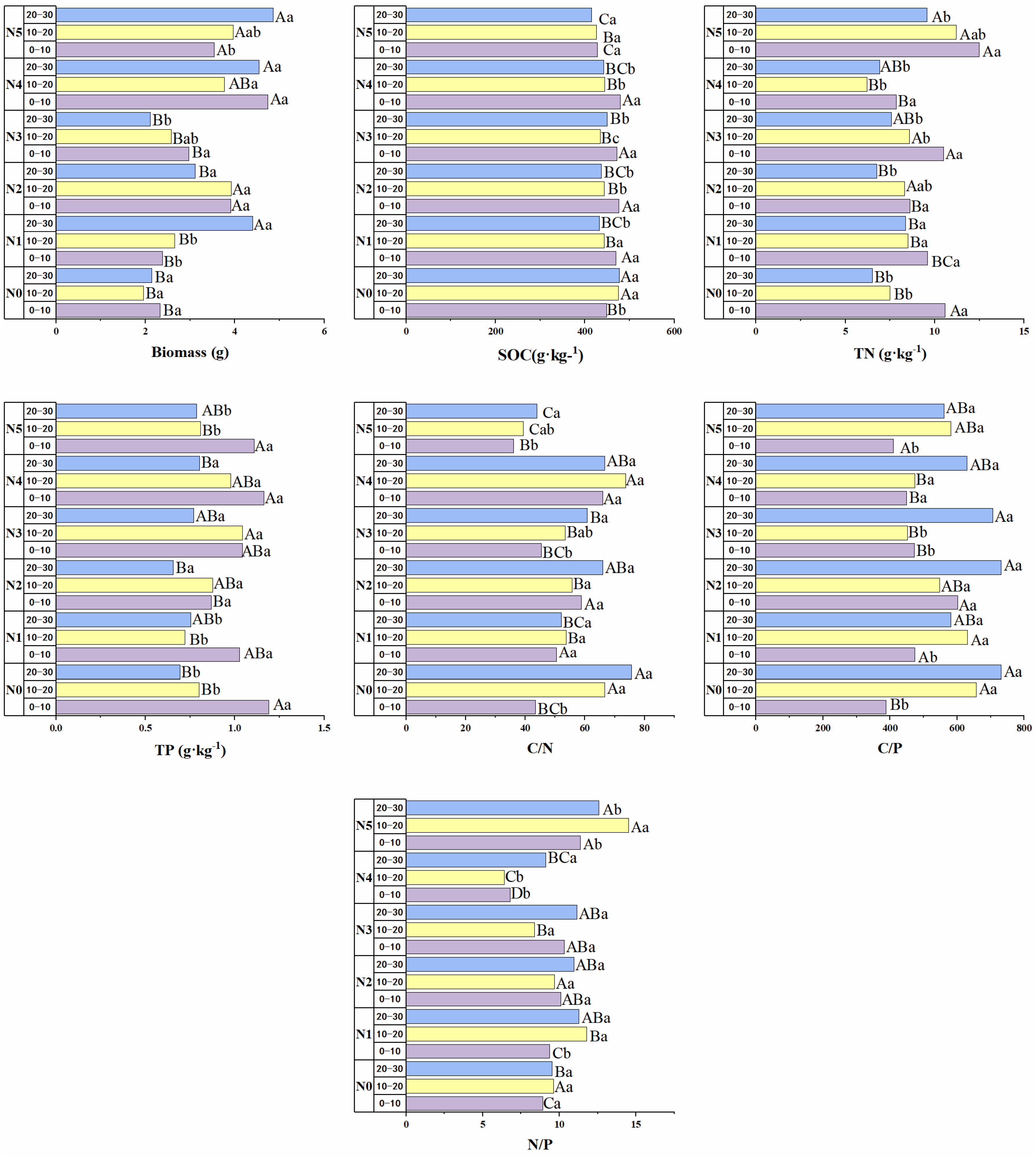
3.1. Fine-Root Biomass and Ecological Stoichiometry
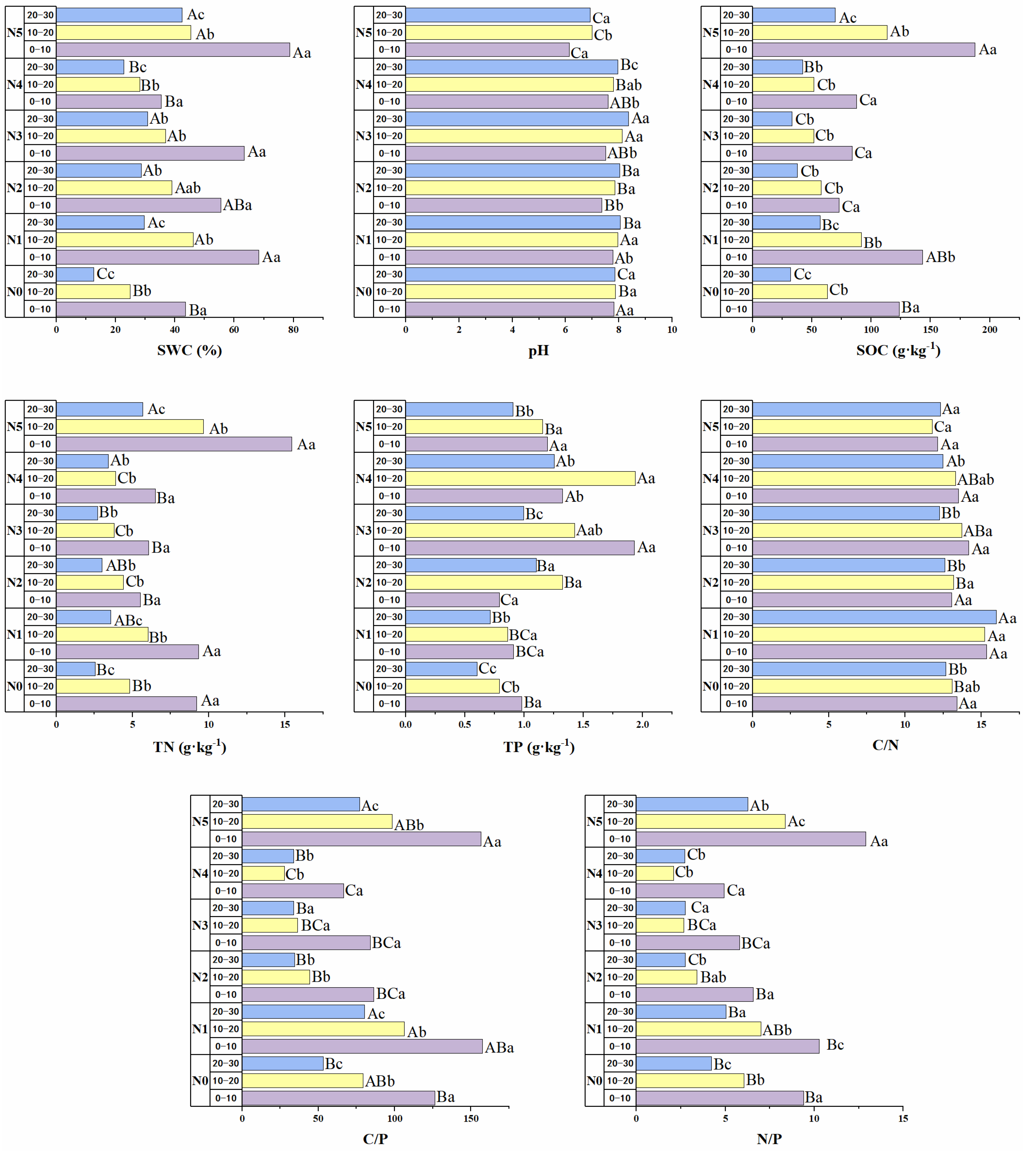
3.2. Soil Environmental Factors and Ecological Stoichiometry
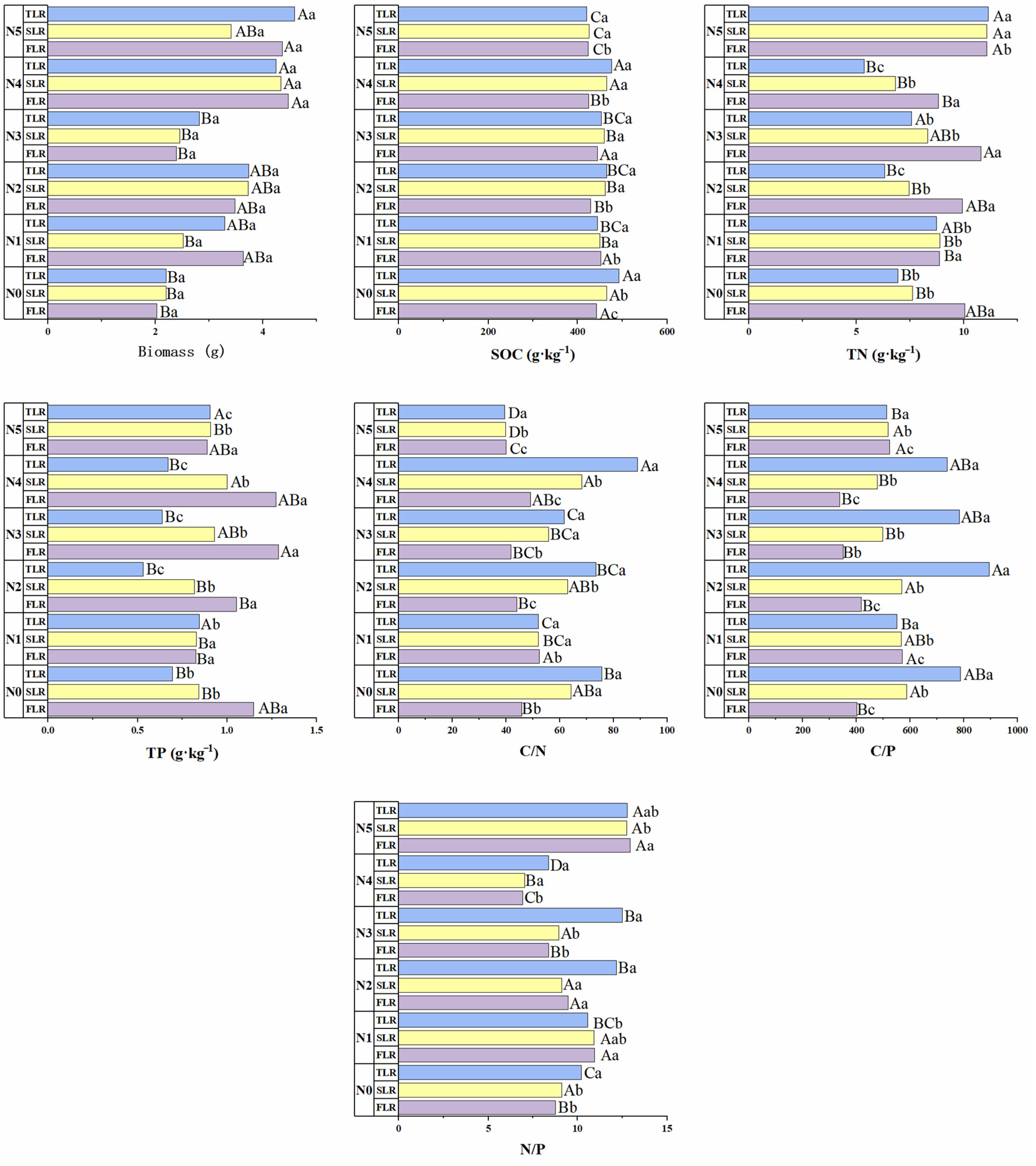
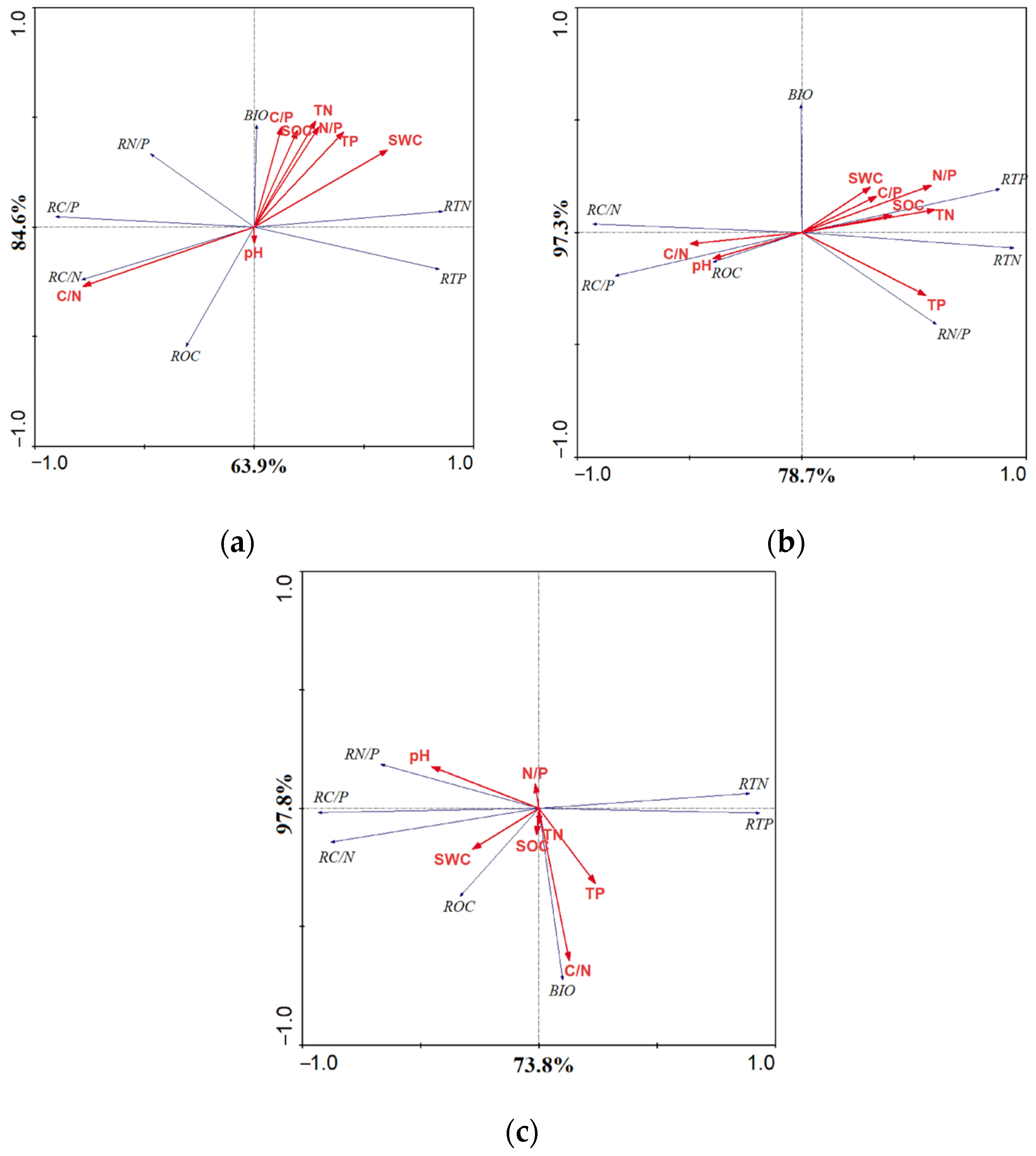
3.3. Fine-Root Soil Ecological Stoichiometry
4. Discussion
4.1. Ecological Stoichiometry Effects of Different Proportions of Nitrogen Inputs on the Fine Roots of Picea Schrenkiana
4.2. Effect of Different Proportions of Nitrogen Source Inputs on the Soil Ecological Stoichiometry of Picea Schrenkiana
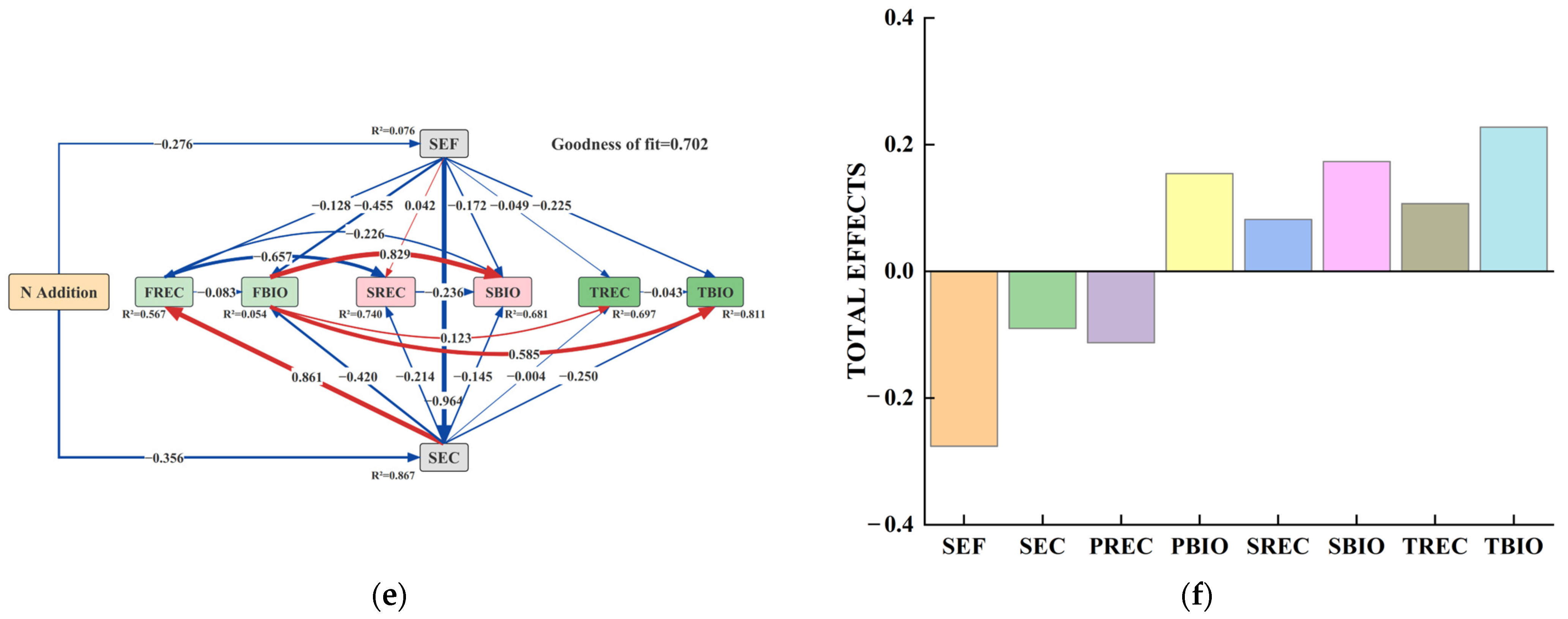
4.3. Ecological Stoichiometry Correlation and Influencing Factors of Fine-Root Soil of Picea Schrenkiana with Different Proportions of Nitrogen Inputs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamarque, J.F.; Kiehl, J.T.; Brasseur, G.P.; Butler, T.; Cameron-Smith, P.; Collins, W.D.; Collins, W.J.; Granier, C.; Hauglustaine, D.; Hess, P.G.; et al. Assessing future nitrogen deposition and carbon cycle feedback using a multimodel approach: Analysis of nitrogen deposition. J. Geophysres-Atmos. 2005, 110, D19. [Google Scholar] [CrossRef] [Green Version]
- He, J.Z.; Zhang, L.M. Key processes and microbial mechanisms of soil nitrogen transformation. Microbiol. China 2013, 40, 98–108. [Google Scholar]
- Valverde-Barrantes, O.J.; Freschet, G.T.; Roumet, C.; Blackwood, C.B. A worldview of root traits: The influence of ancestry, growth form, climate and mycorrhizal association on the functional trait variation of fine-root tissues in seed plants. New Phytol. 2017, 215, 1562–1573. [Google Scholar] [CrossRef] [Green Version]
- Heljä-Sisko, H.; Kirsi, M.; Seppo, K.; Esko, V.; Eino, M. Below- and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. Forest Ecol. Manag. 2002, 165, 317–326. [Google Scholar]
- Fan, A.L.; Zhang, L.H.; Chen, T.T.; Chen, Y.H.; Jiang, Q.; Jia, L.Q.; Wang, X.; Chen, G.S. Response of fine root stoichiometric traits to nitrogen addition in ectomycorrhizal and arbuscular mycorrhizal tree species in an evergreen broad-leaved forest. Acta Ecol. Sin. 2020, 40, 4966–4974. [Google Scholar]
- Zhu, F.F.; Yoh, M.; Gilliam, F.S.; Lu, X.K.; Mo, J.M. Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition: Insights from fine root responses to nutrient additions. PLoS ONE 2013, 8, e82661. [Google Scholar] [CrossRef] [Green Version]
- Mo, Q.F.; Zou, B.; Li, Y.W.; Chen, Y.; Zhang, W.X.; Mao, R.; Ding, Y.Z.; Wang, J.; Lu, X.K.; Li, X.B.; et al. Response of plant nutrient stoichiometry to fertilization varied with plant tissues in a tropical forest. Sci. Rep. 2015, 5, 14605. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.Z.; Xiong, D.C.; Feng, J.X.; Xu, C.S.; Zhong, B.Y.; Deng, F.; Chen, Y.Y.; Chen, G.S.; Yang, Y.S. Ecophysiological effects of simulated nitrogen deposition on fine roots of Chinese fir (Cunninghamia lanceolata) seedlings. Acta Ecol. Sin. 2017, 37, 74–83. [Google Scholar]
- Song, L.; Kuang, F.H.; Skiba, U.; Zhu, B.; Liu, X.J.; Levy, P.; Dore, A.; Fowler, D. Bulk deposition of organic and inorganic nitrogen in southwest China from 2008 to 2013. Environ. Pollut. 2017, 227, 157–166. [Google Scholar] [CrossRef]
- Li, Y.N. Effects of Nitrogen forms and Nitrogen Gradient on Nutritional Growth of Cymbidium Seedling. Master’s Thesis, Beijing Forestry University, Beijing, China, 2010. [Google Scholar]
- Wei, L.L.; Chen, C.R.; Yu, S. Uptake of organic nitrogen and preference for inorganic nitrogen by two Australian native Araucariaceae species. Plant Ecol. Divers. 2015, 8, 259–264. [Google Scholar] [CrossRef]
- Silva, R.C.; Rondina, A.B.; Zangaro, W.; Oliveira, H.C. Inorganic nitrogen sources alter the root morphology of neotropical tree seedlings from different successional groups. Trees 2021, 35, 875–887. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wang, Y.L.; Jin, B.W. Studies on regulating function of forest hydrology and microclimate in arid and semi-arid area of west. Sci. Silvae Sin. 2001, 37, 120–125. [Google Scholar]
- Sun, X.J.; Chang, S.L.; Song, C.C.; Zhang, Y.T. Age-related N, P, and K stoichiometry in different organs of Picea schrenkiana. Chin. J. Ecol. 2018, 37, 1291–1298. [Google Scholar]
- Wang, X.L.; Li, K.H.; Song, W.; Adeli, M.; Liu, X.J. Atmospheric Nitrogen Dry Deposition in Forest Ecosystem in the Central Tianshan Mountains, Xinjiang. Arid Zone Res. 2016, 33, 830–836. [Google Scholar]
- Pregitzer, K.S. Fine roots of trees—A new perspective. New Phytol. 2002, 154, 267–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, S.D. Agrochemical Analysis of Soil, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 268–270. [Google Scholar]
- Chen, M.; Cheng, H.Z.; Yao, X.D.; Cao, L.R.; Chen, R.; Chen, G.S.; Wang, X.H. Effects of soil warming and nitrogen addition on the morphological and chemical characteristics of fine roots in different order classes if the Chinese fir. Acta Ecol. Sin. 2023; accepted. [Google Scholar]
- Högberg, P.; Nordgren, A.; Ågren, G.I. Carbon allocation between tree root growth and root respiration in boreal pine forest. Oecologia 2002, 132, 579–581. [Google Scholar] [CrossRef]
- Xiong, D.C.; Huang, J.X.; Yang, Z.J.; Lu, Z.L.; Chen, G.S.; Yang, Y.S.; Xie, J.S. Nutrient heterogeneity in fine roots of six subtropical natural tree species. Acta Ecol. Sin. 2012, 32, 4343–4351. [Google Scholar] [CrossRef] [Green Version]
- Ostertag, R. Effects of nitrogen and phosphorus availability on fine-root dynamics in Hawaiian montane forests. Ecology 2001, 82, 485–499. [Google Scholar] [CrossRef]
- Li, H.X.; Xu, C.L.; Guo, S.P.; Guo, C.B.; Bai, H. Effects of different nitrogen supply on biomass, N and P concentration and their seasonal variation of Larix olgensis seedlings. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2014, 38, 79–84. [Google Scholar]
- An, W.L.; Zeng, C.S.; Wang, W.Q. Simulation of effect of acid rain on greenhouse gases emissions in paddy field of Fuzhou plain. Acta Sci. Circumstantiae 2017, 37, 3984–3994. [Google Scholar]
- Guo, R.Q.; Xiong, D.C.; Song, T.T.; Cai, Y.Y.; Chen, T.T.; Chen, W.Y.; Zheng, X.; Chen, G.S. Effects of simulated nitrogen deposition on stoichiometry of fine roots of Chinese fir (Cunninghamia lanceolata) seedlings. Acta Ecologica Sinica 2018, 38, 6101–6110. [Google Scholar]
- Zhao, J.J. The Effects of Fine Root Physiological and Ecological Traits to Added Nitrogen in Schrenk’s Spruce (Picea schrenkiana) of the Tianshan Mountains, China. Master’s Thesis, Xinjiang University, Urumqi, China, 2019. [Google Scholar]
- Yu, J.S.; Song, Z.P.; Hou, J.H. Effects of nitrogen addition on N-P allocation strategies in different organs of Pinus tabulaeformis in its natural forest. Acta Ecol. Sin. 2022, 42, 732–741. [Google Scholar]
- Li, W.B.; Jin, C.J.; Guan, D.X.; Wang, Q.K.; Wang, A.Z.; Yuan, F.H.; Wu, J.B. The effects of simulated nitrogen deposition on plant root traits: A meta-analysis. Soil Boil. Biochem. 2015, 82, 112–118. [Google Scholar] [CrossRef]
- He, J.H.; Wang, F.C.; Zhang, L.S.; Chen, F.S.; Xia, C.K.; Zhou, Y.F.; Quan, J.W.; Shen, L.L.; Liu, Q. Effects of nitrogen and phosphorus addition on plant-litter-soil stoichiometric characteristics in a subtropical fir plantation. Acta Agric. Univ. Jiangxiensis 2022, 13, 834184. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, e103. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.J. Effect of Nitrogen Addition on Nitrogen Acquisition Strategies of Three Tree Species of Korean Pine Broadleaved Forests in Xiaoxing’ an Mountains. Master’s Thesis, Northeast Forestry University, Harbin, China, 2020. [Google Scholar]
- Zhang, J.B.; Cai, Z.C.; Zhu, T.B.; Yang, W.Y.; Müller, C. Mechanisms for the retention of inorganic N in acidic forest soils of southern China. Sci. Rep. 2013, 3, 2342. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, P.; Emmett, B.A.; Kjønaas, O.J.; Koopmans, C.J.; Tietema, A. Impact of nitrogen deposition on nitrogen cycling in forests: A synthesis of NITREX data. Forest Ecol. Manag. 1998, 101, 37–55. [Google Scholar] [CrossRef]
- Tian, P.; Liu, S.E.; Wang, Q.k.; Sun, T.; Blagodatskaya, E. Organic N deposition favours soil C sequestration by decreasing priming effect. Plant Soil. 2019, 445, 439–451. [Google Scholar] [CrossRef]
- Ye, Y.H.; Gao, Y.; Liu, Y.L.; Han, Y.Y.; Ye, H. Short-term effects of nitrogen deposition on soil physical and chemical properties of Tibetan forest-grassland landscape boundary. Anim. Husb. Feed. Sci. 2018, 10, 249–256. [Google Scholar]
- Li, M. Effects of Nitrogen Addition on Carbon, Nitrogen, Phosphorus and Microbial Characteristics of Soil Aggregates in Stipa Baicalensis Grassland. Master’s Thesis, Inner Mongolia Normal University, Hohhot, China, 2020. [Google Scholar]
- Du, Y.G.; Cao, Y.F.; Zhou, G.; Guo, X.W. Effect of Simulated Nitrogen Deposition and Adding Precipitation and Its Interaction on Soil N2O Emissions in Alpine Meadow. Chin. J. Grassl. 2017, 39, 116–120. [Google Scholar]
- Bonnie, L.K.; Sarah, E.H.; Laurie, E.K. Effects of Long-Term Nitrogen Addition on Microbial Enzyme Activity in Eight Forested and Grassland Sites: Implications for Litter and Soil Organic Matter Decomposition. Ecosystems 2009, 12, 1–15. [Google Scholar]
- Tu, L.H.; Hu, T.X.; Zhang, J.; Li, R.H.; Dai, H.Z.; Luo, S.H. Response of soil organic carbon and nutrients to simulated nitrogen deposition in Pleioblastus amarus plantation, Rainy Area of West China. Chin. J. Plant Ecol. 2011, 35, 125–136. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J.; Gu, J.C.; Yu, L.Z.; Wang, Z.Q. Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation. Ann. Forest. Sci. 2015, 72, 435–442. [Google Scholar] [CrossRef]
- Tu, L.H.; Hu, H.L.; Hu, T.X.; Zhang, J.; Xiao, Y.L.; Luo, S.H.; Li, R.H.; Dai, H.Z. Effects of simulated nitrogen deposition on soil enzyme activities in a Betula luminifera plantation in Rainy Area of West China. Chin. J. Appl. Ecol. 2012, 23, 2129–2134. [Google Scholar]
- Zhang, J.Y.; Li, Y.S.; Yu, Z.H.; Xie, Z.H.; Liu, J.J.; Wang, G.H.; Liu, X.B.; Wu, J.J.; Herbert, S.J.; Jin, J. Nitrogen Cycling in the Crop-Soil Continuum in Response to Elevated Atmospheric CO2 Concentration and Temperature—A Review. Sci. Agric. Sin. 2021, 54, 1684–1701. [Google Scholar]
- Zhou, H.; Shi, H.B.; Zhang, W.C.; Wang, W.G.; Su, Y.D.; Yan, Y. Evaluation of Organic-Inorganic Nitrogen Application on Maize Yield and Nitrogen Leaching by DNDC Model. Trans. Chin. Soc. Agric. Mach. 2021, 52, 291–301, 249. [Google Scholar]
- Yu, H.; Pan, Z.T.; Chen, Z.Q.; Chen, Z.B.; Feng, L.J. Effects of nitrogen addition on soil chemical properties and leaf functional traits of Dicranopteris dichotoma in the red soil erosion area of southern China. Chin. J. Appl. Environ. Biol. 2021, 27, 46–53. [Google Scholar]
- Wang, L.L. Research on Ecological Stoichiometry of Carbon, Nitrogen and Phosphorus and Soil Fertility Change in the Larix pfincipis-rupprechti Plantation. Master’s Thesis, Northwest A&F University, Xianyang, China, 2016. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gradient | Organic N/kg·hm−1·a−1 | Inorganic N/kg·hm−2·a−1 |
|---|---|---|
| N0 | 0 | 0 |
| N1 | 10 | 0 |
| N2 | 7 | 3 |
| N3 | 5 | 5 |
| N4 | 3 | 7 |
| N5 | 0 | 10 |
| Cumulative Percentage Variance | Axis I | Axis II | Axis III | Axis IV |
|---|---|---|---|---|
| Organic nitrogen relation | 63.90 | 84.60 | 99.30 | 100.00 |
| Inorganic nitrogen relation | 78.70 | 97.30 | 99.80 | 100.00 |
| Mixed organic–inorganic nitrogen relation | 73.80 | 97.80 | 99.10 | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Gong, L.; Ding, Z.; Wu, X. Fine-Root Soil Stoichiometry of Picea schrenkiana Fisch. et Mey. and Its Correlation with Soil Environmental Factors under Different Nitrogen Input Levels in the Tianshan Mountains, Xinjiang. Forests 2023, 14, 426. https://doi.org/10.3390/f14020426
Zhang H, Gong L, Ding Z, Wu X. Fine-Root Soil Stoichiometry of Picea schrenkiana Fisch. et Mey. and Its Correlation with Soil Environmental Factors under Different Nitrogen Input Levels in the Tianshan Mountains, Xinjiang. Forests. 2023; 14(2):426. https://doi.org/10.3390/f14020426
Chicago/Turabian StyleZhang, Han, Lu Gong, Zhaolong Ding, and Xue Wu. 2023. "Fine-Root Soil Stoichiometry of Picea schrenkiana Fisch. et Mey. and Its Correlation with Soil Environmental Factors under Different Nitrogen Input Levels in the Tianshan Mountains, Xinjiang" Forests 14, no. 2: 426. https://doi.org/10.3390/f14020426