
The Synergistic Effects of AMF Inoculation and Boron Deficiency on the Growth and Physiology of Camellia oleifera Seedlings
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Treatments
2.2. Sampling and Plant Growth Parameters
2.3. Boron Analysis and Physiological Parameter Evaluation
2.4. Statistical Analysis
3. Results
3.1. The Effect of Boron Deficiency and AMF Inoculation on the Growth Parameters
3.2. Boron-Deficiency Effect on AMF Root Colonization Rate, Activity, and Morphology
3.3. Effect of Boron Deficiency and AMF Inoculation on Boron Content and Accumulation
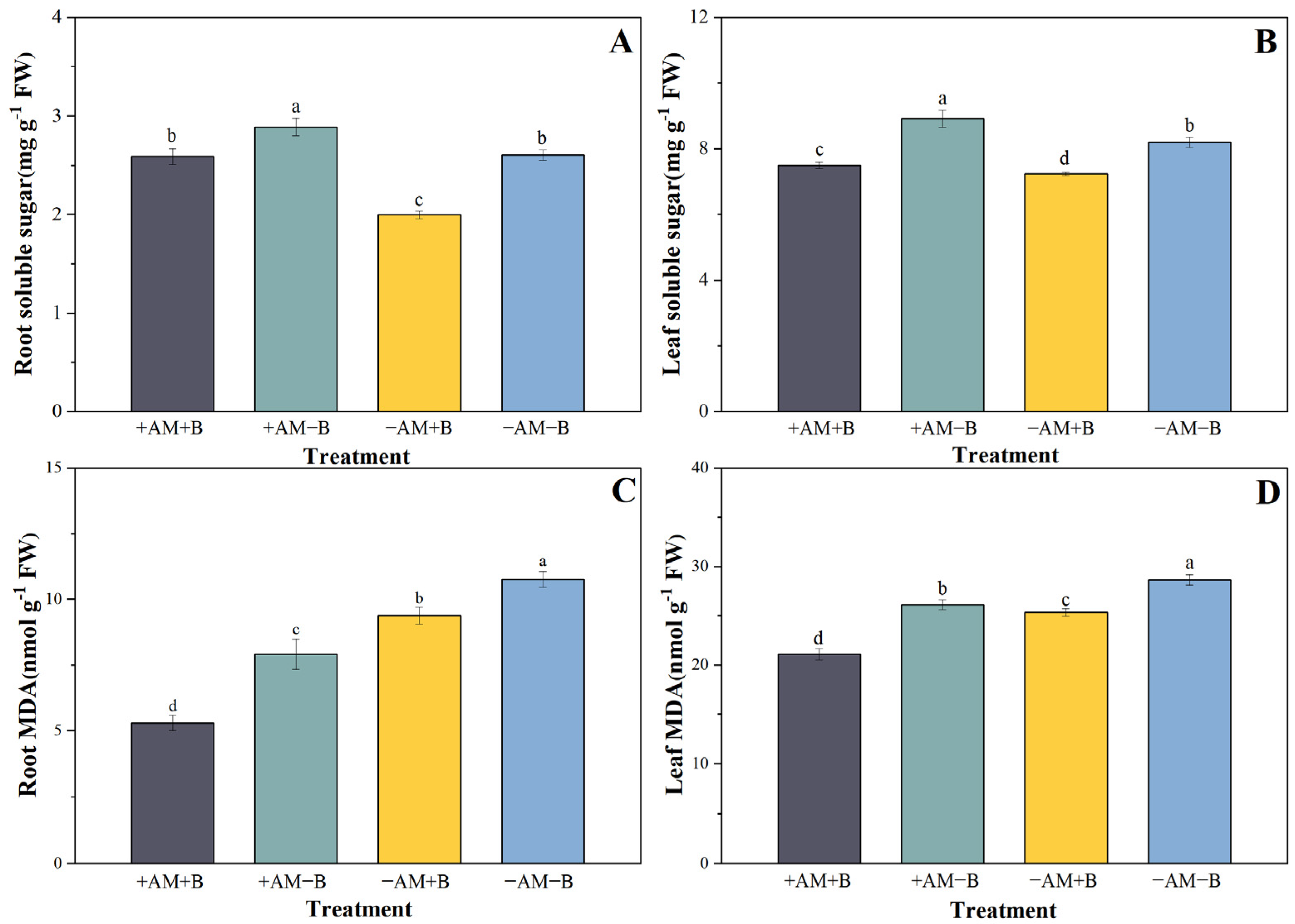
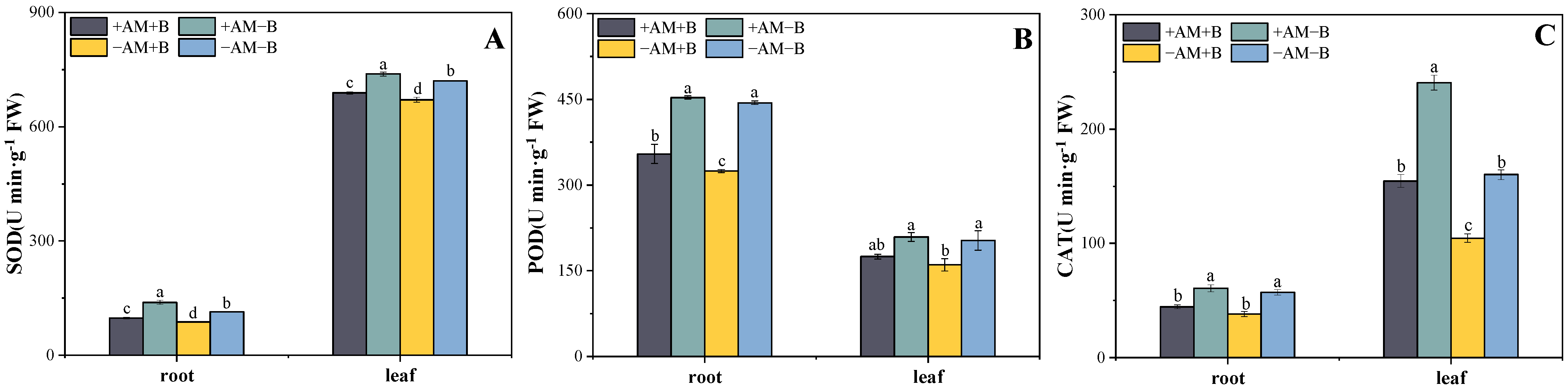
3.4. Effect of Boron Deficiency and AMF Inoculation on Antioxidant Enzyme Activity
3.5. Principal Component Analysis
4. Discussion
4.1. Effect of Boron Deficiency on AMF Colonization
4.2. Effect of AMF on Plant Growth, Boron Absorption, and Utilization
4.3. Effect of AMF and Boron Deficiency on Plant Physiological Characteristics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yan, L.; Li, S.; Muhammad, R.; Jiang, C.C. Proline metabolism and biosynthesis behave differently in response to boron-deficiency and toxicity in Brassica napus. Plant Physiol. Biochem. 2021, 167, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Mehboob, N.; Yasirasir, T.A.; Ul-Allah, S.; Nawaz, A.; Ahmad, N.; Hussain, M. Interactive effect of boron application methods and boron-tolerant bacteria (Bacillus sp. MN54) improves nodulation, grain yield, profitability and biofortification of kabuli chickpea grown under irrigated and rainfed conditions. J. Soil Sci. Plant Nutr. 2022, 22, 5068–5085. [Google Scholar] [CrossRef]
- Wu, X.W.; Riaz, M.; Yan, L.; Jiang, C.C. Distribution and mobility of foliar-applied boron (10B) in citrange rootstock under different boron conditions. Plant Growth Regul. 2020, 39, 575–582. [Google Scholar] [CrossRef]
- Dell, B.; Huang, L.B. Physiological response of plants to low boron. Plant Soil 1997, 193, 103–120. [Google Scholar] [CrossRef]
- Subedi, K.D.; Gregory, P.J.; Gooding, M.J. Boron accumulation and partitioning in wheat cultivars with contrasting tolerance to boron deficiency. Plant Soil 1999, 214, 141–152. [Google Scholar] [CrossRef]
- Atique-ur-Rehman; Farooq, M.; Nawaz, A.; Rehman, A.; Iqbal, S. Soil application of boron improves the tillering, leaf elongation, panicle fertility, yield and its grain enrichment in fine-grain aromatic rice. J. Plant Nutr. 2015, 38, 338–354. [Google Scholar] [CrossRef]
- García-Sánchez, F.; Simón-Grao, S.; Martínez-Nicolás, J.J.; Alfosea-Simón, M.; Liu, C.G.; Chatzissavvidis, C.; Pérez-Pérez, J.G.; Cámara-Zapata, J.M. Multiple stresses occurring with boron toxicity and deficiency in plants. J. Hazard. Mater. 2020, 397, 122713. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees, a review. Mycorrhiza 2011, 21, 71–90. [Google Scholar] [CrossRef]
- Liu, C.G.; Dai, Z.; Cui, M.Y.; Lu, W.K.; Sun, H.W. Arbuscular mycorrhizal fungi alleviate boron toxicity in Puccinellia tenuiflora under the combined stresses of salt and drought. Environ. Pollut. 2018, 240, 557–565. [Google Scholar] [CrossRef]
- Simón-Grao, S.; Nieves, M.; Martínez-Nicolás, J.J.; Alfosea-Simón, M.; Cámara-Zapata, J.M.; Fernández-Zapata, J.C.; García-Sánchez, F. Arbuscular mycorrhizal symbiosis improves tolerance of Carrizo citrange to excess boron supply by reducing leaf B concentration and toxicity in the leaves and roots. Ecotox. Environ. Saf. 2019, 173, 322–330. [Google Scholar] [CrossRef]
- Lavola, A.; Aphalo, P.J.; Lehto, T. Boron and other elements in sporophores of ectomycorrhizal and saprotrophic fungi. Mycorrhiza 2001, 21, 155–165. [Google Scholar] [CrossRef]
- Nehls, U. Mastering ectomycorrhizal symbiosis: The impact of carbohydrates. J. Exp. Bot. 2008, 59, 1097–1108. [Google Scholar] [CrossRef]
- Möttönen, M.; Lehto, T.; Aphalo, P.J. Growth dynamics and mycorrhizas of Norway spruce (Picea abies) seedings in relation to boron supply. Trees 2001, 15, 319–326. [Google Scholar] [CrossRef]
- Räisänen, M.; Repo, T.; Lehto, T. Cold acclimation was partially impaired in boron deficient Norway spruce seedings. Plant Soil 2007, 292, 271–282. [Google Scholar] [CrossRef]
- Bolaños, L.; Redondo-Nieto, M.; Bonilla, I.; Wall, L.G. Boron requirement in the Discaria trinervis (Rhamnaceae) and Frankia symbiotic relationship. Its essentiality for Frankia BCU110501 growth and nitrogen fixation. Physiol. Plant. 2002, 115, 563–570. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Mao, P.; Yu, D.S.; Xu, L. Typical soil pH change characteristics in red soil area in the past 25 years—Taking Yujiang County of Jiangxi Province as an example. Act. Pedol. Sin. 2018, 55, 1545–1553. (In Chinese) [Google Scholar]
- Lin, Y.L.; Li, Z.Y.; Zhang, L.P.; Wu, F.; Yang, Y.; Tan, M.X.; Hu, D.N. Effects of organic phosphorus and AM fungi on growth, root morphology and photosynthetic characteristics of Camellia oleifera. Non-Wood For. Res. 2021, 39, 121–128+210. (In Chinese) [Google Scholar]
- Wang, J.E.; Li, D.W.; Zhang, Y.L.; Zhao, Q.; He, Y.M.; Gong, Z.H. Defence responses of pepper (Capsicum annuum L.) infected with incompatible and compatible strains of Phytophthora capsici. Eur. J. Plant Pathol. 2013, 136, 625–638. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piche, Y. Ink and vinegar, a simple staining technique for arbuscular mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed]
- Bingham, F.T. Boron. In Methods of Soil Analysis Part 2: Chemical and Microbiological Properties; American Society of Agronomy: Madison, WI, USA, 1982; pp. 431–446. [Google Scholar]
- Carmak, I.; Horst, J.H. Effects of aluminum on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, H.; Liu, J.N.; Huan, X.J.; Wang, Q.C.; Li, L.; Liu, Y.J.; Qin, P. Effects of drought and re-watering treatment on physiological and biochemical indexes of different drought-resistant wheat varieties/lines at seedling stage. Acta Agric. Boreali-Occident. Sin. 2020, 29, 1795–1802. (In Chinese) [Google Scholar]
- Zhou, W.; Leul, M. Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul. 1999, 27, 99–104. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Marschner, H. Phosphatase activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus. Soil Biol. Biochem. 1994, 26, 387–395. [Google Scholar] [CrossRef]
- Tanaka, Y.; Yano, K. Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied. Plant Cell Environ. 2005, 28, 1247–1254. [Google Scholar] [CrossRef]
- Kazadi, T.A.; Lwalaba, J.L.W.; Ansey, B.K.; Muzulukwau, J.M.; Katabe, G.M.; Karul, M.I.; Baert, G.; Haesaert, G.; Mundende, R.P.M. Effect of phosphorus and arbuscular mycorrhizal fungi (AMF) inoculation on growth and productivity of maize (Zea mays L.) in a tropical ferralsol. Gesunde Pflanz. 2022, 74, 159–165. [Google Scholar] [CrossRef]
- Camacho-Cristóbal, J.J.; Martín-Rejano, E.M.; Herrera-Rodríguez, M.B.; Navarro-Gochicoa, M.T.; Rexach, J.; González-Fontes, A. Boron deficiency inhibits root cell elongation via an ethylene/auxin/ros-dependent pathway in Arabidopsis seedlings. J. Exp. Bot. 2015, 66, 3831–3840. [Google Scholar] [CrossRef] [PubMed]
- Zarea, M.J.; Karimi, N.; Goltapeh, E.M.; Ghalavand, A. Effect of cropping systems and arbuscular mycorrhizal fungi on soil microbial activity and root nodule nitrogenase. J. Saudi Soc. Agric. Sci. 2011, 10, 109–120. [Google Scholar] [CrossRef]
- Chen, L.S.; Han, S.; Qi, Y.P.; Yang, L.T. Boron stresses and tolerance in citrus. Afr. J. Biotechnol. 2012, 11, 5961–5969. [Google Scholar]
- Schellenbaum, L.; Berta, G.; Ravolanirina, F.; Tisserant, B.; Gianinazzi, S.; Fitter, A.H. Influence of endomycorrhizal infection on root morphology in a micropropagated woody plant species (Vitis vinifera L.). Ann. Bot. 1991, 68, 135–141. [Google Scholar] [CrossRef]
- Borkowska, B. Growth and photosynthetic activity of micropropagated strawberry plants inoculated with endomycorrhizal fungi (AMF) and growing under drought stress. Acta Physiol. Plant. 2002, 24, 365–370. [Google Scholar] [CrossRef]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Singh, H.P.; Chaum, S. Foliar application of glycinebetaine regulates soluble sugars and modulates physiological adaptations in sweet potato (Ipomoea batatas) under water deficit. Protoplasma 2020, 257, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Pandey, A.; Kumar, B.; Palni, L.M.S. Enhancement in growth and quality parameters of tea [Camellia sinensis (L.) O. Kuntze] through inoculation with arbuscular mycorrhizal fungi in an acid soil. Biol. Fertil. Soils 2010, 46, 427–433. [Google Scholar] [CrossRef]
- Huang, Z.; Zou, Z.; He, C.; He, Z.; Zhang, Z.; Li, J. Physiological and photosynthetic responses of melon (Cucumis melo L.) seedlings to three Glomus species under water deficit. Plant Soil 2011, 339, 391–399. [Google Scholar] [CrossRef]
- Prasad, T.K. Role of catalase in inducing chilling tolerance in pre-emergent maize seedlings. Plant Physiol. 1997, 114, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, N.; Grimm, B.; Wobus, U.; Weschke, W. Differential response of antioxidant compounds to salinity stress in salt tolerant and salt-sensitive seedlings of foxtail millet (Setaria italica). Physiol. Plant. 2000, 109, 435–442. [Google Scholar] [CrossRef]
- Singh, K.; Yadav, S.K.; Mishra, R.K. Differential effect of salinity on thermotolerance of SOD isoforms in seven varieties of Vigna mungo (L.) Hepper. Plant Physiol. Rep. 2019, 24, 279–288. [Google Scholar] [CrossRef]
- Mazza, G.; Charles, C.; Ouchet, M.; Ricard, J.; Raynaud, J. Isolation, purification and physico-chemical properties of turnip peroxidases. Biochim. Biophys. Acta 1968, 167, 89–98. [Google Scholar] [CrossRef]
- Akensous, F.Z.; Anli, M.; Boutasknit, A.; Ben-Laouane, R.; Ait-Rahou, Y.; Ahmed, H.B.; Nasri, N.; Hafidi, M.; Meddich, A. Boosting date palm (Phoenix dactylifera L.) growth under drought stress: Effects of innovative biostimulants. Gesunde Pflanz. 2022, 74, 961–982. [Google Scholar] [CrossRef]
- Wang, J.P.; Yuan, J.H.; Ren, Q.; Zhang, B.; Zhang, J.C.; Huang, R.Z.; Wang, G.G. Arbuscular mycorrhizal fungi enhanced salt tolerance of Gleditsia sinensis by modulating antioxidant activity, ion balance and P/N ratio. Plant Growth Regul. 2022, 97, 33–49. [Google Scholar] [CrossRef]
- Pandey, N.; Archana. Antioxidant responses and water status in Brassica seedlings subjected to boron stress. Acta Physiol. Plant. 2013, 35, 697–706. [Google Scholar] [CrossRef]
- Begum, N.; Akhtar, K.; Ahanger, M.A.; Iqbal, M.; Wang, P.P.; Mustafa, N.S.; Zhang, L.X. Arbuscular mycorrhizal fungi improve growth, essential oil, secondary metabolism, and yield of tobacco (Nicotiana tabacum L.) under drought stress conditions. Environ. Sci. Pollut. Res. 2021, 28, 45276–45295. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | +AM+B | +AM−B | −AM+B | −AM−B |
|---|---|---|---|---|
| Height elongation (cm) | 3.45 ± 0.21 a | 2.81 ± 0.29 a | 1.35 ± 0.08 b | 1.26 ± 0.18 b |
| Ground diameter thickening (mm) | 1.28 ± 0.14 a | 1.04 ± 0.18 a | 1.05 ± 0.11 a | 0.42 ± 0.04 b |
| Root biomass (g) | 5.15 ± 0.51 a | 4.85 ± 0.40 a | 4.57 ± 1.00 a | 3.40 ± 0.50 b |
| Shoot biomass (g) | 10.03 ± 0.36 a | 9.47 ± 0.16 a | 9.64 ± 0.16 a | 7.29 ± 0.46 b |
| Total biomass (g) | 16.02 ± 0.81 a | 13.47 ± 0.74 ab | 15.56 ± 0.82 a | 11.29 ± 0.97 b |
| R/S ratio | 0.69 ± 0.01 a | 0.69 ± 0.01 a | 0.67 ± 0.03 a | 0.43 ± 0.05 b |
| Treatment | Principal Component Score | Scores | Sort | |
|---|---|---|---|---|
| F1 | F2 | |||
| +AM+B | 3.58 | 0.54 | 2.52 | 1 |
| +AM−B | −0.96 | 2.50 | 0.09 | 2 |
| −AM+B | 0.99 | −2.30 | −0.01 | 3 |
| −AM−B | −3.61 | −0.74 | −2.59 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, M.; Fan, J.; Ding, W.; Chen, L.; Luo, J.; Liu, Y.; Mei, L. The Synergistic Effects of AMF Inoculation and Boron Deficiency on the Growth and Physiology of Camellia oleifera Seedlings. Forests 2023, 14, 1126. https://doi.org/10.3390/f14061126
Liu J, Zhang M, Fan J, Ding W, Chen L, Luo J, Liu Y, Mei L. The Synergistic Effects of AMF Inoculation and Boron Deficiency on the Growth and Physiology of Camellia oleifera Seedlings. Forests. 2023; 14(6):1126. https://doi.org/10.3390/f14061126
Chicago/Turabian StyleLiu, Junying, Mengxue Zhang, Jie Fan, Wenna Ding, Longsheng Chen, Jie Luo, Yongzhong Liu, and Li Mei. 2023. "The Synergistic Effects of AMF Inoculation and Boron Deficiency on the Growth and Physiology of Camellia oleifera Seedlings" Forests 14, no. 6: 1126. https://doi.org/10.3390/f14061126