
Effect of Sun Exposure of the Horse Chestnut (Aesculus hippocastanum L.) on the Occurrence and Number of Cameraria ohridella (Lepidoptera: Gracillariidae)
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Place of Research and Plant Material
2.2. Estimation of Orchid Miner Abundance Using Pheromone Traps
2.3. Sampling of Chestnut Leaves Damaged by C. ohridella
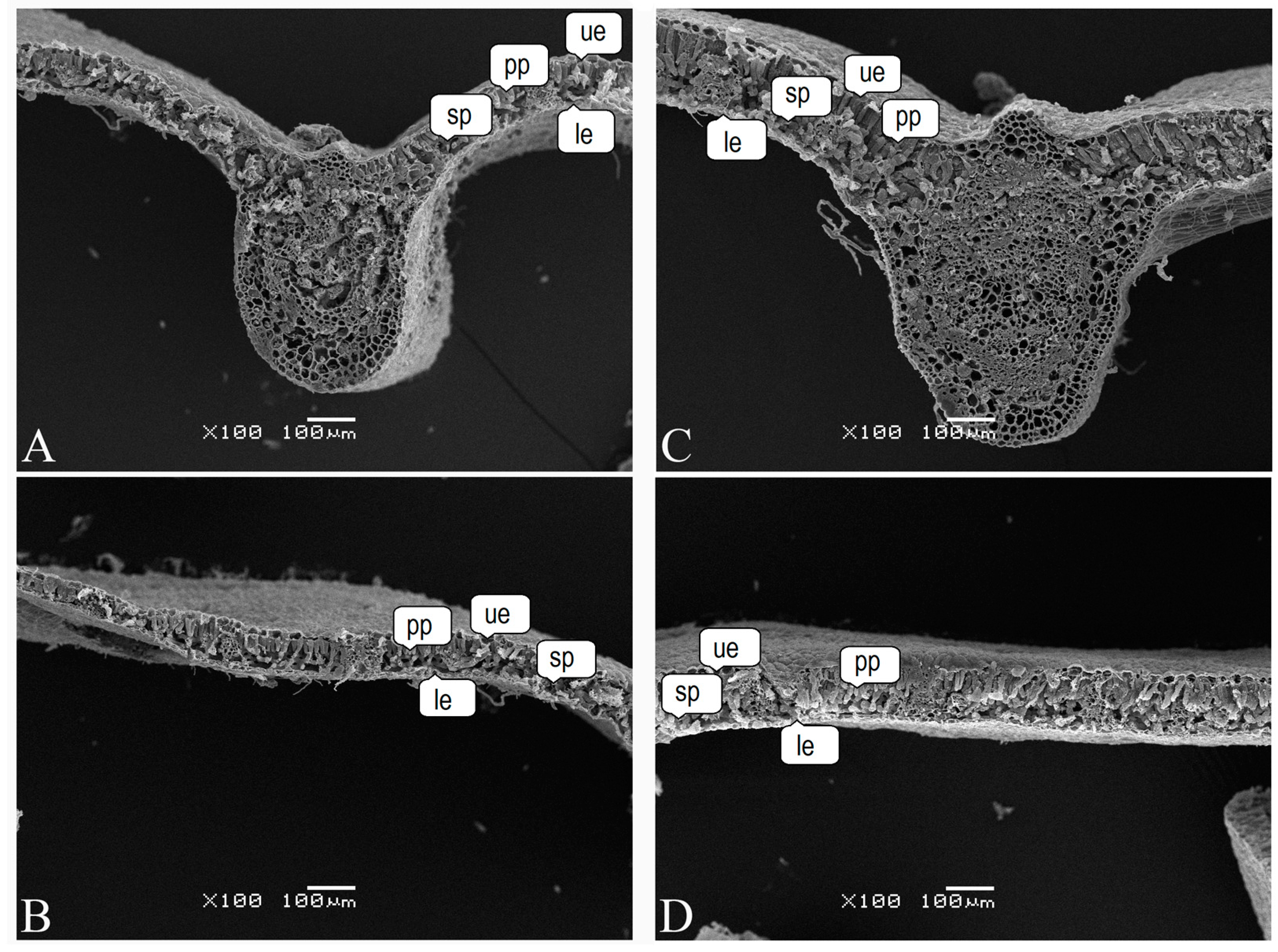
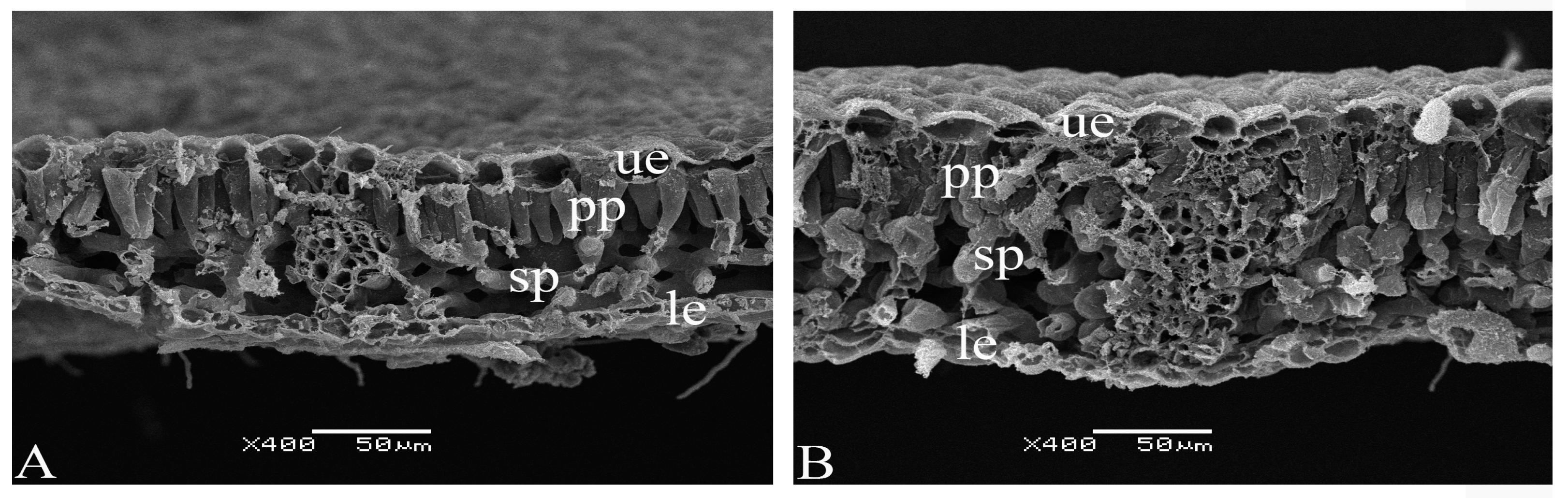
2.4. Scanning Electron Microscopy
2.5. Determination of the Composition of Phenolic Compounds in Chestnut Leaves
2.6. Statistical Analysis
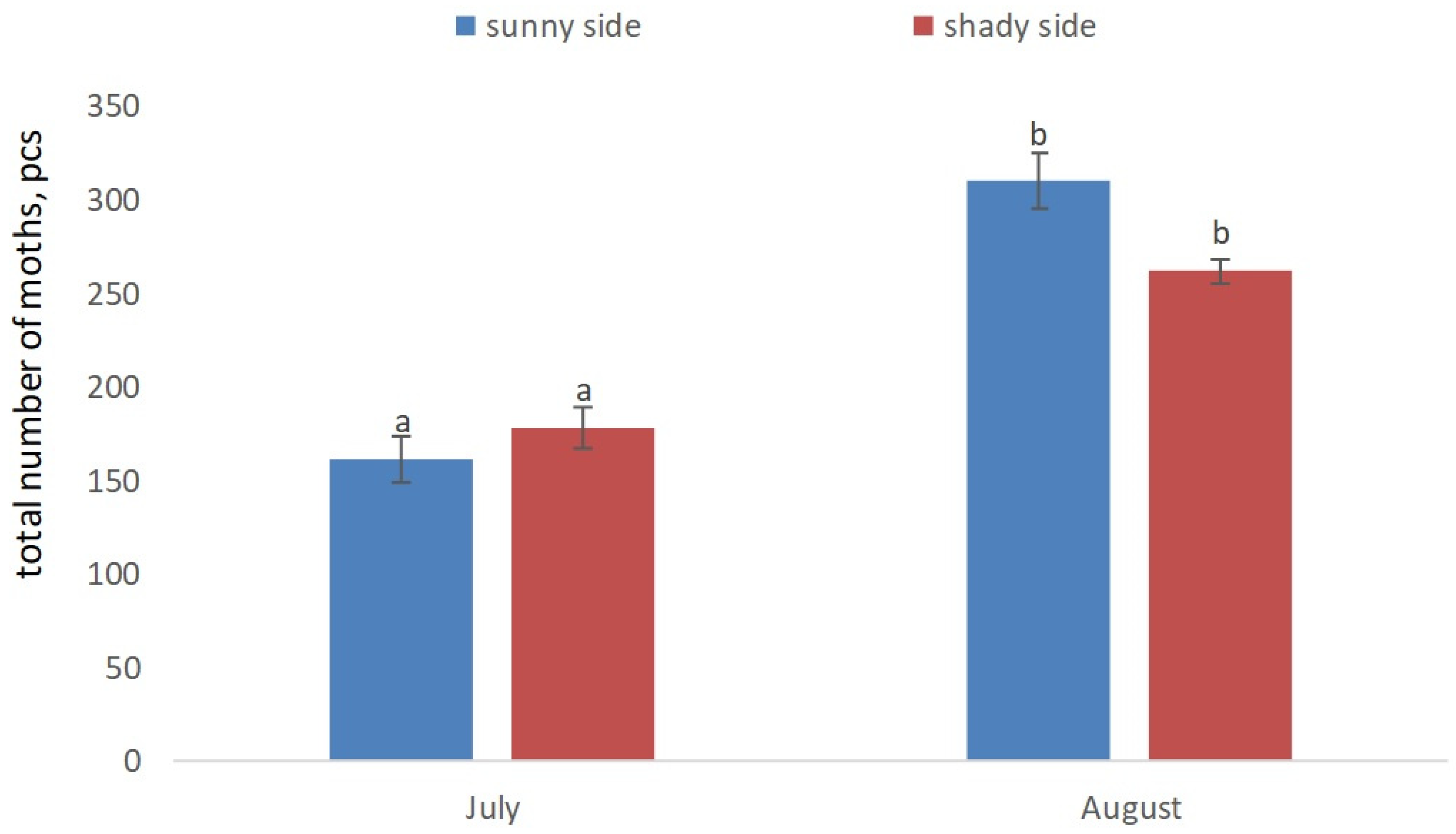
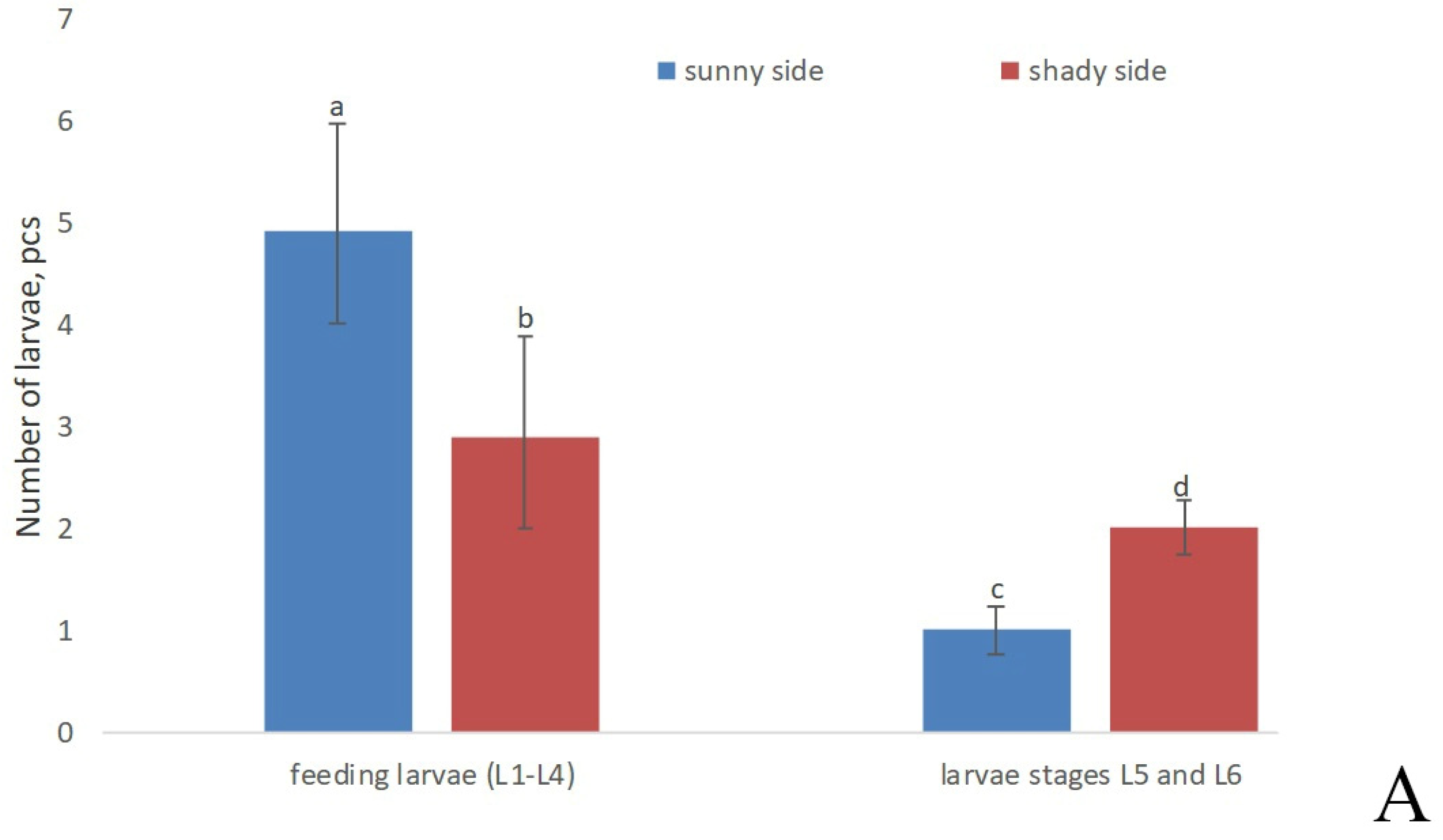
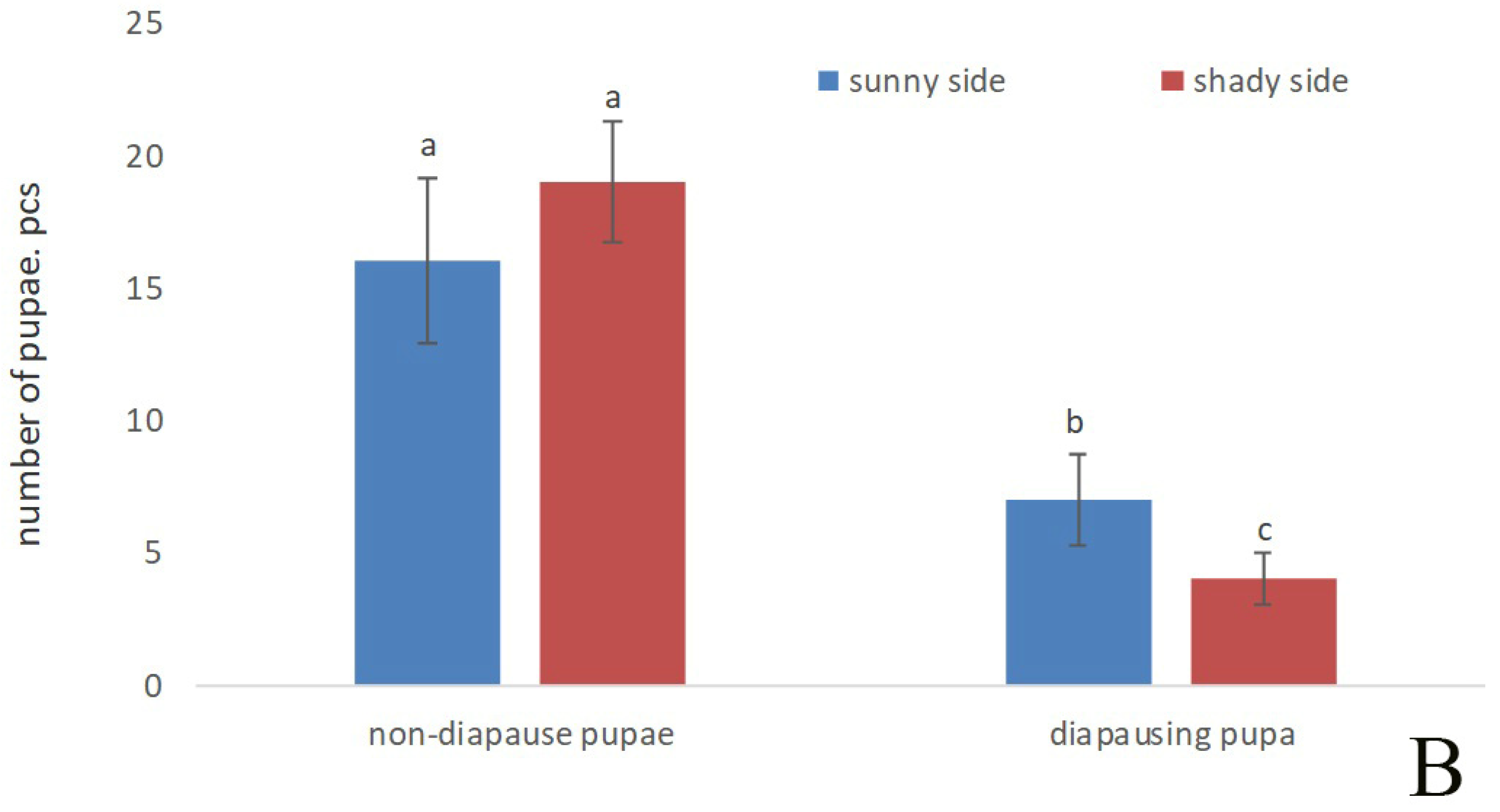
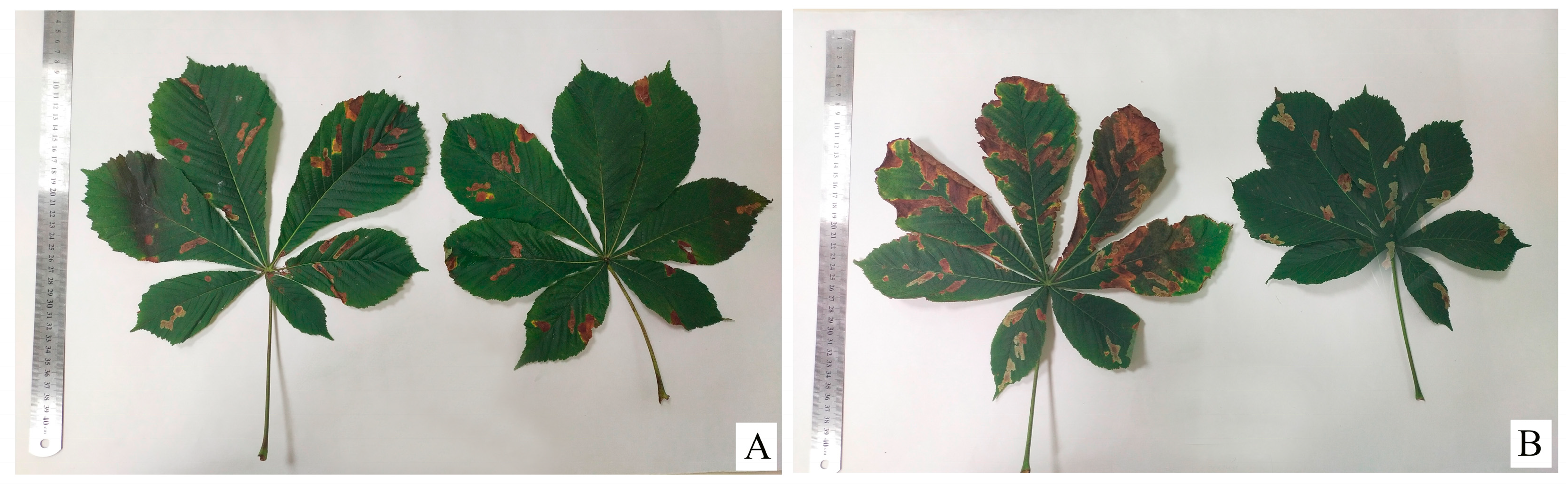
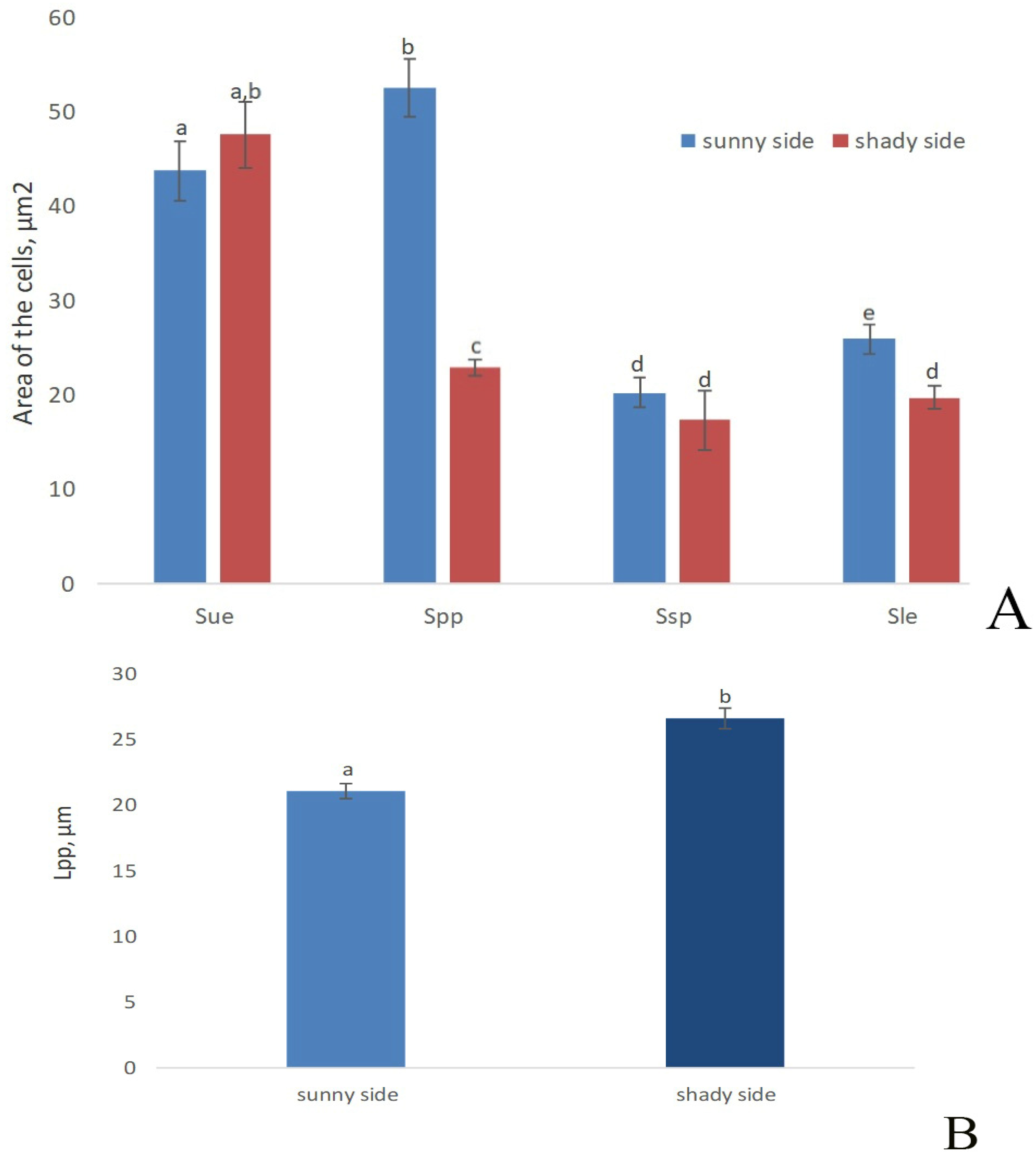
3. Results
Metabolic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takács, Á.; Kiss, M.; Gulyás, Á.; Tanács, E.; Kántor, N. Solar permeability of different tree species in Szeged, Hungary. Geogr. Pannonica 2016, 20, 32–41. [Google Scholar] [CrossRef]
- Chanon, A.M. Studies on the Reproductive Capacity of Aesculus parviflora and Aesculus pavia: Opportunities for Their Improvement through Interspecific Hybridization. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2005; p. 264. [Google Scholar]
- Freer-Smith, P.H.; Webber, J.F. Tree pests and diseases: The threat to biodiversity and the delivery of ecosystem services. Biodivers. Conserv. 2017, 26, 3167–3181. [Google Scholar] [CrossRef]
- Simova-Tosic, D.; Filev, S. Contribution to the knowledge of leaf miners of horse chestnut. Zast. Bilja 1985, 36, 235–239. [Google Scholar]
- Deschka, G.; Dimic, N. Cameraria ohridella n. sp. aus Macedonien, Jugoslavien (Lepidoptera, Lithocolletidae). Acta Entomol. Jugosl. 1986, 22, 11–23. [Google Scholar]
- Kirichenko, N.I.; Karpun, N.N.; Zhuravleva, E.N.; Shoshina, E.I.; Anikin, V.V.; Musolin, D.L. Invasion genetics of the horse-chestnut leaf miner, Cameraria ohridella (Lepidoptera: Gracillariidae), in European Russia: A case of successful involvement of citizen science in studying an alien insect pest. Insects 2023, 14, 117. [Google Scholar] [CrossRef] [PubMed]
- Tomiczek, C.; Krehan, H. The horsechestnut leafmining moth (Cameraria ohridella): A new pest in central Europe. J. Arboric. 1998, 24, 144–148. [Google Scholar]
- Gninenko, Y.I.; Orlinskiy, A.D. New phytophages of tree plantations. Plant Prot. Quarant. 2004, 4, 33. [Google Scholar]
- Golosova, M.A.; Gninenko, Y.I. The appearance of the Ohrid leafminer on horse chestnut in Moscow. Les. Vest. 2006, 2, 43–46. (In Russian) [Google Scholar]
- Beguinot, J. Comparing the ratios of leaf-acceptance prior to egg-laying among several species of leaf-mining moths upon Quercus sp. pl. (Lepidoptera: Gracillariidae, Tischeriidae). Entomol. Gener. 2009, 32, 145–154. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Haratym, W. Changes in leaf tissues of common horse chestnut (Aesculus hippocastanum L.) colonised by the horse-chestnut leaf miner (Cameraria ochridella Deschka and Dimic). Acta Agrobot. 2011, 64, 11–22. [Google Scholar] [CrossRef]
- Balder, H.; Jäckel, B.; Schomlling, S. Effects of the horse chestnut leaf-miner Cameraria ohridella Deschka & Dimic on the frost hardness of Aesculus hippocastanum L. In Proceedings of the 1st International Cameraria Symposium—Cameraria ohridella and Other Invasive Leaf-Miners in Europe, Prague, Czech Republic, 24–27 March 2004; IOCB: Prague, Czech Republic, 2004; p. 4. [Google Scholar]
- Tarwacki, G.; Bystrowski, C.; Celmer-Warda, K. Effect of sun-exposure of the horse chestnut (Aesculus hippocastanum L.) on the occurrence and number of parasitoids of the horse chestnut leafminer (Cameraria ohridella Deschka & Dimic) in central Poland in 2004–2006. Folia For. Pol. (Ser. A) 2012, 54, 56–62. [Google Scholar]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Bidart-Bouzat, M.G.; Imeh-Nathaniel, A. Global change effects on plant chemical defenses against insect herbivores. J. Integr. Plant Biol. 2008, 50, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Holoborodko, K.; Seliutina, O.; Alexeyeva, A.; Brygadyrenko, V.; Ivanko, I.; Shulman, M.; Pakhomov, O.; Loza, I.; Sytnyk, S.; Lovynska, V.; et al. The Impact of Cameraria ohridella (Lepidoptera, Gracillariidae) on the State of Aesculus hippocastanum Photosynthetic Apparatus in the Urban Environment. Int. J. Plant Biol. 2022, 13, 223–234. [Google Scholar] [CrossRef]
- Kulbat, K. The role of phenolic compounds in plant resistance. Biotechnol. Food Sci. 2016, 80, 97–108. [Google Scholar]
- Materska, M.; Pabich, M.; Sachadyn-Król, M.; Konarska, A.; Weryszko-Chmielewska, E.; Chilczuk, B.; Staszowska-Karkut, M.; Jackowska, I.; Dmitruk, M. The Secondary Metabolites Profile in Horse Chestnut Leaves Infested with Horse-Chestnut Leaf Miner. Molecules 2022, 27, 5471. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Haratym, W. Leaf micromorphology of Aesculus hippocastanum L. and damage caused by leaf-mining larvae of Cameraria ohridella Deschka and Dimic. Acta Agrobot. 2012, 65, 25–33. [Google Scholar] [CrossRef]
- Kehrli, P.; Bacher, S. Date of leaf litter removal to prevent emergence of Cameraria ohridella in the following spring. Entomol. Exp. Appl. 2003, 107, 159–162. [Google Scholar] [CrossRef]
- Pavan, F.; Barro, P.; Bernardinelli, I.; Gambon, N.; Zandigiacomo, P. Cultural control of Cameraria ohridella on horsechestnut in urban areas by removing fallen leaves in autumn. J. Arboric. 2003, 29, 253–258. [Google Scholar] [CrossRef]
- Floricel, M.; Mitrea, I.; Oltean, I.; Florian, T.; Varga, M.I.; Vasian, I.; Hulujan, I.B. The testing of some products in order to monitor the Cameraria ohridella Deschka-Dimic species (Lepidoptera: Gracilariidae). Agrolife Sci. J. 2018, 7, 53–60. [Google Scholar]
- Jagiełło, R.; Walczak, U.; Iszkuło, G.; Karolewski, P.; Baraniak, E.; Giertych, M.J. Impact of Cameraria ohridella on Aesculus hippocastanum growth and long-term effects of trunk injection with pesticides. Int. J. Pest Manag. 2019, 65, 33–43. [Google Scholar] [CrossRef]
- Plotnikova, L.S.; Aleksandrova, M.S.; Belyaeva, Y.E.; Nemova, E.M.; Ryabova, N.V.; Yakushina, E.I. Drevesnye rasteniya Glavnogo botanicheskogo sada imeni N. V. Tsitsina RAN: 60 let introduktsii. In Tree Plants of the N. V. Tsitsin Chief Botanical Garden RAS: 60 Years of Introduction; Demidov, A.S., Ed.; Science: Moscow, Russia, 2005; p. 586. [Google Scholar]
- Konarska, A.; Grochowska, M.; Haratym, W.; Tietze, M.; Weryszko-Chmielewska, E.; Lechowski, L. Changes in Aesculus hippocastanum leaves during development of Cameraria ohridella. Urban For. Urban Green. 2020, 56, 126793. [Google Scholar] [CrossRef]
- Šefrová, H.; Skuhravý, V. The larval morphology of Cameraria ohridella Deschka & Dimić compared with the genus Phyllonorycter Hübner (Lepidoptera, Gracillariidae). Acta Univ. Agric. Silvic. Mendel. Brun. 2000, 48, 23–30. [Google Scholar]
- Zunke, U.; Fernandez-Nunez, M.; Ilmberger, N.; Hofmeier, M.; Konitz, K.; Doobe, G. Zur Biologie und Populationsdynamik der Rosskastanienminiermotte und Ihrer Verbreitung in Hamburg. Cameraria ohridella, das HAM-CAM-Projekt 2001/04 in Hamburg, Hamburg. 2003; p. 38. Available online: http://cameraria.galk.de/hamburg.html (accessed on 9 February 2018).
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Marinova, G.; Batchvarov, V. Evaluation of the methods for determination of the free radical scavenging activity by DPPH. Bulg. J. Agric. Sci. 2011, 17, 11–24. [Google Scholar]
- Han, X.; Yang, K.; Gross, R.W. Multi-dimensional mass spectrometry-based shotgun lipidomics and novel strategies for lipidomic analyses. Mass Spectrom. Rev. 2012, 31, 134–178. [Google Scholar] [CrossRef] [PubMed]
- Bednova, O.V. Ohridsky miner Cameraria ohridella Deschka & Dimic: Features of invasion foci and prospects of biological population control. For. Bull. 2022, 26, 5–16. (In Russian) [Google Scholar]
- Kashtanova, O.A.; Tkachenko, O.B.; Kondratieva, V.V.; Voronkova, T.V.; Olekhnovich, L.S. Resistance of horse chestnut species (Aesculus L.) to ohrid miner or chestnut miner moth (Cameraria ohridella Deschka & Dimic). Bull. Moscow Soc. Natur. Biol. Ser. 2020, 125, 45–51. (In Russian) [Google Scholar]
- Cardé, R.T.; Elkinton, J.S. Field trapping with attractants: Methods and interpretation. In Techniques in Pheromone Research; Springer: New York, NY, USA, 1984; pp. 111–129. [Google Scholar]
- Šefrova, H.; Laštůvka, Z. Dispersal of the horse chestnut leaf miner, Cameraria ohridella Deschka & Dimić, 1986, in Europe: Its course, ways and causes (lepidoptera: Gracillariidae). Entomol. Zeits. 2001, 111, 194–198. [Google Scholar]
- Deschka, G. Schmetterlinge als Einwanderer. Stapfia 1995, 37, 77–128. [Google Scholar]
- Kalinová, B.; Svatoŝ, A.; Kindl, J.; Hovorka, O.; Hrdý, I.; Kuldova, J.; Hoskovec, M. Sex pheromone of horse-chestnut leafminer Cameraria ohridella and its use in a pheromone-based monitoring system. J. Chem. Ecol. 2003, 29, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Beguinot, J. The remarkably low leaf-selectivity prior to oviposition in the moth-pest Cameraria ohridella is not unique to this species within the genus Cameraria. Rev. Ecol. (Terre Vie) 2009, 64, 333–341. [Google Scholar] [CrossRef]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T. A meta-analysis of preference–performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef]
- Heitland, W.; Freise, J. Verbreitung der Roßkastanien-Miniermotte, Cameraria ohridella (Lep, Gracillariidae) in Deutschland. Distribution of the horse-chestnut leaf miner, Cameraria ohridella (Lep., Gracillariidae) in Germany. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2001, 13, 131–134. [Google Scholar]
- Forister, M.L.; Nice, C.C.; Fordyce, J.A.; Gompert, Z. Host range evolution is not driven by the optimization of larval performance: The case of Lycaeides melissa (Lepidoptera: Lycaenidae) and the colonization of alfalfa. Oecologia 2009, 160, 551–561. [Google Scholar] [CrossRef]
- Jagiełło, R.; Baraniak, E.; Guzicka, M.; Karolewski, P.; Łukowski, A.; Giertych, M.J. One step closer to understanding the ecology of Cameraria ohridella (Lepidoptera: Gracillariidae): The effects of light conditions. Eur. J. Entomol. 2019, 116, 42–51. [Google Scholar] [CrossRef]
- Birner, A.; Bohlander, F. Mine development of horse chestnut leaf-miner (Cameraria ohridella) on leafs exposed to sunlight or shade. In Cameraria ohridella and Other Invasive Leaf-Miners in Europe, Proceedings of the 1st International Cameraria Symposium, Prague, Czech Republic, 24–27 March 2004; Kalinova, B., Ed.; Institute of Organic Chemistry and Biochemistry ASCR: Prague, Czech Republic, 2004; p. 5. [Google Scholar]
- Barthlott, W.; Neinhuis, C. Purity of the sacred lotus or escape from contamination in biological surfaces. Planta 1997, 202, 1–8. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Isaias, R.M.S. Redifferentiation of leaflet tissues during midrib gall development in Copaifera langsdorffii (Fabaceae). S. Afr. J. Bot. 2010, 76, 239–248. [Google Scholar] [CrossRef]
- Dardeau, F.; Pointeau, S.; Ameline, A.; Laurans, F.; Cherqui, A.; Lieutier, F.; Sallé, A. Host manipulation by a herbivore optimizes its feeding behaviour. Anim. Behav. 2014, 95, 49–56. [Google Scholar] [CrossRef]
- Narawatthana, S. The Regulation of Leaf Thickness in Rice (Oryza sativa L.). Ph.D. Thesis, University of Sheffield, Sheffield, UK, 2013. [Google Scholar]
- Chen, L.L.; Zhang, K.; Gong, X.C.; Wang, H.Y.; Gao, Y.H.; Wang, X.Q.; Hu, Y.G. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 2020, 19, 108–119. [Google Scholar] [CrossRef]
- Molano-Flores, B. Herbivory and calcium concentrations affect calcium oxalate crystal formation in leaves of Sida (Malvaceae). Ann. Bot. 2001, 88, 387–391. [Google Scholar] [CrossRef]
- Nakata, P.A. Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar] [CrossRef]
- Bernards, M.; Båstrup-Spohr, L. Phenylpropanoid metabolism induced by wounding and insect herbivory. In Induced Resistance to Herbivory; Schaller, A., Ed.; Springer: New York, NY, USA, 2008; pp. 189–211. [Google Scholar]
- Pagare, S.; Bhatia, M.; Tripathi, N.; Pagare, S.; Bansal, Y.K. Secondary metabolites of plants and their role: Overview. Curr. Trends Biotechnol. Pharm. 2015, 9, 293–304. [Google Scholar]
- Seliutina, O.V.; Shupranova, L.V.; Holoborodko, K.K.; Shulman, M.V.; Bobylev, Y.P. Effect of Cameraria ohridella on accumulation of proteins, peroxidase activity and composition in Aesculus hippocastanum leaves. Regul. Mech. Biosyst. 2020, 11, 299–304. [Google Scholar] [CrossRef]
- Holoborodko, K.K.; Seliutina, O.V.; Ivanko, I.A.; Alexeyeva, A.A.; Shulman, M.V.; Pakhomov, O.Y. Effect of Cameraria ohridella feeding on Aesculus hippocastanum photosynthesis. Regul. Mech. Biosyst. 2021, 12, 346–352. [Google Scholar] [CrossRef]
- Salminen, J.P.; Karonen, M. Chemical ecology of tannins and other phenolics: We need a change in approach. Func. Ecol. 2011, 25, 325–338. [Google Scholar] [CrossRef]
- Oszmiański, J.; Kalisz, S.; Aneta, W. The content of phenolic compounds in leaf tissues of white (Aesculus hippocastanum L.) and red horse chestnut (Aesculus carea H.) colonized by the horse chestnut leaf miner (Cameraria ohridella Deschka & Dimić). Molecules 2014, 19, 14625–14636. [Google Scholar]
- Kopacka, M.; Zemek, R. Spatial variability in the level of infestation of the leaves of horse chestnut by the horse chestnut leaf miner, Cameraria ohridella (Lepidoptera: Gracillariidae) and in the number of adult moths and parasitoids emerging from leaf litter in an urban environment. Eur. J. Entomol. 2017, 114, 42. [Google Scholar]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P.; Stavroulaki, V.; Sumbele, S. “Carbon gain vs. water saving, growth vs. defence”: Two dilemmas with soluble phenolics as a joker. Plant Sci. 2014, 227, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Moreira, X.; Zas, R.; Sampedro, L. Differential allocation of constitutive and induced chemical defenses in pine tree juveniles: A test of the optimal defense theory. PLoS ONE 2012, 7, e34006. [Google Scholar] [CrossRef]
- Melo De Pinna, G.F.A.; Kraus, J.E.; de Menezes, N.L. Morphology and anatomy of leaf mine in Richterago riparia Roque (Asteraceae) in the campos rupestres of Serra do Cipó, Brazil. Braz. J. Biol. 2002, 62, 179–185. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Peak Height, % of Scale | |
|---|---|---|
| Sunny Side | Shady Side | |
| Polyphenols | ||
| 2-butene-1,4-diol | 0.21 a | 0.05 bc |
| L-(-)-arabitol | 1.70 a | 0.29 c |
| D-fucitol | 0.95 a | 0.44 b |
| 1,5-anhydroglucitol | 1.34 a | 0.06 c |
| D-mannitol | 0.01 bc | 0.55 a |
| 1,5-angidroglucitol | 8.71 b | 10.33 a |
| scillo-inositol | 1.80 a | 1.26 ab |
| galactinol | 10.59 bc | 15.1 a |
| D-glucitol | 0.94 a | 0.60 b |
| maltitol | 0.01 c | 3.96 a |
| Carboxylic esters | ||
| D-(+)- ribono-1.4-lactone | 0.23 ab | 0.31 a |
| gluconolactone | 1.25 a | 0.01 c |
| Organic acid | ||
| glucopyranuronic acid | 0.14 b | 0.49 a |
| ribonic acid | 0.97 a | 0.36 bc |
| quininic acid | 7.50 bc | 11.75 a |
| gluonic acid | 7.39 ab | 8.29 a |
| gallic acid | 0.42 a | 0.12 b |
| palmitic acid | 0.27 a | 0.01 b |
| D-(+)-galacturonic acid | 0.49 a | 0.03 bc |
| 4-aminosalicylic acid | 0.01 c | 2.03 a |
| 5-aminosalicylic acid | 0.01 b | 0.31 a |
| sinapinic acid | 0.36 a | 0.27 a |
| betulinic acid | 0.44 a | 0.41 a |
| Sugar derivatives | ||
| D-erythro-2-pentulose | 0.60 a | 0.41 b |
| methyl-a-D-glucofuranoside | 0.55 ab | 0.70 a |
| D-psicofuranose | 9.15 a | 9.45 a |
| D-(-)-tagatofuranose | 0.01 b | 0.48 a |
| DL- arabinofuranoside | 2.66 ab | 3.04 a |
| methyl galactoside | 6.12 a | 5.25 ab |
| b-D-(+)-talophyranose | 1.87 ab | 2.40 a |
| talofuranose | 1.63 a | 1.30 ab |
| deoxyglucose | 0.12 ab | 0.19 a |
| a-D-ribofuranose | 0.12 b | 0.28 a |
| L-sorbopyranose | 0.09 ab | 0.15 a |
| glucosylspingosine | 6.67 a | 2.31 bc |
| D-turanose | 0.01 c | 3.28 a |
| methyl-a-N-acetyl-D-galactoside | 5.20 a | 0.01 d |
| D-(-)-sorbofuranose | 1.55 a | 1.85 a |
| D-trehalose | 2.52 a | 1.50 b |
| D-(-)-xylophyranose | 5.63 a | 0.01 d |
| 1-c-octylhexopyranose | 1.95 a | 2.13 a |
| DL-arabinopyranose | 2.54 a | 2.62 a |
| b-lyxopyranose | 0.01 b | 0.44 a |
| a-D-lactose | 0.01 b | 0.31 a |
| D-ribofuranose | 0.38 ab | 0.72 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogoutdinova, L.R.; Tkacheva, E.V.; Konovalova, L.N.; Tkachenko, O.B.; Olekhnovich, L.S.; Gulevich, A.A.; Baranova, E.N.; Shelepova, O.V. Effect of Sun Exposure of the Horse Chestnut (Aesculus hippocastanum L.) on the Occurrence and Number of Cameraria ohridella (Lepidoptera: Gracillariidae). Forests 2023, 14, 1079. https://doi.org/10.3390/f14061079
Bogoutdinova LR, Tkacheva EV, Konovalova LN, Tkachenko OB, Olekhnovich LS, Gulevich AA, Baranova EN, Shelepova OV. Effect of Sun Exposure of the Horse Chestnut (Aesculus hippocastanum L.) on the Occurrence and Number of Cameraria ohridella (Lepidoptera: Gracillariidae). Forests. 2023; 14(6):1079. https://doi.org/10.3390/f14061079
Chicago/Turabian StyleBogoutdinova, Liliya R., Ekaterina V. Tkacheva, Ludmila N. Konovalova, Oleg B. Tkachenko, Ludmila S. Olekhnovich, Alexander A. Gulevich, Ekaterina N. Baranova, and Olga V. Shelepova. 2023. "Effect of Sun Exposure of the Horse Chestnut (Aesculus hippocastanum L.) on the Occurrence and Number of Cameraria ohridella (Lepidoptera: Gracillariidae)" Forests 14, no. 6: 1079. https://doi.org/10.3390/f14061079