
Long-Term Nitrogen Addition Exerts Minor Effects on Microbial Community but Alters Sensitive Microbial Species in a Subtropical Natural Forest
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Soil Sample Collection
2.3. Measurement of Soil Properties
2.4. DNA Extraction and Sequencing
2.5. Data Analysis
3. Results
3.1. Soil Properties
3.2. Microbial Community Diversity
3.3. Microbial Community Composition and Structure
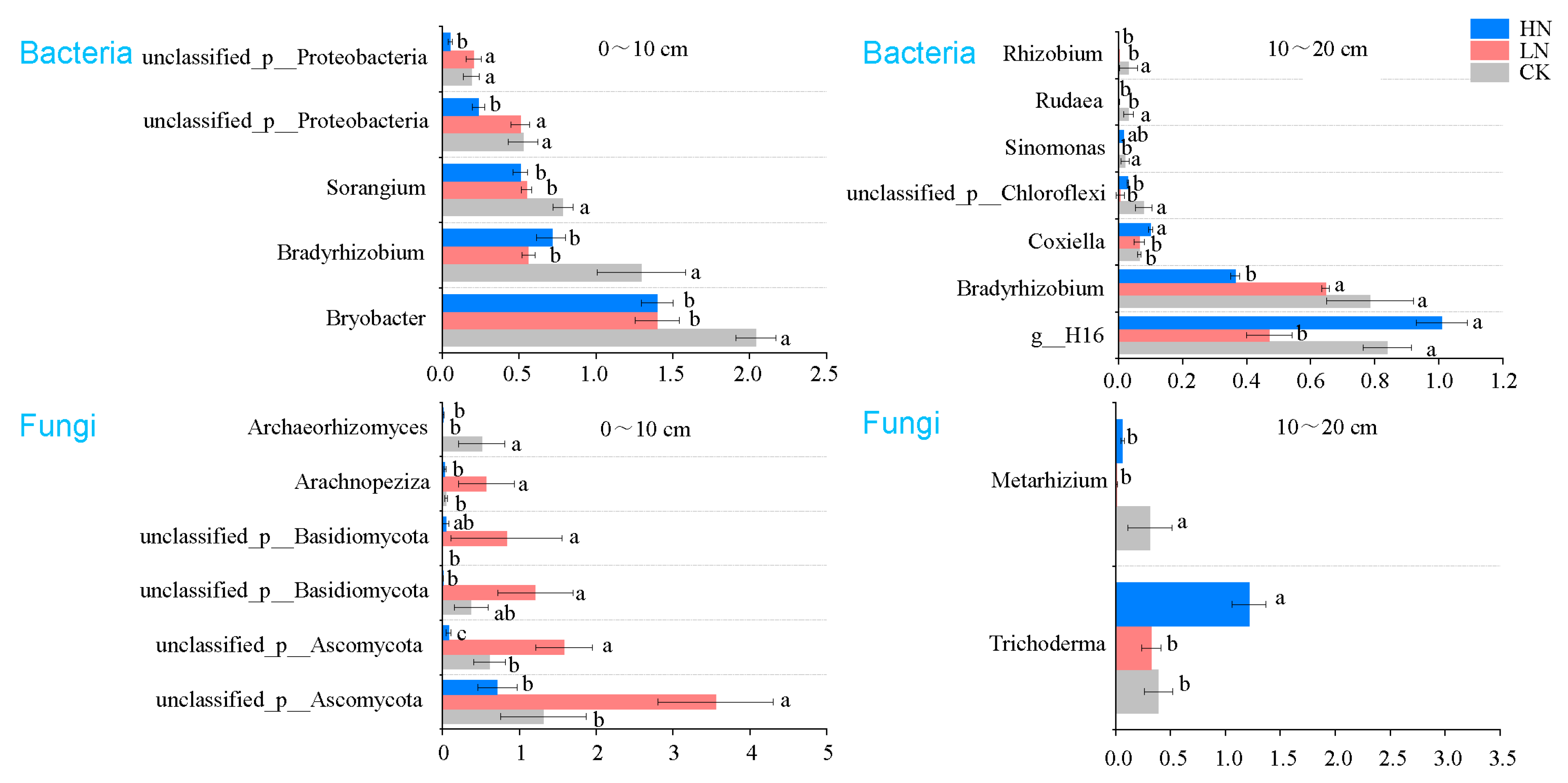
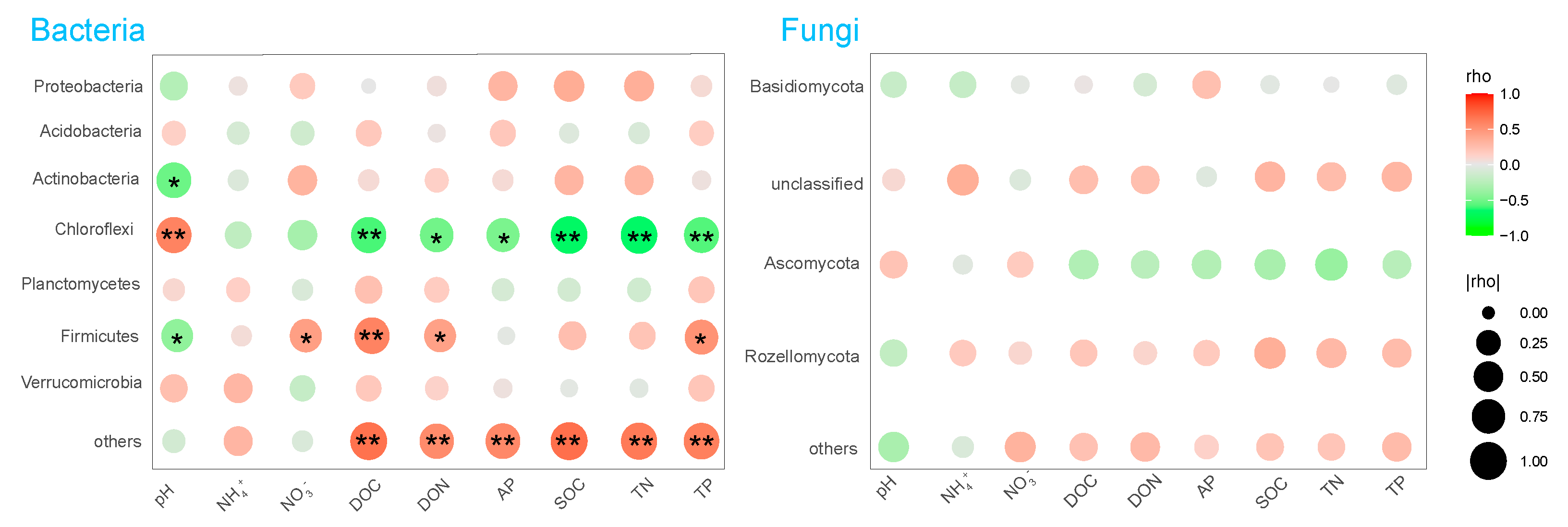
3.4. Sensitive Microbial Groups
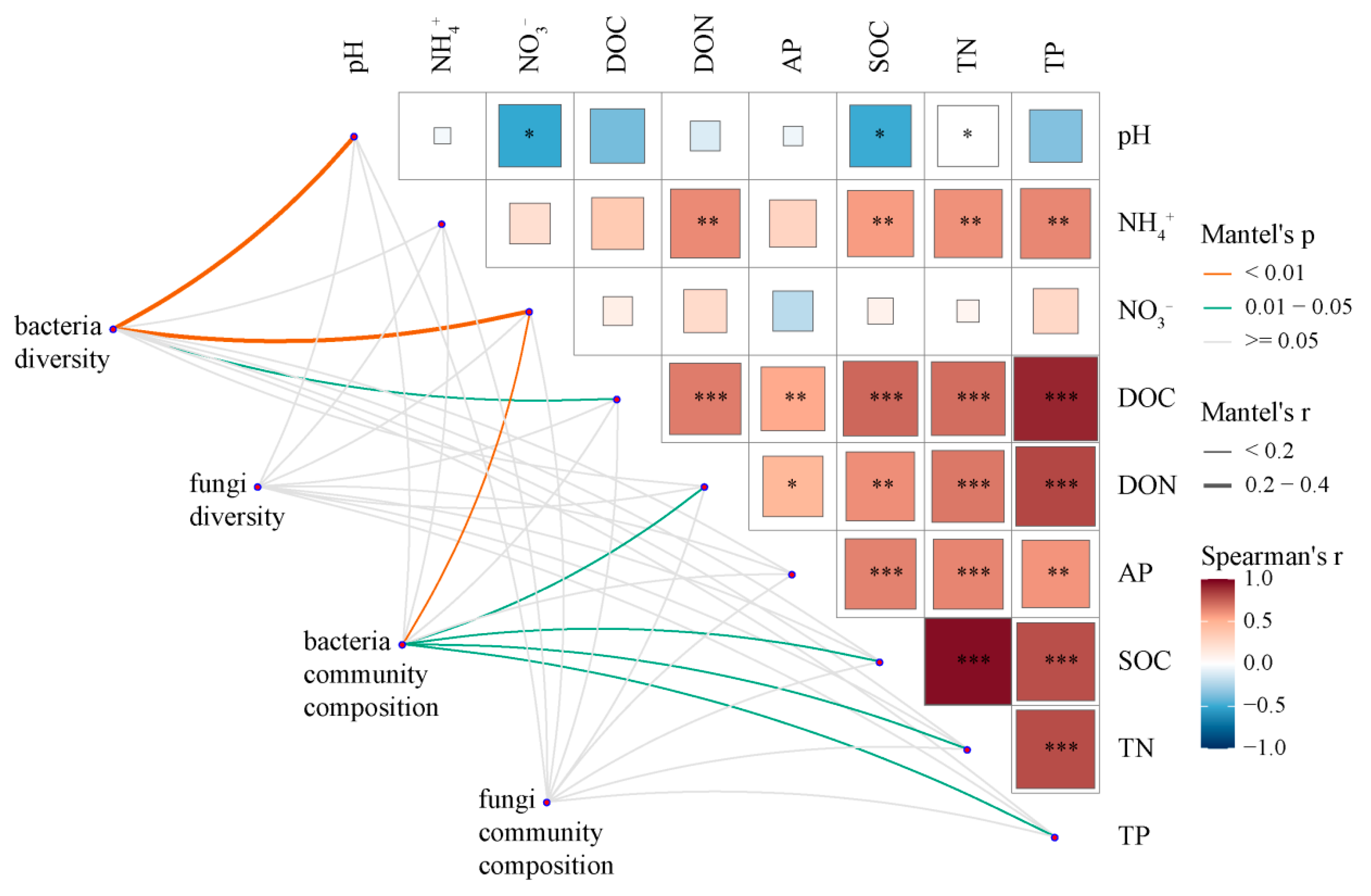
3.5. Relationships between Microbial Community Diversity and Structure and Soil Properties
4. Discussions
4.1. Effects of N Deposition on Soil Properties
4.2. Effects of N Deposition on Soil Microbial Community Diversity, Composition, and Structure
4.3. Effects of N Deposition on Microbial Sensitive Groups
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ackerman, D.; Millet, D.B.; Chen, X. Global estimates of inorganic nitrogen deposition across four decades. Glob. Biogeochem. Cycles 2019, 33, 100–107. [Google Scholar] [CrossRef]
- He, W.; Zhang, M.; Jin, G.; Sui, X.; Zhang, T.; Song, F. Effects of nitrogen deposition on nitrogen-mineralizing enzyme activity and soil microbial community structure in a Korean pine plantation. Microb. Ecol. 2021, 81, 410–424. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Emmett, B.A.; Britton, A.J.; Caporn, S.J.M.; Dise, N.B.; Helliwell, R.; Jones, L.; Leake, J.R.; Leith, I.D.; Sheppard, L.J.; et al. Impacts of atmospheric nitrogen deposition: Responses of multiple plant and soil parameters across contrasting ecosystems in long-term field experiments. Glob. Chang. Biol. 2012, 18, 1197–1215. [Google Scholar] [CrossRef]
- Stevens, C.J.; David, T.I.; Storkey, J.; Fox, C. Atmospheric nitrogen deposition in terrestrial ecosystems: Its impact on plant communities and consequences across trophic levels. Funct. Ecol. 2018, 32, 1757–1769. [Google Scholar] [CrossRef]
- Dai, Z.M.; Xiong, X.Q.; Zhu, H.; Xu, H.J.; Leng, P.; Li, J.H.; Tang, C.; Xu, J.M. Association of biochar properties with changes in soil bacterial, fungal and fauna communities and nutrient cycling processes. Biochar 2021, 3, 239–254. [Google Scholar] [CrossRef]
- Liu, X.; Duan, L.; Mo, J.; Du, E.; Shen, J.; Lu, X.; Zhang, Y.; Zhou, X.; He, C.; Zhang, F. Nitrogen deposition and its ecological impact in China: An overview. Environ. Pollut. 2011, 159, 2251–2264. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J. The global nitrogen-phosphorus imbalance. Science 2022, 375, 266–267. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, G.; Chen, H.; Chen, C.; Wang, J.; Ai, S.; Wei, D.; Li, D.; Ma, B.; Tang, C.; et al. Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. ISME J. 2020, 14, 757–770. [Google Scholar] [CrossRef]
- Moore, J.A.; Anthony, M.A.; Pec, G.J.; Trocha, L.K.; Trzebny, A.; Geyer, K.M.; van Diepen, L.T.A.; Frey, S.D. Fungal community structure and function shifts with atmospheric nitrogen deposition. Glob. Chang. Biol. 2021, 27, 1349–1364. [Google Scholar] [CrossRef]
- Zuccarini, P.; Asensio, D.; Sardans, J.; Ogaya, R.; Peñuelas, J. Changes in soil enzymatic activity in a P-limited Mediterranean shrubland subject to experimental nitrogen deposition. Appl. Soil Ecol. 2021, 168, 104159. [Google Scholar] [CrossRef]
- Papanikolaou, N.; Britton, A.J.; Helliwell, R.C.; Johnson, D. Nitrogen deposition, vegetation burning and climate warming act independently on microbial community structure and enzyme activity associated with decomposing litter in low-alpine heath. Glob. Chang. Biol. 2010, 16, 3120–3132. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L. Microbial community response to nitrogen deposition in northern forest ecosystems. Soil Biol. Biochem. 2004, 36, 1443–1451. [Google Scholar] [CrossRef]
- Wang, C.; Lu, X.; Mori, T.; Mao, Q.; Zhou, K.; Zhou, G.; Nie, Y.; Mo, J. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Michel, K.; Pfeffer, M. Soil microbial community structure in European forests in relation to forest type and atmospheric nitrogen deposition. Plant Soil 2010, 343, 37–50. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, H.Y.H.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef]
- Sha, M.H.; Xu, J.; Zheng, Z.C.; Fa, K.Y. Enhanced atmospheric nitrogen deposition triggered little change in soil microbial diversity and structure in a desert ecosystem. Glob. Ecol. Conserv. 2021, 31, e01879. [Google Scholar] [CrossRef]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil microbial resources for improving fertilizers efficiency in an integrated plant nutrient management system. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.K.; Gao, Q.Z.; Ganjurjav, H.; Wang, X.X.; Geng, W. “Rare biosphere” plays important roles in regulating soil available nitrogen and plant biomass in alpine grassland ecosystems under climate changes. Agric. Ecosyst. Environ. 2019, 279, 187–193. [Google Scholar] [CrossRef]
- Li, Y.; Bezemer, T.M.; Yang, J.; Lü, X.; Li, X.; Liang, W.; Han, X.; Li, Q. Changes in litter quality induced by N deposition alter soil microbial communities. Soil Biol. Biochem. 2019, 130, 33–42. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.R.; Zhang, X.; Mao, Q.G.; Li, X.Z.; You, Y.M.; Wang, J.X.; Zheng, M.H.; Zhang, W.; Lu, X.K.; et al. Nitrogen addition reduces soil bacterial richness, while phosphorus addition alters community composition in an old-growth N rich tropical forest in Southern China. Soil Biol. Biolchem. 2018, 127, 22–30. [Google Scholar] [CrossRef]
- Xia, Z.W.; Yang, J.Y.; Sang, C.P.; Wang, X.; Sun, L.F.; Jiang, P.; Wang, C.; Bai, E. Phosphorus reduces negative effects of nitrogen addition on soil microbial communities and functions. Microorganisms 2020, 8, 1828. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; Qiu, X.R.; Sun, Y.; Liu, S.N.; Hu, H.L.; Xie, J.L.; Chen, G.; Xiao, Y.L.; Tang, Y.; Tu, L.H. C: N: P stoichiometry responses to 10 years of nitrogen addition differ across soil components and plant organs in a subtropical Pleioblastus amarus forest. Sci. Total Environ. 2021, 796, 148925. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, H.; An, S.; Bhople, P.; Davlatbekov, F. Geographic distance and soil microbial biomass carbon drive biogeographical distribution of fungal communities in Chinese Loess Plateau soils. Sci. Total Environ. 2019, 660, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.M.; Lan, Z.C.; Hu, S.J.; Bai, Y.F. Effects of nitrogen enrichment on belowground communities in grassland: Relative role of soil nitrogen availability vs. soil acidification. Soil Biol. Biochem. 2015, 89, 99–108. [Google Scholar] [CrossRef]
- Jia, M.Q.; Gao, Z.W.; Huang, J.; Li, J.; Liu, Z.Y.; Zhang, G.G.; Liu, F.H.; Wang, Z.W.; Han, G.D. Soil bacterial community is more sensitive than fungal community to nitrogen supplementation and climate warming in Inner Mongolian desert steppe. J. Soils Sediments 2022, 23, 405–421. [Google Scholar] [CrossRef]
- Elizabeth, T.B.; Carly, J.S. Nitrogen deposition and climate: An integrated synthesis. Trends Ecol. Evol. 2022, 37, 541–552. [Google Scholar]
- Zelenev, V.V.; Bruggen, A.H.C.V.; Semenov, A.M. Modeling Wave-like dynamics of oligotrophic and copiotrophic bacteria along wheat roots in response to nutrient input from a growing root tip. Ecol. Model. 2005, 188, 404–417. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Fan, Y.X.; Zhong, X.J.; Lin, T.C.; Lyu, M.K.; Wang, M.H.; Hu, W.F.; Yang, Z.J.; Chen, G.S.; Guo, J.F.; Yang, Y.S. Effects of nitrogen addition on DOM-induced soil priming effects in a subtropical plantation forest and a natural forest. Biol Fertil Soils 2019, 56, 205–216. [Google Scholar] [CrossRef]
- Yan, Z.B.; Kim, N.; Han, W.X.; Guo, Y.L.; Han, T.S.; Du, E.Z.; Fang, J.Y. Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant Soil 2015, 388, 147–155. [Google Scholar] [CrossRef]
- Zhao, X.X.; Tian, Q.X.; Huang, L.; Lin, Q.L.; Wu, J.J.; Liu, F. Fine-root functional trait response to nitrogen deposition across forest ecosystems: A meta-analysis. Sci. Total Environ. 2022, 844, 157111. [Google Scholar] [CrossRef]
- Fan, Y.X.; Lin, F.; Yang, L.M.; Zhong, X.; Wang, M.H.; Zhou, J.C.; Chen, Y.M.; Yang, Y.S. Decreased soil organic P fraction associated with ectomycorrhizal fungal activity to meet increased P demand under N application in a subtropical forest ecosystem. Biol. Fertil. Soils 2018, 54, 149–161. [Google Scholar] [CrossRef]
- Zhou, J.C.; Liu, X.F.; Xie, J.S.; Lyu, M.; Zheng, Y.; You, Z.T.; Fan, Y.X.; Lin, C.F.; Chen, G.S.; Chen, Y.M.; et al. Nitrogen addition affects soil respiration primarily through changes in microbial community structure and biomass in a subtropical natural forest. Forests 2019, 10, 435. [Google Scholar] [CrossRef]
- Zhong, X.L.; Li, J.T.; Li, X.J.; Ye, Y.C.; Liu, S.S.; Hallett, P.D.; Ogden, M.R.; Naveed, M. Physical protection by soil aggregates stabilizes soil organic carbon under simulated N deposition in a subtropical forest of China. Geoderma 2016, 185, 323–332. [Google Scholar] [CrossRef]
- Pereira, E.A.O.; Melo, V.F.; Abate, G.; Masini, J.C. Adsorption of glyphosate on Brazilian subtropical soils rich in iron and aluminum oxides. J. Environ. Sci. Health B 2019, 54, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Bowman, W.D.; Cleveland, C.C.; Halada, L.; Hresko, J.; Baron, J.S. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Lucas, R.W.; Klaminder, J.; Futter, M.N.; Bishop, K.H.; Egnell, G.; Laudon, H.; Högberg, P. A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams. For. Ecol. Manag. 2011, 262, 95–104. [Google Scholar] [CrossRef]
- Guan, B.; Xie, B.H.; Yang, S.S.; Hou, A.X.; Chen, M.; Han, G.X. Effects of five years’ nitrogen deposition on soil properties and plant growth in a salinized reed wetland of the Yellow River Delta. Ecol. Eng. 2019, 136, 160–166. [Google Scholar] [CrossRef]
- Turner, B.L.; Blackwell, M.S.A. Isolating the influence of pH on the amounts and forms of soil organic phosphorus. Eur. J. Soil Sci. 2013, 64, 249–259. [Google Scholar] [CrossRef]
- Crowley, K.F.; McNeil, B.E.; Lovett, G.M.; Canham, C.D.; Driscoll, C.T.; Rustad, L.E.; Denny, E.; Hallett, R.A.; Arthur, M.A.; Boggs, J.L.; et al. Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the northeastern United States? Ecosystems 2012, 15, 940–957. [Google Scholar] [CrossRef]
- Fan, Y.X.; Yang, L.M.; Zhong, X.J.; Yang, Z.J.; Lin, Y.Y.; Guo, J.F.; Chen, G.S.; Yang, Y.S. N addition increased microbial residual carbon by altering soil P availability and microbial composition in a subtropical Castanopsis forest. Geoderma 2020, 375, 114470. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, T.; Luo, Y.; Liu, J.; Lu, X.; Ye, Q.; Wang, S.; Huang, J.; Mao, Q.; Mo, J.; et al. Temporal patterns of soil carbon emission in tropical forests under long-term nitrogen deposition. Nat. Geosci. 2022, 15, 1002–1010. [Google Scholar] [CrossRef]
- Maaroufi, N.I.; Nordin, A.; Palmqvist, K.; Hasselquist, N.J.; Forsmark, B.; Rosenstock, N.P.; Wallander, H.; Gundale, M.J. Anthropogenic nitrogen enrichment enhances soil carbon accumulation by impacting saprotrophs rather than ectomycorrhizal fungal activity. Glob. Chang. Biol. 2019, 25, 2900–2914. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Y.; Zhu, Y.J.; Wang, B.; Liu, D.; Bai, H.; Jin, L.; Wang, B.T.; Ruan, H.H.; Mao, L.F.; Jin, F.J.; et al. Effects of nitrogen addition on rhizospheric soil microbial communities of poplar plantations at different ages. For. Ecol. Manag. 2021, 494, 119328. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Baath, E. Fungal and bacterial growth responses to N fertilization and pH in the 150-year ‘Park Grass’ UK grassland experiment: N and pH influence on microbial growth in grassland soils. FEMS Microbiol. Ecol. 2011, 76, 89–99. [Google Scholar] [CrossRef]
- Yin, C.T.; Jones, K.L.; Peterson, D.E.; Garrett, K.A.; Hulbert, S.H.; Paulitz, T.C. Members of soil bacterial communities sensitive to tillage and crop rotation. Soil Biol. Biochem. 2010, 42, 2111–2118. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McNulty, S.; Fernandez, I.J.; Boggs, J.; Schlesinger, W.H. Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments. For. Ecol. Manag. 2006, 222, 459–468. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Soong, J.L.; Fuchslueger, L.; Maranon-Jimenez, S.; Torn, M.S.; Janssens, I.A.; Penuelas, J.; Richter, A. Microbial carbon limitation: The need for integrating microorganisms into our understanding of ecosystem carbon cycling. Glob. Chang. Biol. 2019, 26, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Martins da Costa, E.; Azarias Guimarães, A.; Soares de Carvalho, T.; Louzada Rodrigues, T.; Rose de Almeida Ribeiro, P.; Lebbe, L.; Willems, A.; Fatima, M.; de Souza Moreira, F.M. Bradyrhizobium forestalis sp. nov., an efficient nitrogen-fixing bacterium isolated from nodules of forest legume species in the Amazon. Arch. Microbiol. 2018, 200, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Jordan, D.; Mcdonald, G.A. Effect of phosphate-solubilizing bacteria and vesiculararbuscular mycorrhizae on tomato growth and soil microbial activity. Biol. Fertil. Soils 1997, 26, 79–87. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Kulichevskaya, I.S.; Huber, K.J.; Overmann, J. Defining the taxonomic status of described subdivision 3 Acidobacteria: Proposal of Bryobacteraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 498–501. [Google Scholar] [CrossRef] [PubMed]
- Menkis, A.; Urbina, H.; James, T.Y.; Rosling, A. Archaeorhizomyces borealis sp. nov. and a sequence-based classification of related soil fungal species. Fungal Biol. 2014, 118, 943–955. [Google Scholar] [CrossRef]
- Rosling, A.; Cox, F.; Cruz-Martinez, K.; Ihrmark, K.; Grelet, G.A.; Lindahl, B.D.; Menkis, A.; James, T.Y. Archaeorhizomycetes: Unearthing an ancient class of ubiquitous soil fungi. Science 2011, 333, 876–879. [Google Scholar] [CrossRef]
- St Leger, R.J.; Wang, J.B. Metarhizium: Jack of all trades, master of many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef]
- Zhan, S.W.; Gan, Y.T.; Xu, B.L.; Xue, Y.Y. The parasitic and lethal effects of Trichoderma longibrachiatum against Heterodera avenae. Biol. Control 2014, 72, 1–8. [Google Scholar] [CrossRef]
- Maeda, K.; Spor, A.; Edel-Hermann, V.; Heraud, C.; Breuil, M.C.; Bizouard, F.; Toyoda, S.; Yoshida, N.; Steinberg, C.; Philippot, L. N2O production, a widespread trait in fungi. Sci. Rep. 2015, 5, 9697. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | 0–10 cm Layer | 10–20 cm Layer | ||||
|---|---|---|---|---|---|---|
| CT | LN | HN | CT | LN | HN | |
| pH | 4.03 ± 0.05 a | 3.88 ± 0.18 a | 3.88 ± 0.01 a | 4.14 ± 0.08 a | 4.10 ± 0.11 a | 4.08 ± 0.05 a |
| NH4+ (mg kg−1) | 24.15 ± 3.57 a | 27.79 ± 8.82 a | 23.21 ± 7.39 a | 18.50 ± 3.41 a | 16.62 ± 4.63 a | 24.84 ± 9.91 a |
| NO3− (mg kg−1) | 1.67 ± 0.46 c | 5.70 ± 1.09 b | 7.91 ± 1.69 a | 1.22 ± 0.20 c | 3.32 ± 0.50 b | 6.91 ± 0.61 a |
| DOC (mg kg−1) | 92.04 ± 34.23 a | 96.76 ± 25.84 a | 75.68 ± 24.50 a | 42.22 ± 7.84 a | 31.34 ± 12.39 a | 33.93 ± 8.79 a |
| DON (mg kg−1) | 18.48 ± 4.86 a | 15.68 ± 4.41 a | 15.03 ± 6.63 a | 4.11 ± 2.93 a | 3.08 ± 8.03 a | 12.15 ± 4.76 a |
| AP (mg kg−1) | 6.07 ± 1.05 a | 3.61 ± 0.75 b | 2.43 ± 0.41 b | 2.43 ± 0.98 a | 2.66 ± 0.77 a | 2.26 ± 0.77 a |
| SOC (g kg−1) | 33.46 ± 7.70 a | 35.41 ± 5.25 a | 31.37 ± 4.14 a | 15.26 ± 1.54 a | 15.38 ± 4.72 a | 14.37 ± 2.67 a |
| TN (g kg−1) | 2.36 ± 0.31 a | 2.41 ± 0.51 a | 2.34 ± 0.40 a | 1.09 ± 0.07 a | 1.06 ± 0.18 a | 1.07 ± 0.13 a |
| TP (g kg−1) | 0.22 ± 0.02 a | 0.20 ± 0.02 a | 0.19 ± 0.02 a | 0.15 ± 0.02 a | 0.14 ± 0.02 a | 0.14 ± 0.02 a |
| Soil Layer | Treatment | Bacteria | Fungi | ||
|---|---|---|---|---|---|
| Chao 1 | Shannon | Chao 1 | Shannon | ||
| 0–10 cm | CT | 1601.16 ± 149.40 a | 5.71 ± 0.12 ab | 519.47 ± 146.66 ab | 2.06 ± 0.72 ab |
| LN | 1598.81 ± 139.02 a | 5.74 ± 0.14 a | 680.28 ± 42.48 a | 3.07 ± 0.65 a | |
| HN | 1424.23 ± 30.98 a | 5.51 ± 0.09 b | 440.25 ± 123.83 b | 1.25 ± 0.53 b | |
| 10–20 cm | CT | 1677.31 ± 80.70 a | 5.51 ± 0.22 a | 594.92 ± 108.37 a | 2.48 ± 0.82 a |
| LN | 1605.78 ± 84.45 ab | 5.50 ± 0.12 a | 514.40 ± 166.28 a | 2.27 ± 0.85 a | |
| HN | 1528.32 ± 36.59 b | 5.48 ± 0.22 a | 605.96 ± 76.50 a | 3.35 ± 0.84 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, M.; Zhong, X.; Xia, Y.; Xu, L.; Zeng, Q.; Yang, L.; Fan, Y. Long-Term Nitrogen Addition Exerts Minor Effects on Microbial Community but Alters Sensitive Microbial Species in a Subtropical Natural Forest. Forests 2023, 14, 928. https://doi.org/10.3390/f14050928
He M, Zhong X, Xia Y, Xu L, Zeng Q, Yang L, Fan Y. Long-Term Nitrogen Addition Exerts Minor Effects on Microbial Community but Alters Sensitive Microbial Species in a Subtropical Natural Forest. Forests. 2023; 14(5):928. https://doi.org/10.3390/f14050928
Chicago/Turabian StyleHe, Min, Xiaojian Zhong, Yun Xia, Linglin Xu, Quanxin Zeng, Liuming Yang, and Yuexin Fan. 2023. "Long-Term Nitrogen Addition Exerts Minor Effects on Microbial Community but Alters Sensitive Microbial Species in a Subtropical Natural Forest" Forests 14, no. 5: 928. https://doi.org/10.3390/f14050928