
Seasonal Cambial Activity and Formation of Secondary Phloem and Xylem in White Oaks (Quercus alba L.)
 , ,
, , {kind=link}
{kind=link}
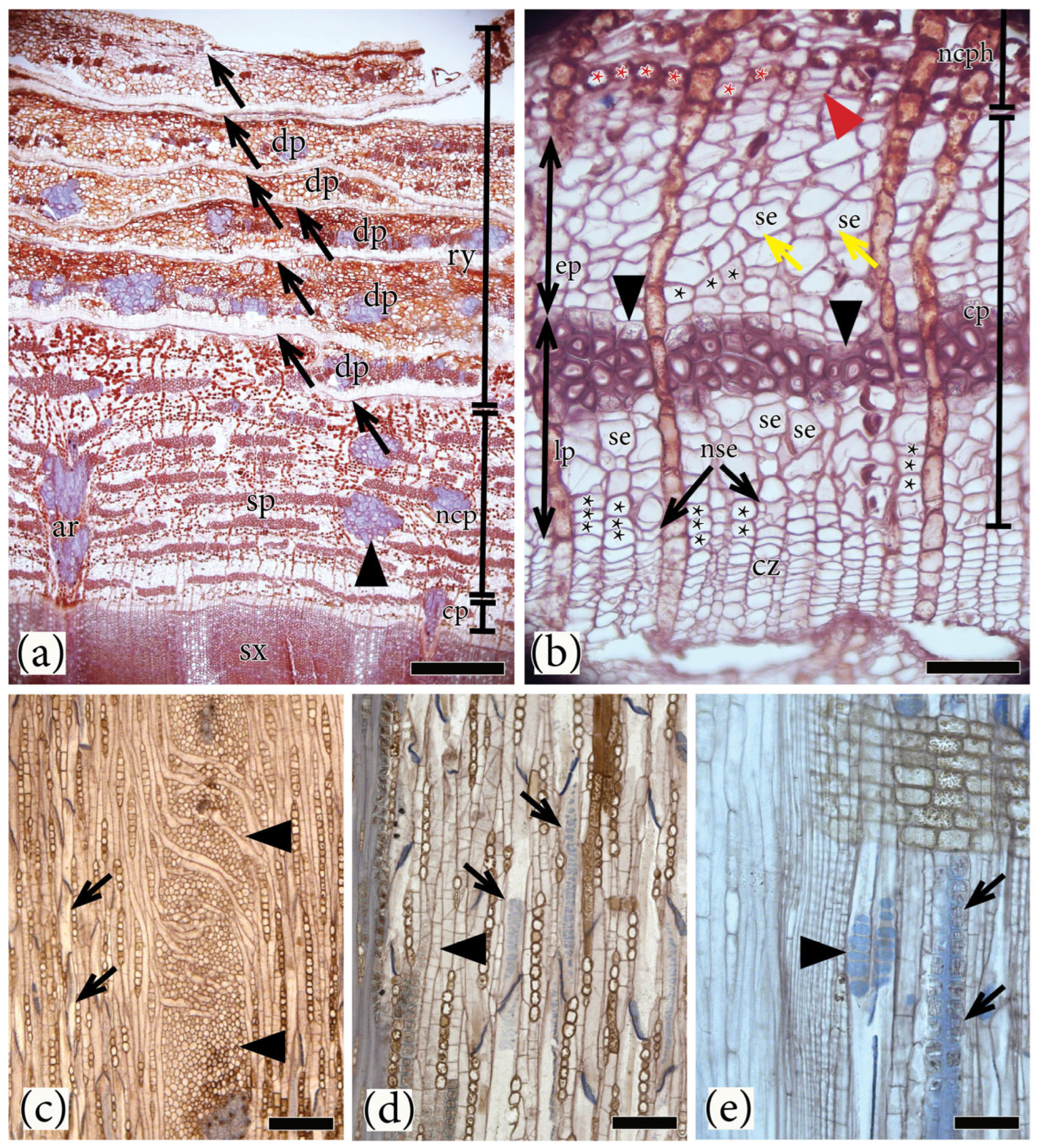{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Area of Study and Climate
2.2. Trees Sampled
2.3. Anatomical Criteria and Equipment Used
3. Results
3.1. General Description of the Wood and Bark
3.1.1. Wood Anatomy of Quercus alba
3.1.2. Bark Anatomy of Quercus alba
3.2. Seasonal Cycle of the Cambium, Wood, and Secondary Phloem Development
4. Discussion
4.1. Latewood and Late Phloem Are Produced While Environmental Conditions Are Still Favorable
4.2. Synchronicity in the Formation of Cambial Products and Growth Rings
4.3. Sieve Elements’ Longevity, Phloem and Wood Anatomical Features
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlquist, S. Comparative Wood Anatomy: Systematic, Ecological, and Evolutionary Aspects of Dicotyledon Wood; Springer: Berlin, Germany, 2001. [Google Scholar]
- Larson, P.R. The Vascular Cambium: Development and Structure; Springer: Heidelberg, Germany, 1994. [Google Scholar]
- Fritts, H.C. Growth rings of trees: Their correlation with climate. Science 1966, 154, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, R.; Vetter, R.E. Tree-Ring Analysis: Biological, Methodological and Environmental Aspects; CABI Publishing: New York, NY, USA, 1999. [Google Scholar]
- Silva, M.S.; Funch, L.S.; Silva, L.B. The growth ring concept: Seeking a broader and unambiguous approach covering tropical species. Biol. Rev. 2019, 94, 1161–1178. [Google Scholar] [CrossRef] [PubMed]
- Mattos, P.P.; Seitz, R.A.; Muniz, G.I.B. Growth rings based on periodical shoot growth. In Tree-Ring Analysis: Biological, Methodological and Environmental Aspects; Wimmer, R., Vetter, R.E., Eds.; CABI Publishing: New York, NY, USA, 1999; pp. 139–145. [Google Scholar]
- Dünisch, O.; Bauch, J.; Gasparotto, L. Formation of increment zones and intraannual growth dynamics in the xylem of Swietenia macrophylla, Carapa guianensis, and Cedrela odorata (Meliaceae). IAWA J. 2002, 23, 101–119. [Google Scholar] [CrossRef]
- Baker, J.C.; Santos, G.M.; Gloor, M.; Brienen, R.J. Does Cedrela always form annual rings? Testing periodicity across South America using radiocarbon dating. Trees 2017, 31, 1999–2009. [Google Scholar] [CrossRef] [Green Version]
- Artschwager, E. The time factor in the differentiation of secondary xylem and phloem in pecan. Am. J. Bot. 1950, 37, 15–24. [Google Scholar] [CrossRef]
- Derr, W.F.; Evert, R.F. The cambium and seasonal development of the phloem in Robinia pseudoacacia. Am. J. Bot. 1967, 54, 147–153. [Google Scholar] [CrossRef]
- Evert, R.F. The cambium and seasonal development of the phloem in Pyrus malus. Am. J. Bot. 1963, 50, 149–159. [Google Scholar] [CrossRef]
- Davis, J.D.; Evert, R.F. Seasonal development of the secondary phloem in Populus tremuloides. Bot. Gaz. 1968, 129, 1–8. [Google Scholar] [CrossRef]
- Alfieri, F.J.; Evert, R.F. Seasonal development of the secondary phloem in Pinus. Am. J. Bot. 1968, 55, 518–528. [Google Scholar] [CrossRef]
- Tucker, C.M.; Evert, R.F. Seasonal development of the secondary phloem in Acer negundo. Am. J. Bot. 1969, 56, 275–284. [Google Scholar] [CrossRef]
- Artschwager, E. Growth Studies on Guayule (Parthenium argentatum); US Department of Agriculture: Washington, DC, USA, 1945; p. 885. [Google Scholar]
- Ghouse, A.K.M.; Hashmi, S. Peridiocity of cambium and the formation of xylem and phloem in Mimusops elengi L., an evergreen member of tropical India. Flora 1983, 173, 479–487. [Google Scholar] [CrossRef]
- Gilbert, S.G. Evolutionary significance of ring porosity in woody angiosperms. Bot. Gaz. 1940, 102, 105–120. [Google Scholar] [CrossRef]
- Evert, R.F. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body—Their Structure, Function and Development, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; 601p. [Google Scholar]
- Hacke, U.G.; Spicer, R.; Schreider, S.G.; Plavková, L. An ecophysiological and developmental perspective on variation in vessel diameter. Plant Cell Environ. 2017, 40, 831–845. [Google Scholar] [CrossRef]
- Christmas, M.A.; Sperry, J.S.; Smith, D.D. Rare pits, large vessels and extreme vulnerability to cavitation in a ring-porous tree species. New Phytol. 2012, 193, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Govarts, R.; Frodin, D.G. World Checklist and Bibliography of Fagales (Betulaceae, Corylaceae, Fagaceae and Ticodendraceae); Royal Botanical Gardens, Kew: London, UK, 1998. [Google Scholar]
- Valencia, S.A. Diversidad del género Quercus (Fagaceae) en México. Bot. Sci. 2004, 75, 33–53. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Navarro, M.; Oyama, K.; García-Oliva, F.; Torres-Miranda, A.; Terrazas, T. Seasonal temperatura and precipitation regimes drive variation in the Wood of oak species (Quercus) along a climatic gradient in western Mexico. IAWA J. 2023, 44, 1–16. [Google Scholar] [CrossRef]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Bräker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol. Rev. 2003, 78, 119–148. [Google Scholar] [CrossRef] [Green Version]
- NOAA National Centers for Environmental Information. Climate at a Glance: City Time Series. Available online: https://www.ncdc.noaa.gov/cag/ (accessed on 25 May 2022).
- Sass, J.E. Botanical Microtechnique, 3rd ed.; Iowa State College Press: Ames, IA, USA, 1958. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw Hill: New York, NY, USA, 1940. [Google Scholar]
- Cheadle, V.I.; Gifford, E.M.; Esau, K. A staining combination for phloem and contiguous tissues. Stain. Technol. 1953, 28, 49–53. [Google Scholar] [CrossRef]
- Angyalossy, V.; Pace, M.R.; Evert, R.F.; Marcati, C.R.; Oskolski, A.A.; Terrazas, T.; Kotina, E.; Lens, F.; Mazzoni, S.C.; Angeles, G.; et al. IAWA list of microscopic bark features. IAWA J. 2016, 37, 517–615. [Google Scholar] [CrossRef] [Green Version]
- Lara, N.O.T.; Marcati, C.R. Cambial dormancy lasts 9 months in a tropical evergreen species. Trees 2016, 30, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Lara, N.O.T.; da Silva, M.R.; Nogueira, A.; Marcati, C.R. Duration of cambial activity is determined by water availability while cambial stimulus is day-length dependent in a Neotropical evergreen species. Environ. Exp. Bot. 2017, 141, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Angyalossy, V.; Pace, M.R.; Marcati, C.R.; Evert, R.F. Phloem development, growth markers, and sieve-tube longevity in two Neotropical trees. IAWA J. 2021, 42, 31–49. [Google Scholar] [CrossRef]
- Evert, R.F. Some aspects of phloem development in Tilia americana. Am. J. Bot. 1962, 49, 659. [Google Scholar]
- Imagawa, H.; Ishida, S. Study of the wood formation in trees. Report III. Occurrence of the overwintering cells in cambial zone in several ring-porous trees. Res. Bull. Col. Exp. For. Hokkaido. Univ. 1972, 29, 207–221. [Google Scholar]
- Evert, R.F. Phloem structure in Pyrus communis L. and its seasonal changes. Univ. Calif. Publ. Bot. 1960, 32, 127–194. [Google Scholar]
- Esau, K. Phloem structure in the grapevine, and its seasonal changes. Hildegardia 1948, 18, 217–295. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.D.; Evert, R.F. Cycle of phloem development in woody vines. Bot. Gaz. 1970, 131, 128–138. [Google Scholar] [CrossRef]
- Aloni, R. Vascular Differentiation and Plant Hormones; Springer: Cham, Switzerland, 2021; p. 339. [Google Scholar]
- Yañez-Espinosa, L.; Terrazas, T.; Lopez-Mata, L. Phenology and radial stem growth peridiocity in evergreen subtropical rainforest trees. IAWA J. 2010, 31, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.H. Xylem Structure and the Ascent of Sap; Springer: Berlin, Germany, 1983. [Google Scholar]
- Tyree, M.T.; Davis, S.D.; Cochard, H. Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA J. 1994, 15, 335–360. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.D.; Sperry, J.S.; Hacke, U.G. The relationship between xylem conduit diameter and cavitartion caused by freezing. Am. J. Bot. 1999, 10, 1367–1372. [Google Scholar] [CrossRef] [Green Version]
- Charrier, G.; Charra-Vaskou, K.; Kasuga, J.; Cochard, H.; Mayr, S.; Améglio, T. Freeze-thaw stress: Effects of temperature on hydraulic conductivity and ultrasonic activity in ten woody angiosperms. Plant Physiol. 2014, 164, 992–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcati, C.R.; Angyalossy, V.; Evert, R.F. Seasonal variation in wood formation of Cedrela fissilis (Meliaceae). IAWA J. 2006, 27, 199–211. [Google Scholar] [CrossRef]
- Lima, A.C. Sazonalidade da Atividade Cambial em Lianas. Master’s Thesis, Universidade de São Paulo, São Paulo, Brazil, 2012. [Google Scholar]
- Lens, F.; Gleason, S.M.; Bartolami, G.; Brodersen, C.; Delzon, S.; Jansen, S. Functional xylem characteristics associated with drought-induced embolism in angiosperms. New Phytol. 2022, 236, 2019–2036. [Google Scholar] [CrossRef] [PubMed]
- Carlquist, S. Ecological Strategies of Xylem Evolution; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Olson, M.; Pace, M.R.; Anfodillo, T. The vulnerability to drought induced embolism-conduit diameter link: Breaching the anatomy-physilogy divide. IAWA J. 2023, 44, 1–20. [Google Scholar] [CrossRef]
- Hacke, U.G.; Jacobsen, A.L.; Pratt, R.B. Vessel diameter and vulnerability to draught-induced embolism: Within-tissue and across-species patterns and the issue of survivorship bias. IAWA J. 2023, 44, 1–16. [Google Scholar] [CrossRef]
- Jacobsen, A.L.; Pratt, R.B. Vessel diameter polymorphism determines vulnerability-to-embolism curve shape. IAWA J. 2023, 44, 1–15. [Google Scholar] [CrossRef]
- González-González, B.D.; García-González, I.; Vázquez-Ruiz, R.A. Comparative cambial dynamics and phenology of Quercus robur L. and Q. pirenaica Willd. in an Atlantic forest of the northwestern Iberian Peninsula. Trees 2013, 27, 1571–1585. [Google Scholar] [CrossRef]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons; Clarendon Press: Oxford, UK, 1950; Volumes 1 and 2. [Google Scholar]
- Record, S.J.; Hess, R.W. Timbers of the New World; Arno Press: New York, NY, USA, 1972. [Google Scholar]
- Sousa, V.B.; Leal, S.; Quilhó, T.; Pereira, H. Characterization of cork oak (Quercus suber) wood anatomy. IAWA J. 2009, 30, 149–161. [Google Scholar] [CrossRef]
- Sousa, V.B.; Cardoso, S.; Pereira, H. Age trends in the wood anatomy of Quercus faginea. IAWA J. 2014, 35, 293–306. [Google Scholar] [CrossRef]
- Şen, A.U.; Quilhó, T.; Pereira, H. Bark anatomy of Quercus cerris L. var. cerris from Turkey. Turk. J. Bot. 2011, 35, 45–55. [Google Scholar] [CrossRef]
- Gričar, J.; Jagodic, Š.; Prislan, P. Structure and subsequent seasonal changes in the bark of sessile oaks (Quercus petraea). Trees 2015, 29, 747–757. [Google Scholar] [CrossRef]
- Balzano, A.; Čufar, K.; De Micco, V. Xylem and phloem formation dynamics in Quercus ilex L. at a dry site in southern Italy. Forests 2021, 12, 188. [Google Scholar] [CrossRef]
- Aronoff, S.; Dainty, J.; Gorham, J.; Srivastava, L.M.; Swanson, C.A. Phloem Transport; Plenum Press: New York, NY, USA, 1974. [Google Scholar]
- Evert, R.F. Dicotyledons. In Sieve-Elements: Comparative Structure, Induction and Development; Behnke, H.-D., Sjolund, R.D., Eds.; Springer: Berlin, Germany, 1990; pp. 103–137. [Google Scholar]
- Tucker, C.M. Seasonal phloem development in Ulmus americana. Am. J. Bot. 1968, 55, 716. [Google Scholar]
- Lamoureux, C.H. Phloem tissue in Angiosperms and Gymnosperms: Cell types and their spatial distribution; longevity of sieve elements; changes in old phloem. In Phloem Transport; Aronoff, S., Dainty, J., Gorham, J., Srivastava, L.M., Swanson, C.A., Eds.; Plenum Press: New York, NY, USA, 1974. [Google Scholar]
- Esau, K. The Phloem. Handbuch der Pflanzenanatomie; Band 5, Teil 2; Gebrüder Borntraeger: Berlin, Germany; Stuttgart, Germany, 1969. [Google Scholar]
- Holdheide, W. Anatomie mitteleuropäischer Gehölzrinden (mit mikrophotographischem Atlas). In Handbuch der Mikroskopie in der Technik; Band 5, Heft 1; Umschau Verlag: Frankfurt am Main, Germany, 1951; pp. 193–367. [Google Scholar]
- Wheeler, E.A.; Baas, P.; Manchester, S.R. Wood anatomy of modern and fossil Fagales in relation to phylogenetic hypotheses, familial classification, and patterns of character evolution. Int. J. Plant Sci. 2022, 183, 61–86. [Google Scholar] [CrossRef]
- Feuillat, F.; Dupouey, J.L.; Sciama, D.; Keller, R. A new attempt at discrimination between Quercus petraea and Quercus robur based on wood anatomy. Can. J. Forest Res. 1997, 27, 343–351. [Google Scholar] [CrossRef]
- Carlquist, S. Vasicentric tracheids as a drought survival mechanism in the woody flora of southern California and similar regions; review of vasicentric tracheids. Aliso 1985, 11, 37–68. [Google Scholar]
- Carlquist, S. How wood evolves: A new synthesis. Botany 2012, 90, 901–940. [Google Scholar] [CrossRef] [Green Version]
- Pratt, R.B.; Castro, V.; Jacobsen, A.L. The functional significance of tracheids co-occurring with vessels in xylem of Eudicots suggest role in embolism tolerance. IAWA J. 2023, 44, 1–18. [Google Scholar] [CrossRef]
- Ziemińska, K. The role of imperforate tracheary elements and narrow vessels in wood capacitance of angiosperm trees. IAWA J. 2023, 44, 1–14. [Google Scholar] [CrossRef]
- Barotto, A.J.; Fernandez, M.E.; Gyenge, J.; Meyra, A.; Martinez-Meier, A.; Monteoliva, S. First insight into the functional role of vasicentric tracheids and parenchyma in eucalyptus species with solitary vessels: Do they contribute to xylem efficiency or safety? Tree Physiol. 2016, 36, 1485–1497. [Google Scholar] [CrossRef] [Green Version]
- Whitmore, T.C. Studies in systematic bark morphology. IV. The bark of beech, oak and sweet chestnut. New Phytol. 1962, 62, 161–169. [Google Scholar] [CrossRef]
- Pereira, H.; Tavares, F.; Sousa, V.; Quilhó, T. Bark anatomy and cell size variation in Quercus faginea. Turk. J. Bot. 2013, 37, 561–570. [Google Scholar]
- Trockenbrodt, M. Qualitative structural changes during bark development in Quercus ruber, Ulmus glabra, Populus tremula and Betula pendula. IAWA Bull. 1991, 12, 5–22. [Google Scholar] [CrossRef]
- Pereira, H. Cork: Biology, Production and Uses; Elsevier: Amsterdam, The Netherlands, 2007; 160p. [Google Scholar]
- Sousa, V.; Ferreira, J.P.A.; Miranda, I.; Quilhó, T.; Pereira, H. Quercus rotundifolia bark as a source of polar extracts: Structural and chemical characterization. Forests 2021, 12, 1160. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pace, M.R.; Dutra, R.; Marcati, C.R.; Angyalossy, V.; Evert, R.F. Seasonal Cambial Activity and Formation of Secondary Phloem and Xylem in White Oaks (Quercus alba L.). Forests 2023, 14, 920. https://doi.org/10.3390/f14050920
Pace MR, Dutra R, Marcati CR, Angyalossy V, Evert RF. Seasonal Cambial Activity and Formation of Secondary Phloem and Xylem in White Oaks (Quercus alba L.). Forests. 2023; 14(5):920. https://doi.org/10.3390/f14050920
Chicago/Turabian StylePace, Marcelo R., Rafaella Dutra, Carmen R. Marcati, Veronica Angyalossy, and Ray F. Evert. 2023. "Seasonal Cambial Activity and Formation of Secondary Phloem and Xylem in White Oaks (Quercus alba L.)" Forests 14, no. 5: 920. https://doi.org/10.3390/f14050920