1. Introduction
Despite the fact that spring forest geophytes live together in bulk numbers and often create homogeneous stands, previous research has been implemented on individual or infraindividual levels in laboratories (e.g., [
1]), in situ (e.g., [
2]) or, in some cases, using both approaches (e.g., [
3,
4,
5]).
Spring ephemerals of temperate deciduous forests appear shortly after the snow has melted and senesce shortly after the overstory canopy has closed [
6]. During this period, geophytes complete their aboveground growth, which includes fruit production, within 2 months [
7]. Their leaves senesce when the light intensity decreases. Their photosynthetic response is mostly determined by adaptations to seasonal changes in light availability related to the dynamics of the overstory canopy [
8].
Spring ephemerals have high photosynthetic rates that allow them to rapidly accumulate carbohydrates and complete their aboveground growth in a few weeks [
7]. To sustain high photosynthetic activities in early spring, the plants must be able to absorb water efficiently at low soil temperatures and to allocate large amounts of nutrients to the shoot to compensate for lower enzymatic activity at low temperatures [
7].
During rapid shoot growth and fruiting, the bulbs and aboveground organs appeared to be competing sinks for the photosynthetically fixed carbon [
1]. A comparative study [
9] examined the photosynthetic induction dynamics of four deciduous forest understory herbs to sunflecks. It was discovered that early spring herbs responded as full-sun plants with a higher light compensation point and greater net photosynthesis at light saturation. The Photosynthetic Photon Flux Density (PPFD) required to maintain an induced photosynthetic state was significantly greater in spring-green herbs than in summer-green herbs. In field populations of geophytes, as the overstory leaf area index increased, declines occurred in all gas exchange characteristics except in apparent quantum efficiency [
4]. Spring forest geophytes showed high resorption efficiency during leaf senescence [
7].
The ecophysiological traits of seven spring forest geophytes, including the early flowering spring ephemerals
Ranunculus ficaria L. and
Corydalis cava (L.) Koerte were compared by Popović et al. [
2]. It was observed that all species showed similar seasonal dynamics of the assessed parameters, but the average seasonal values of photosynthesis and the light compensation point differed significantly. Another comparative study [
3] showed that spring forest geophytes had the lowest N uptake capacity in both laboratory and field experiments. However, they maintained substantial root uptake capacity throughout the summer when they showed no photosynthetic activity, and the mean residence time of N was also significant in these plants.
A
13CO
2-trace experiment focused on the translocation of current photosynthetic products to individual organs [
10]. It showed that fruit-forming plants retained leaves longer than vegetative plants and foliar photosynthetic products were largely transported to bulbs. Additionally, the bract-clipping significantly reduced seed production; therefore, the current photosynthesis of leafy bracts might be a major carbon source for fruit development.
Some parts of the research have been manipulated in order to examine, e.g., the responses of leafing phenology and photosynthesis to soil warming in seven understory species with various leaf habits [
11]. The soil temperature in the warming plots was electrically maintained 5 °C higher than in the control plots. Based on this experience, it can be said that physiological responses to soil warming were highly species-specific, but the soil warming impacts on understory plants were most prominent in spring, especially for phenological characteristics. Another manipulated examination focused on the effects of air and soil temperature during the growth of spring ephemerals [
6]. The results showed that the final dry mass was higher at the lower temperature regime, while leaves lasted longer than at the higher temperature regime. In addition, both air and soil temperatures had an impact on the growth of forest geophytes.
The phenological and physiological responses of plants to climate change are key issues for understanding the global warming impact on ecosystems [
11]. Thus, a better understanding of the factors controlling the growth rate of spring ephemerals is needed before we can predict these plants’ response to climate changes [
7].
The objectives of this study were (i) to measure the Net Ecosystem CO2 Exchange (NEE) and evapotranspiration (ET) fluxes of forest geophytes during their vegetation period; (ii) to compare leaf and stand level net photosynthesis; (iii) to examine the dependence of NEE on light, air temperature (Tair) and leaf area (LA) as well as the connection between NEE and ET.
2. Materials and Methods
2.1. Study Site
The examinations were carried out in a 1.5 ha oak forest patch of Szent István University Gödöllő Botanical Garden (Gödöllő 19°14′ E, 47°25′ N, 250 m elevation) [
12]. The surface of the study site is flat, so the topographical conditions did not influence the results. Three different ground plant communities or stands were selected within a 100-square-metre area. The investigated stands were at the same distance from the edge of the forest and a few metres from each other, thereby ensuring the greatest possible similarity of abiotic factors. The examined communities were dominated by three spring forest geophytes widely common in Europe [
5,
13] boasting three different subterranean storage organs:
Anemone ranunculoides L. had rhizome,
Corydalis solida (L.) Clairv. was tuberous and
Ranunculus ficaria L. had swollen, club-like root-tubers. The geophytes dominated in the patches (d = 60 cm, A = 0.2826 m
2) of the stands selected for examination. The abundance of the selected geophyte species in the studied patches was representative of the entire stand. Their abundance was also indicated by the fact that their proportion in the ground canopy and the aboveground phytomass was at least 90 percent within the examined patches during the entire study period. All selected geophyte stands bordered other geophyte stands in the forest.
Mean annual temperature of the Botanical Garden is 10.5 °C; the garden’s average amount of precipitation is 587 mm [
14]. The vegetation period lasts from March to mid-November. The coolest month is January with an average temperature of 1 °C, while the warmest month is July with an average temperature of 22 °C. The driest month is January with an average monthly rainfall of less than 20 mm.
Taxon nomenclature followed the Euro + Med PlantBase [
15].
2.2. Measurements and Data Collection
NEE of patches of geophyte vegetation was carried out episodically on a weekly basis during the whole vegetation period using chamber technique operating in open system and Ciras 2 infrared gas analyser (PP Systems, Hitchin, UK). The transparent, non-destructive, portable plexiglass chamber was circle-shaped with a diameter of 60 cm. Previously, we had successfully examined different grassland types by using the same technique (e.g., [
16,
17]). NEE measurements of each stand were implemented between 11 am and 2 pm within the three selected permanent patches. The exact location was identified by a circular metal plate (d = 60 cm) sunk into the soil. Measurements were conducted between 30 Mar and 18 May, between 30 March and 4 May and between 23 March and 12 May in the cases of
Anemone ranunculoides,
Corydalis solida and
Ranunculus ficaria, respectively. The infrared gas analyser (IRGA) measured the difference in CO
2 flux (CO
2 concentrations of both the reference and analysed air at the inbound and outbound pipes of the chamber) and the ET of the stand covered by the chamber (ET was determined by measuring the differences in water vapour concentrations of reference /in/ and analysed /out/ air), while PPFD and T
air were measured by the IRGA during the gas exchange measurements. The gas analyser examined six samples per minute for an average of 60 min for each patch of geophyte vegetation.
Average temperatures of the given stand were used for NEE calculations. The following differential equation was applied when calculating CO
2 flux:
where ‘
F’ is the stand level flux of (NEE)/CO
2 uptake of the plant stand, vegetation and soil respiration in the case of CO
2 flux or the resultant of the ET from the soil surface covered by vegetation in the case of ET/ (mol m
−2 s
−1); ‘Δ
c’ represents the difference in incoming and outgoing concentrations of /CO
2 or H
2O/ (mol m
−3); ‘
Q’ is the flow rate (m
3 s
−1); and ‘
A’ is the soil surface covered by the chamber (m
2).
In the subsequent quality control, raw data was manually filtered where negative ET values and outliers that were recorded at the initial and final phases of the measurement (due to human respiration) had not been taken into account.
Leaf photosynthesis (on 6 replicates from 6 different plants) of the dominant species was measured twice during intensive root–shoot allocation (13 April) and reallocation (4 May) periods. The standard cuvette of the same infrared gas analyser (PP Systems, Hitchin, UK) and permanent plots were used for leaf photosynthesis measurements in order to compare the infraindividual (or leaf level) and stand level net photosynthetic rate.
As for estimating the LA of the stands, all leaves of the plants were counted in each permanent patch and each measuring occasion. Samples were collected (at least three samples/species) from outside the examined area in order to avoid the disturbance of the patches. These samples were selected with the consideration that they should match the current phenological phase and the average leaf size of the measured stand. The exact leaf area was determined by using graph paper and the average value was multiplied by the number of leaves counted within the examined patches.
Micrometeorological measurements and LA estimations were used to determine the dependence of the measured NEE values on some abiotic (PPFD and Tair) and biotic (LA) factors. HOBO (MicroStation, Onset, MA, USA) micrometeorological station was installed among the selected geophyte stands. The sensors of this station sampled the temperature and moisture of soil (both at 5 cm depth) as well as PPFD in 5 min intervals during the entire vegetation period of the ephemerals and submitted these data to the automatic data recording unit.
Current phenological phases of geophytes including the number of blooming specimens were constantly recorded together with the results of physiological measurements.
2.3. Statistical Analyses
The Kolmogorov–Smirnov test was performed to test for normal distribution of the data. For testing the homoscedasticity, Bartlett’s test was used. For normally distributed data, Student’s t-test and one-way analysis of variance (ANOVA) was applied to identify significant differences between datasets, while, for non-normally distributed data, the nonparametric Mann–Whitney test was performed instead at significance level of 0.05. Linear regression and non-linear regression were calculated using SigmaPlot2012 (SPSS Inc., Chicago, IL, USA) in order to evaluate the influence of PPFD, Tair, LA and ET on NEE. To estimate the unknown parameters, curve fitting was established by minimizing the sum of the squared residuals (method of ordinary last squares—OLS). Coefficient of determination (r2) was used to explain how well the variation from the continuous explanatory variable could be predicted. Only significant regressions were represented in the figures indicating the strength of the regression. The most significant regression was fitted to the data in all cases. Statistical analyses were calculated using SigmaPlot2012. Figures were created using the same software.
3. Results
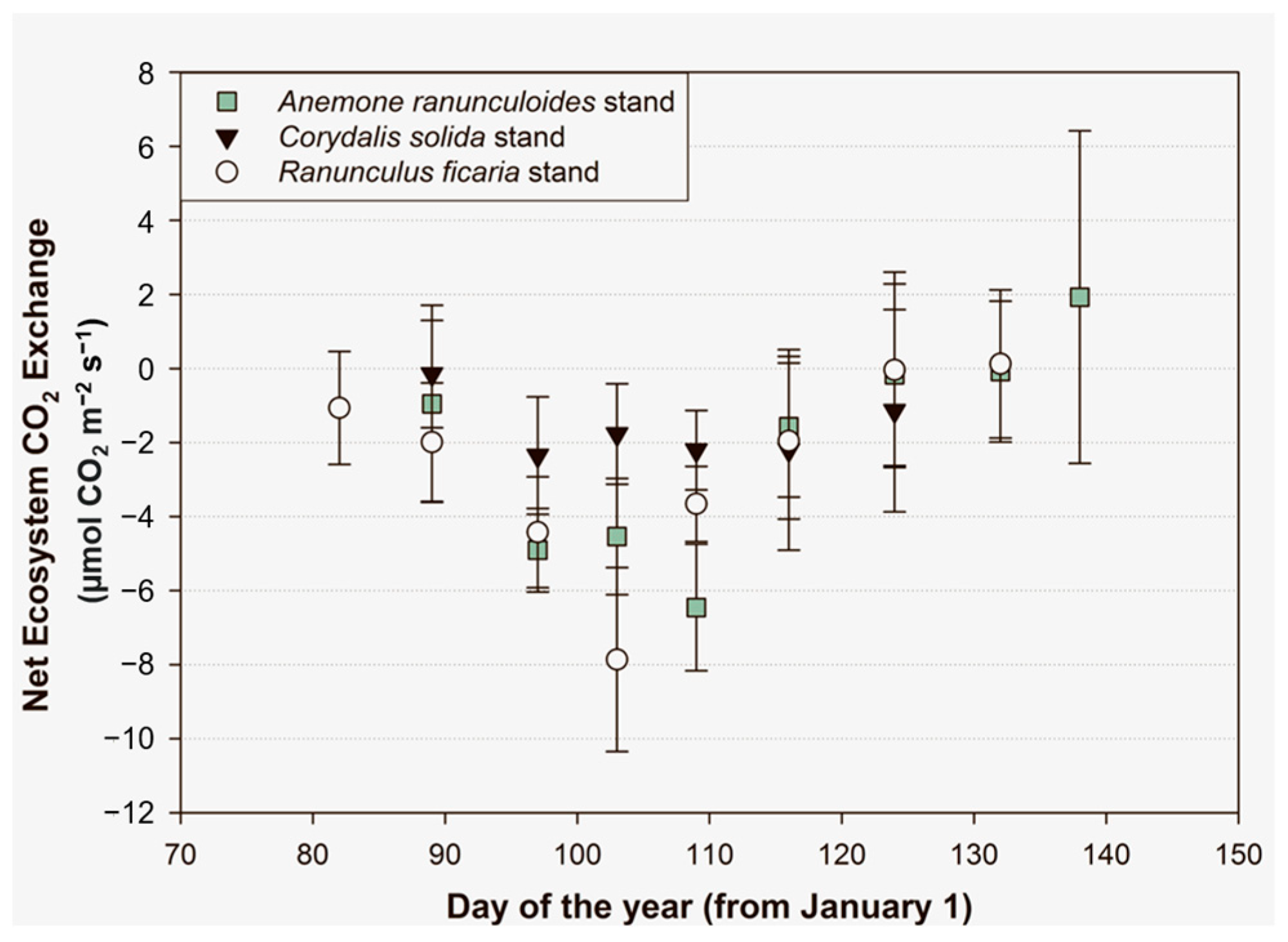
At stand level, all three species were carbon sinks from the beginning of the vegetation period until the end of it (
Corydalis solida) or until the last phase of fruit formation (
Anemone ranunculoides and
Ranunculus ficaria) (
Figure 1). The largest amount of carbon sink was measured in the
R. ficaria stand four weeks after the appearance of the aboveground shoots. CO
2 emission was detected in the cases of
R. ficaria and
A. ranunculoides but only in the last week of their aboveground lifecycle. The extent of carbon sink activity increased rapidly and constantly during the first four weeks in the
R. ficaria patch. A similar trend could be observed within the
A. ranunculoides stand while the carbon fixation of the patch dominated by
C. solida was smaller but more balanced. Apart from the first and the last weeks of the experiment, the
C. solida stand showed a nearly constant NEE value. Based on the cumulated averages, the carbon sink was the largest in the
R. ficaria stand: it was 25% higher than in
A. ranunculoides and more than double than that of
C. solida. It can be noted in all three groups that the standard deviation of NEE was higher at the beginning and at the end of the geophytes’ aboveground lifecycle but lower during the in-between period, including during the blooming phenophase. All three geophytes completed their aboveground lifecycle—including fruit production—within 8 weeks. This correlates to the lifecycle length of North American spring forest geophytes [
7].
C. solida had the shortest lifecycle of only 6 weeks. From the end of March until the beginning of May, it finished fruit production and its aboveground phytomass had disappeared. On the contrary, the other two plants had a lifecycle of 8 weeks.
R. ficaria appeared in the second half of March and its aboveground parts were last observable and measurable in mid-May. The lifecycle of
A. ranunculoides followed that of
R. ficaria with a one-week shift (
Figure 1).
Leaf photosynthesis exceeded NEE fluxes at all sites and during the measuring period (
Figure 2), with the exception of the
Corydalis solida stand during leaf senescence (
Figure 2b). During intensive leaf development (
Figure 2a), the differences were significant for
Anemone (
p < 0.001),
Corydalis (
p < 0.001) and
Ranunculus (
p < 0.01).
Regarding stand level carbon uptake and PPFD, a positive correlation was observed for all three species. The strongest linear relationship (
r2 = 0.84) was found in the case of
A. ranunculoides (
Figure 3), which proved to be significant (
F = 32.15,
p < 0.01). The highest NEE value was measured at the first half of the vegetation cycle when PPFD was also the largest because shrub layer and canopy had not come into leaf yet.
To the contrary of radiation, a weak negative correlation was found between NEE and T
air in all three stands. As spring advanced and T
air increased, the storage organs of ephemerals were filled, and, thus, their carbon sequestration [
4] and storage capacity decreased considerably. As for the
A. ranunculoides stand (
Figure 4), the measurement made in mid-April reduced the negative correlation. In this case, at the peak of blooming, a high T
air corresponded to the largest amount of carbon sequestration. Nevertheless, the relationship was still the strongest at this type.
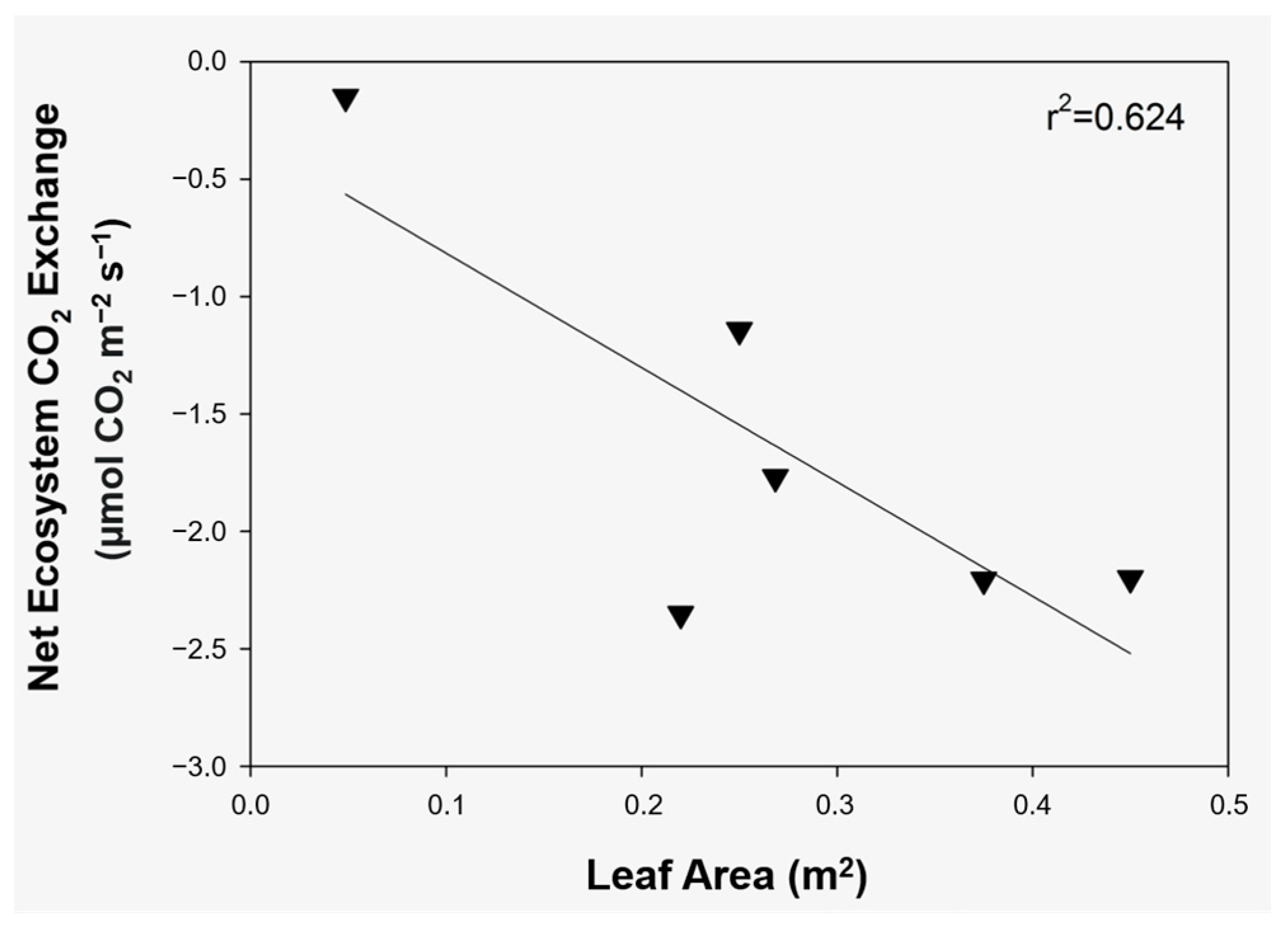
In all stands, a positive correlation was observed between the stand level CO
2 fixation and LA, and, thus, the intensive carbon sink was measured only when the leaf area considerably increased. A significant (
F = 25.54,
p < 0.01) linear correlation and a strong, but not significant, linear correlation provided the strongest connections in
A. ranunculoides (
Figure 5) and
C. solida (
Figure 6), respectively.
The average NEE positive correlation was measured in all three types between stand level carbon uptake and ET. It was the strongest in the
R. ficaria stand in which a significant (
F = 24.51,
p < 0.01) linear relationship (
r2 = 0.84) was observed (
Figure 7) between NEE and ET. The linear relationship for the patch of
A. ranunculoides was also strong, but no significant correlation was revealed regarding the averages of CO
2 sink and ET. However, if the data of the sampling days was assessed separately, then the correlation was revealed to be significant, but a positive correlation cannot exclusively be observed in the seasonal dynamics of NEE and ET (
Figure 8). During the first three weeks a weak, and insignificant, but a positive correlation was found between the stand level CO
2 uptake and ET (
Figure 8a–c). In the fourth week, at the time of full foliage development and the peak of blooming, this positive and linear relationship (
r2 = 0.35) reached its highest level and became significant (
F = 146.21,
p < 0.001) (
Figure 8d). A week later, a similarly significant (
F = 57.28,
p < 0.001) and a linear but weak negative regression (
r2 = 0.11) was observed (
Figure 8e). During the last three weeks, an exponential rise to the max type negative curve gave the strongest significant (
p < 0.001) regression between NEE and ET (
Figure 8f /
F = 45.56/,
Figure 8g /
F = 22.80/,
Figure 8h /
F = 22.04/). From the fourth week until the end of the vegetation period, strong and significant (
p < 0.001) regressions were found between these two variables (
Figure 8d–h).
5. Conclusions
Due to the chamber measurement technique used, we could, for the first time, obtain some information on the carbon uptake magnitude of spring forest geophytes. CO
2 exchange measurements proved that the phenological stage of the geophytes do affect their carbon fluxes. During intensive leaf development, a significant difference was observed between leaf and stand level net photosynthesis. Spring forest geophytes maintain their remarkable carbon uptake potential during almost their entire aboveground phenophase. Despite their short overground lifecycle, the remarkable carbon uptake capacity indicates the non-negligible role of geophyte vegetation in the carbon flux of temperate forests. Out of the environmental variables, PPFD and LA strongly influenced the NEE, while the role of T
air was negligible. More stands expressed the dependence of NEE on ET (
Figure 7 and
Figure 8), PPFD (
Figure 3) and T
air (
Figure 4), which provided new proof about the role of stand level operation, stability and regulation during periods when plants have larger leaf areas. This regulation proves that there is no correlation in the initial phase of stand development; rather, it builds up gradually. Moreover, in the case of grounds covered with vegetation, the extent of transpiration is determined, in addition to the main abiotic factors (PPFD, T
air), by the biotic factor: the vegetation and, mainly, the foliage and root system. In conclusion, ET is less dependent on abiotic factors in the case of a larger LA because ET is considerably regulated by the vegetation. Furthermore, the light and temperature conditions at ground level fluctuate in a smaller range compared to bare [
26] or partly covered soils. The results of our examinations can be used for modelling plant stands with similarly rapid dynamics (e.g., weed communities).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







