
Distribution and Prolonged Diapause of the Rowan Seed Predators Argyresthia conjugella (Lepidoptera: Yponomeutidae) and Megastigmus brevicaudis (Hymenoptera: Torymidae) and their Parasitoids in Norway
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Rowanberries
2.2. Rearing of Insects for the Mapping
2.3. Study of Prolonged Diapause
2.4. Identification of Emerged Imagines
3. Results
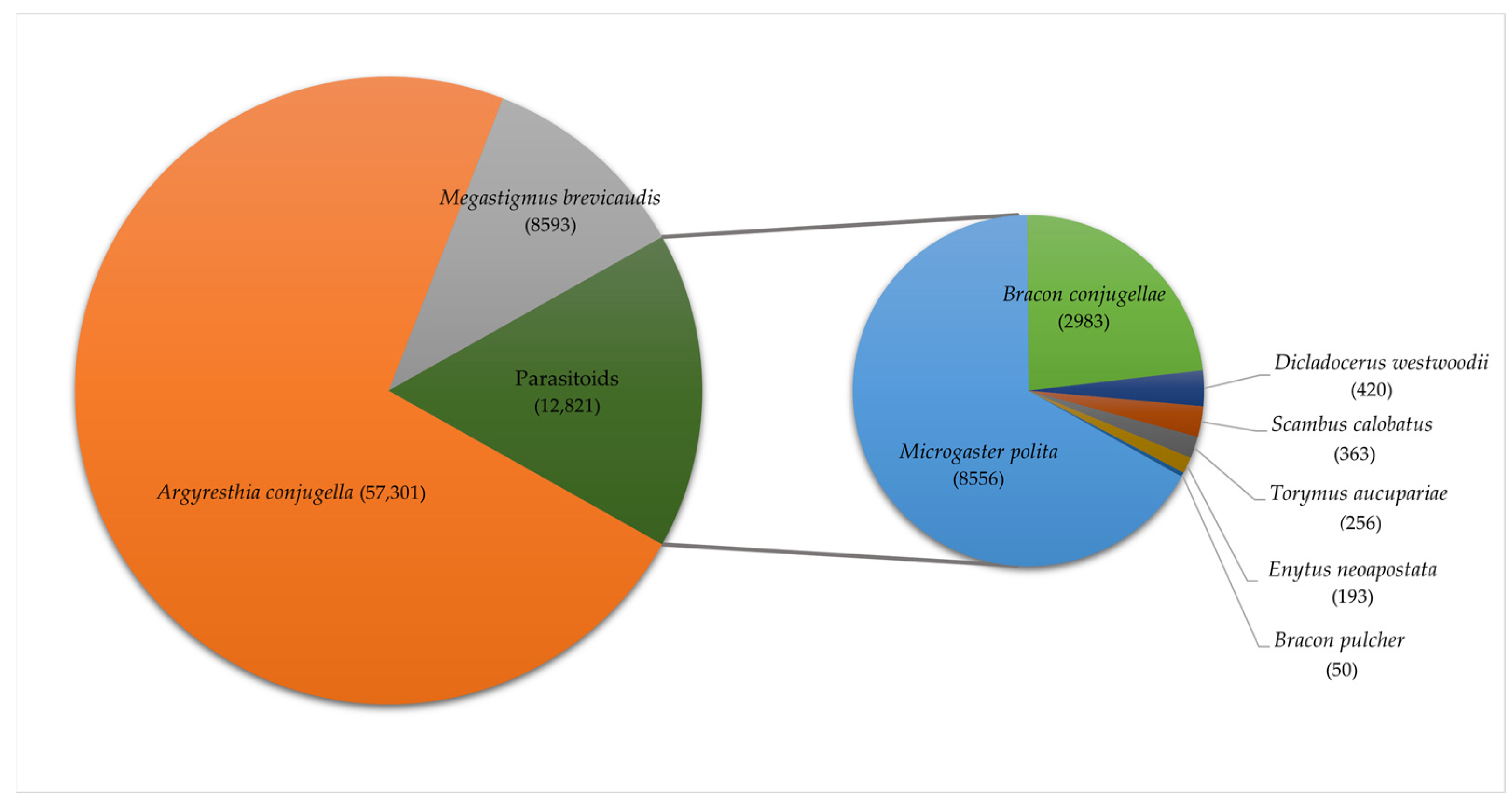
3.1. The Species Found and Their Geographical Distribution
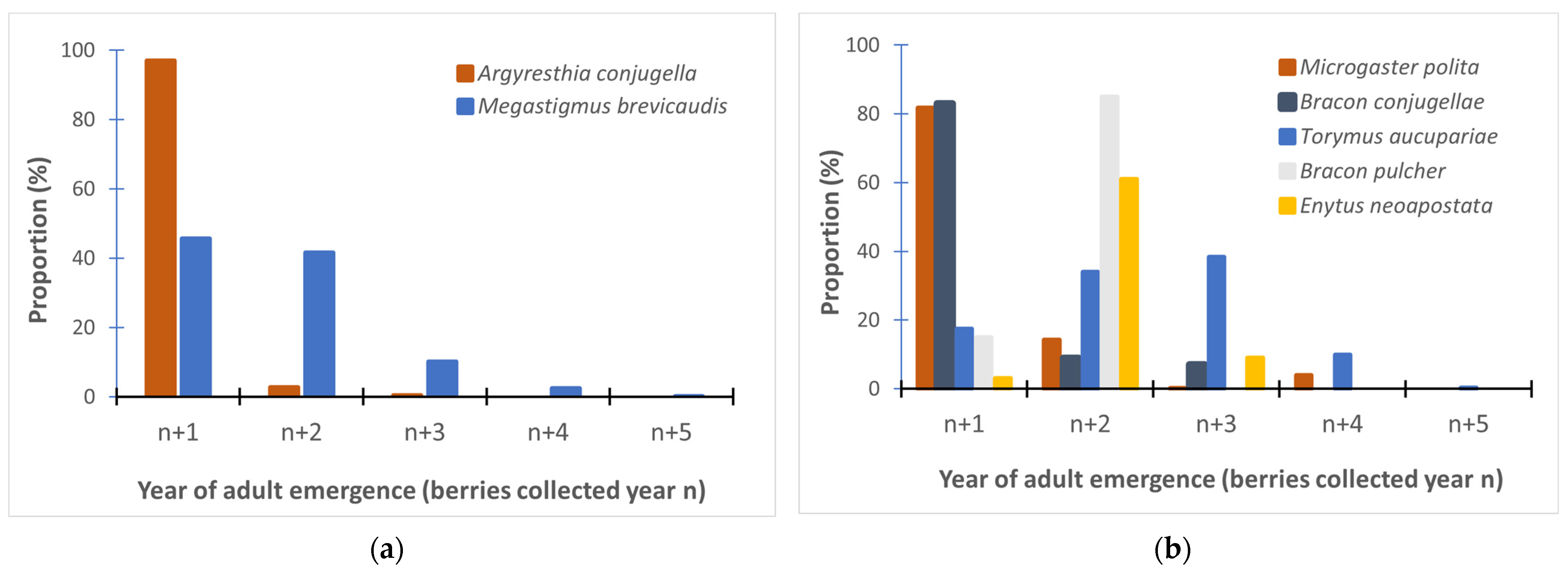
3.2. Prolonged Diapause
4. Discussion
4.1. Geographical Distribution of the Seed Predators
4.2. Biology and Distribution of the Parasitoids
4.3. Prolonged Diapause
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Räty, M.; Caudullo, G.; de Rigo, D. Sorbus aucuparia in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 176–177. Available online: https://www.researchgate.net/publication/299471313_Sorbus_aucuparia_in_Europe_distribution_habitat_usage_and_threats (accessed on 20 January 2023).
- Raspé, O.; Findlay, C.; Jaquemart, A.-L. Sorbus aucuparia L. Biological flora of the British Isles. J. Ecol. 2000, 88, 910–930. [Google Scholar] [CrossRef]
- Koenig, W.D. A brief history of masting research. Phil. Trans. R. Soc. B 2021, 376, 20200423. [Google Scholar] [CrossRef] [PubMed]
- Kobro, S.; Søreide, L.; Djønne, E.; Rafoss, T.; Jaastad, G.; Witzgall, P. Masting of rowan Sorbus aucuparia L. and consequences for the apple fruit moth Argyresthia conjugella Zeller. Popul. Ecol. 2003, 45, 25–30. [Google Scholar] [CrossRef]
- Ahlberg, O. Rönnbärsmalen, Argyresthia conjugella Zell. En redogörelse för undersökningar åren 1921–1926. In Meddelande från Centralanstalten för Försöksväsendet på Jordbruksområdet; Lantbruksentomologiska Avdelingen: Stockholm, Sweden, 1927; Volume 324, pp. 1–127, (In Swedish with English Summary). [Google Scholar]
- Jespersen, C.; Lomholt, O. De danske Frøvepse, Megastigmus Dalman, 1820 (Hymenoptera, Chalcidoidea, Torymidae). Ent. Meddr. 1983, 50, 111–118. [Google Scholar]
- Sperens, U. Fruit production in Sorbus aucuparia L. (Rosaceae) and pre-dispersal seed predation by the apple fruit moth (Argyresthia conjugella Zell.). Oecologia 1997, 110, 368–373. [Google Scholar] [CrossRef]
- Milliron, H.E. Taxonomic and biological investigations in the genus Megastigmus with particular reference to the taxonomy of the Nearctic species (Hymenoptera: Chalcidoidea; Callimomidae). Am. Midl. Nat. 1949, 41, 257–420. [Google Scholar] [CrossRef]
- Roques, A.; Skrzypczyńska, M. Seed-infesting chalcids of the genus Megastigmus Dalman, 1820 (Hymenoptera: Torymidae) native and introduced to the West Palearctic region: Taxonomy, host specificity and distribution. J. Nat. Hist. 2003, 37, 127–238. [Google Scholar] [CrossRef]
- Rouault, G.; Turgeon, J.; Candau, J.-N.; Roques, A.; Aderkas, P.v. Oviposition strategies of conifer seed chalcids in relation to host phenology. Naturwissenschaften 2004, 91, 472–480. [Google Scholar] [CrossRef]
- Hanski, I. Four kinds of extra long diapause in insects: A review of theory and observations. Ann. Zool. Fenn. 1988, 25, 37–53. [Google Scholar]
- Danks, H.V. Insect Dormancy: An Ecological Perspective; Biological Survey of Canada (Terrestrial Arthropods), National Museum of Natural Sciences: Ottawa, ON, Canada, 1987; pp. 175–187. [Google Scholar]
- Bakke, A. Distribution of prolonged diapausing larvae in populations of Laspeyresia strobilella L. (Lep., Tortricidae) from spruce cones. Norsk Entomol. Tidsskr. 1971, 18, 89–93. [Google Scholar]
- Annila, E. Fluctuations in cone and seed insect populations in Norway spruce. In Communicationes Instituti Forestalis Fenniae (Finland); Metsäntutkimuslaitos, 1981; Volume 101, pp. 1–32, (In Finnish with English Summary). [Google Scholar]
- Turgeon, J.J.; Roques, A.; Groot, P. Insect fauna of coniferous seed cones: Diversity, host plant interactions, and management. Annu. Rev. Entomol. 1994, 39, 179–212. [Google Scholar] [CrossRef]
- Krämer, K. Zur Biologie der Ebereschenmotte (Argyresthia conjugella Zell.). Anz. Für Schädlingskunde 1960, 33, 102–107. [Google Scholar] [CrossRef]
- Edland, T. Apple fruit moth (Argyresthia conjugella Zell.): Large attacks on apples are to be expected in 1971. Gartneryrket 1971, 61, 444–445. (In Norwegian) [Google Scholar]
- Corley, J.C.; Capurro, A.F.; Bernstein, C. Prolonged diapause and the stability of host–parasitoid interactions. Theor. Popul. Biol. 2004, 65, 193–203. [Google Scholar] [CrossRef]
- Satake, A.; Bjørnstad, O.N.; Kobro, S. Masting and trophic cascades: Interplay between rowan trees, apple fruit moth, and their parasitoid in southern Norway. Oikos 2004, 104, 540–550. [Google Scholar] [CrossRef]
- Le Lann, C.; van Baaren, J.; Visser, B. Dealing with predictable and unpredictable temperatures in a climate change context: The case of parasitoids and their hosts. J. Exp. Biol. 2021, 224, jeb238626. [Google Scholar] [CrossRef]
- Schøyen, W.M. Beretning om Skadeinsekter og Plantesygdomme i 1898; Grøndahl & Søns Bogtrykkeri: Kristiania, Norway, 1899; pp. 18–19. (In Norwegian) [Google Scholar]
- Edland, T. Apple fruit moth (Argyresthia conjugella Zell.): Forecast of risk for attacks on apple. Methods and results. Gartneryrket 1974, 64, 524–533, (In Norwegian, English Summary). [Google Scholar]
- Edland, T. Benefits of minimum pesticide use in insect and mite control in orchards. In Techniques for Reducing Pesticide Use. Economic and Environmental Benefits; Pimentel, D., Ed.; Wiley & Sons: New York, NY, USA, 1997; pp. 197–220. [Google Scholar]
- Vips Platform. Available online: https://www.vips-landbruk.no/ (accessed on 24 February 2023).
- Skog, T.-E.; Nordskog, B.; Eikemo, H.; Hole, H.; Schjøll, A.F.; Netland, J.; Trandem, N.; Rafoss, T. An open source technology platform for international collaboration and local adaptation of IPM tools. In Book of Abstract; IOBC VIII Workshop on Integrated Soft Fruit Production: Trentino, Italy, 2014; p. 24. [Google Scholar]
- Hogstad, O.; Selås, V.; Kobro, S. Explaining annual fluctuations in breeding density of fieldfares Turdus pilaris—Combined influences of factors operating during breeding, migration and wintering. J. Avian Biol. 2003, 34, 350–354. [Google Scholar] [CrossRef]
- Fox, A.D.; Kobro, S.; Lehikoinen, A.; Lyngs, P.; Väisänen, R.A. Northern Bullfinch Pyrrhula p. pyrrhula irruptive behaviour linked to rowanberry Sorbus aucuparia abundance. Ornis Fenn. 2009, 86, 51–60. [Google Scholar]
- Økland, J. Utbredelse av noen ferskvannsmuslinger i Norge, og litt om European Invertebrate Survey. Fauna 1976, 29, 29–40. [Google Scholar]
- Lampa, S. Rönnbärsmalen (Argyresthia conjugella Zell.). Entomol. Tidskr. 1906, 27, 1–13. Available online: https://archive.org/details/entomologisktids2728190607ento/page/n21/mode/2up (accessed on 1 February 2023).
- Liston, A.; Prous, M.; Vårdal, H. A review of west palaearctic Hoplocampa species, focussing on Sweden (Hymenoptera, Tenthredinidae). Zootaxa 2019, 4615, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Thuróczy, C.K.; Hansen, L.O. Systasis edlandi—A new species of Ormocerinae (Hymenoptera, Pteromalidae) from Norway associated with rowan berries (Sorbus aucuparia) (Rosaceae). Norw. J. Entomol. 2015, 62, 205–209. Available online: http://urn.nb.no/URN:NBN:no-52321 (accessed on 27 February 2023).
- Hawkins, B.A.; Askew, R.R.; Shaw, M.R. Influences of host feeding-niche and foodplant type on generalist and specialist parasitoids. Ecol. Entomol. 1990, 15, 275–280. [Google Scholar] [CrossRef]
- Althoff, D.M. Does parasitoid attack strategy influence host specificity? A test with New World braconids. Ecol. Entomol. 2003, 28, 500–502. [Google Scholar] [CrossRef]
- Yu, D.S.; van Achterberg, K.; Horstmann, K. World Ichneumonoidea 2004. Taxonomy, Biology, Morphology and Distribution. Taxapad 2005. DVD Version.
- Kopelke, J.P.; Nymann, T.; Cazelles, K.; Gravel, D.; Vissault, S.; Roslin, T. Food-web structure of willow-galling sawflies and their natural enemies across Europe. Ecology 2017, 98, 1730. Available online: http://onlinelibrary.wiley.com/doi/10.1002/ecy.1832/suppinfo (accessed on 1 March 2023). [CrossRef]
- Shaw, M.R.; Huddleston, T. Classification and biology of Braconid wasps (Hymenoptera: Braconidae). In Handbooks for the Identification of British Insects; Royal Entomological Society: London, UK, 1991; Volume 7, Part 11; pp. 54–58. [Google Scholar]
- Noyes, J.S.; Universal Chalcidoidea Database. World Wide Web Electronic Publication 2019. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 1 February 2023).
- Askew, R.R. Hymenoptera. 2. Chalcidoidea section b. In Handbooks for the Identification of British Insects; Royal Entomological Society: London, UK, 1968; Volume VIII, p. 18. Available online: https://www.royensoc.co.uk/wp–content/uploads/2022/01/Vol08_Part02b.pdf (accessed on 23 January 2023).
- Horstmann, K. Typenrevision der europäischen Arten der Gattung Diadegma Foerster (syn. Angitia Holmgren) (Hymenoptera: Ichneumonidae). Beitr. Ent. Bd. 1969, 19, 413–472. [Google Scholar] [CrossRef]
- Broad, G.R.; Shaw, M.R.; Fitton, M.G. Ichneumonid wasps (Hymenoptera: Ichneumonidae): Their classification and biology. In Handbook for the Identification of British Insects; Field Studies Council: London, UK, 2018; Volume 7, Part 12; pp. 146, 271. [Google Scholar]
- Shaw, M.R. Notes on some European Microgastrinae (Hymenoptera: Braconidae) in the National Museums of Scotland, with twenty species new to Britain, new host data, taxonomic changes and remarks, and descriptions of two new species of Microgaster Latreille. Entomol. Gaz. 2012, 63, 173–201. [Google Scholar]
- Gauld, I.; Bolton, B. The Hymenoptera; Oxford University Press: Oxford, UK, 1988; p. 217. [Google Scholar]
- Shaw, M.R.; Jennings, M.T.; Quicke, D.L.J. The identity of Scambus planatus (Hartig, 1838) and Scambus ventricosus (Tschek, 1871) as seasonal forms of Scambus calobatus (Gravenhorst, 1829) in Europe (Hymenoptera, Ichneumonidae, Pimplinae, Ephialtini). J. Hymenopt Res. 2011, 23, 55–64. [Google Scholar] [CrossRef]
- Scaramozzino, P.L.; Di Giovanni, F.; Loni, A.; Ricciardi, R.; Lucchi, A. Updated list of the insect parasitoids (Insecta, Hymenoptera) associated with Lobesia botrana (Denis & Schiffermüller, 1775) (Lepidoptera, Tortricidae) in Italy. 2. Hymenoptera, Ichneumonidae, Anomaloninae and Campopleginae. ZooKeys 2018, 772, 47–95. [Google Scholar] [CrossRef]
- Ryan, R.B.; Denton, R.E. Initial Release of Chrysocharis laricinellae and Dicladocerus Westwoodii for Biological Control of the Larch Casebearer in the Western United States; US Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1973; Volume 200, pp. 1–4. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201303127920 (accessed on 1 February 2023).
- Furenhed, S. Ground-Living Predators of the Apple Fruit Moth Argyresthia conjugella (Zell.). Master’s Thesis, Sveriges Lantbruksuniversitet, Uppsala, Sweden, 2006. [Google Scholar]
- Zilsparne, A. Phenologische Daten über der Argyresthia conjugella L. und auswendung von Trichogramma bei dessen bekämpfung im Jahre 1971. Zool. Muz. Raksti 1973, 11, 49–54. [Google Scholar]
- Boivin, T.; Auger-Rozenberg, M.-A. Native fruit, cone and seed insects in the Mediterranean basin. In Insects and Diseases of Mediterranean Forest Systems; Paine, T.D., Lieutier, F., Eds.; Springer: Cham, Switzerland, 2016; pp. 47–88. [Google Scholar]
- Denlinger, D. Insect Diapause; Cambridge University Press: Cambridge, UK, 2022; p. 38. [Google Scholar]
- Tougeron, K.; Brodeur, J.; Le Lann, C.; Van Baaren, J. How climate change affects the seasonal ecology of insect parasitoids. Ecol. Entomol. 2019, 45, 167–181. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Type of Parasitoid (Host Groups Known) | Host in Rowanberries (Ahlberg 1927) [5] | References |
|---|---|---|---|---|
| Bracon (Glabrobracon) conjugellae (Bengtsson, 1924) | Braconidae | Idiobiont ectoparasitoid of concealed hosts (Megastigmus brevicaudis, Argyresthia conjugella, and Pontiana) | M. brevicaudis | [5,34,35,36] |
| Bracon(Glabrobracon) pulcher (Bengtsson, 1924) | Braconidae | Ectoparasitoid of concealed hosts (M. brevicaudis, A. conjugella, and Metzenera lapella) | M. brevicaudis | [5,34,35,36] |
| Dicladocerus westwoodii Westwood, 1832 | Eulophidae | Idiobiont ectoparasitoid (various Lepidoptera and Diptera) | A. conjugella 1 | [37,38] |
| Enytus neoapostata 2 (Horstmann, 1969) | Ichneumonidae | Koinobiont endoparasitoid (Depressaria assimilella) | This parasitoid not found by Ahlberg? 3 | [39,40] |
| Microgaster polita 4 Marshall, 1885 | Braconidae | Koinobiont endoparasitoid (A. conjugella) | A. conjugella | [5,41,42] |
| Scambus calobatus Gravenhorst, 1829 | Ichneumonidae | Idiobiont ectoparasitoid (various Lepidoptera, Coleoptera, and Hymenoptera) | A. conjugella 5 | [5,29,40,43] |
| Torymus aucupariae Rodzianko, 1908 | Torymidae | Not known? (Megastigmus) | M. brevicaudis 6 | [5,37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trandem, N.; Westrum, K.; Hofsvang, T.; Kobro, S. Distribution and Prolonged Diapause of the Rowan Seed Predators Argyresthia conjugella (Lepidoptera: Yponomeutidae) and Megastigmus brevicaudis (Hymenoptera: Torymidae) and their Parasitoids in Norway. Forests 2023, 14, 847. https://doi.org/10.3390/f14040847
Trandem N, Westrum K, Hofsvang T, Kobro S. Distribution and Prolonged Diapause of the Rowan Seed Predators Argyresthia conjugella (Lepidoptera: Yponomeutidae) and Megastigmus brevicaudis (Hymenoptera: Torymidae) and their Parasitoids in Norway. Forests. 2023; 14(4):847. https://doi.org/10.3390/f14040847
Chicago/Turabian StyleTrandem, Nina, Karin Westrum, Trond Hofsvang, and Sverre Kobro. 2023. "Distribution and Prolonged Diapause of the Rowan Seed Predators Argyresthia conjugella (Lepidoptera: Yponomeutidae) and Megastigmus brevicaudis (Hymenoptera: Torymidae) and their Parasitoids in Norway" Forests 14, no. 4: 847. https://doi.org/10.3390/f14040847