
Assessing the Carbon Storage Potential of a Young Mangrove Plantation in Myanmar

Abstract
:1. Introduction
2. Materials and Methods
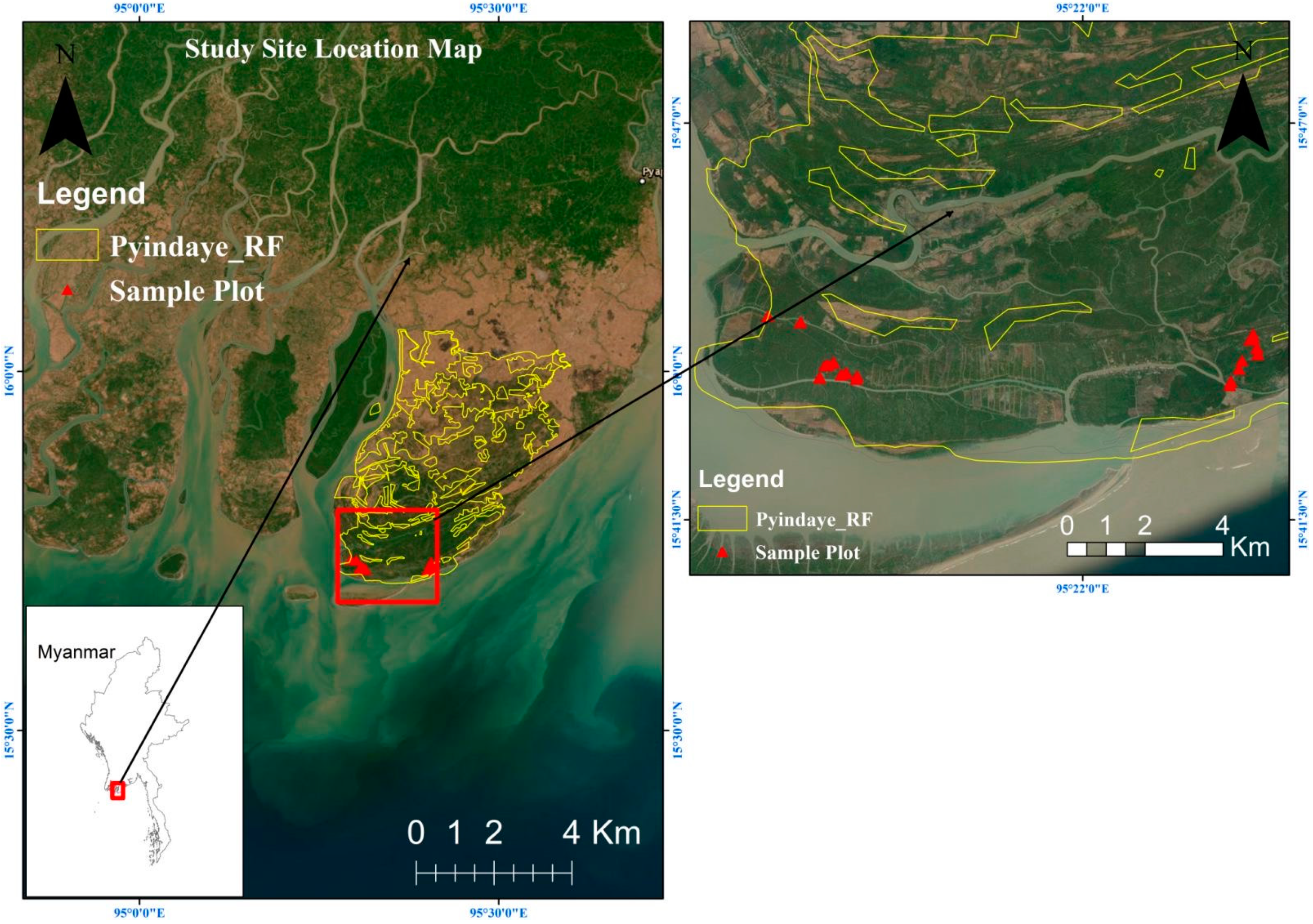
2.1. Study Site
2.2. Field Survey and Data Analysis
2.3. Statistical Analysis
3. Results
3.1. Forest Structure
3.2. Vegetation Biomass and Carbon Storage
3.3. The Relationship between Aboveground Biomass and Stand Basal Area
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Stand Age (Year) | Stem Density (ha−1) | Mean DBH (cm) | Mean H (m) | BA (m2 ha−1) | Biomass (Mg ha−1) | Carbon Stock (Mg C ha−1) | ||
|---|---|---|---|---|---|---|---|---|---|
| AGB | BGB | AGC | BGC | ||||||
| 1 | 4 | 2850 (114) | 10.34 | 3.62 | 29.18 | 194.28 | 85.27 | 91.31 | 33.25 |
| 2 | 4 | 1325 (53) | 6.21 | 2.08 | 4.21 | 23.13 | 11.97 | 10.87 | 4.67 |
| 3 | 4 | 1525 (61) | 17.49 | 4.70 | 43.55 | 359.04 | 141.0 | 168.75 | 54.99 |
| 4 | 4 | 2250 (90) | 13.67 | 4.00 | 42.94 | 348.56 | 138.86 | 163.82 | 54.16 |
| 5 | 4 | 2500 (100) | 8.13 | 3.04 | 15.83 | 102.60 | 47.22 | 48.22 | 18.41 |
| 6 | 4 | 2650 (106) | 14.12 | 4.49 | 42.55 | 385.95 | 163.62 | 181.40 | 63.81 |
| 7 | 4 | 1675 (67) | 13.33 | 3.94 | 27.48 | 208.77 | 86.55 | 98.12 | 33.75 |
| 8 | 4 | 1275 (51) | 11.06 | 3.72 | 19.51 | 169.90 | 65.65 | 79.85 | 25.60 |
| 9 | 3 | 1750 (70) | 9.71 | 3.11 | 16.50 | 117.03 | 49.93 | 55.00 | 19.47 |
| 10 | 3 | 2175 (87) | 8.43 | 2.95 | 18.55 | 123.75 | 52.76 | 58.16 | 20.58 |
| 11 | 5 | 1600 (64) | 11.20 | 4.12 | 16.34 | 103.29 | 47.29 | 48.54 | 18.44 |
| 12 | 5 | 1750 (70) | 11.46 | 3.85 | 19.10 | 119.61 | 54.32 | 56.21 | 21.19 |
| 13 | 5 | 1650 (66) | 12.17 | 3.97 | 23.01 | 176.92 | 74.15 | 83.15 | 28.92 |
| 14 | 5 | 1875 (75) | 12.53 | 4.13 | 25.40 | 180.44 | 78.15 | 84.81 | 30.48 |
| 15 | 5 | 2975 (119) | 10.9 | 3.72 | 30.17 | 200.5 | 90.39 | 94.24 | 35.25 |
| 16 | 3 | 2050 (82) | 13.00 | 4.09 | 30.86 | 213.00 | 91.20 | 100.11 | 35.57 |
| 17 | 3 | 2675 (107) | 15.58 | 4.60 | 55.18 | 403.47 | 168.45 | 189.63 | 65.69 |
| 18 | 5 | 1275 (51) | 20.67 | 6.10 | 47.96 | 406.82 | 156.60 | 191.21 | 61.07 |
| 19 | 3 | 2375 (95) | 10.56 | 3.17 | 29.77 | 226.95 | 93.01 | 106.67 | 36.27 |
| 20 | 3 | 2450 (98) | 14.26 | 4.30 | 42.55 | 368.91 | 153.62 | 173.39 | 59.91 |
| 21 | 3 | 2100 (84) | 9.87 | 3.07 | 22.90 | 174.02 | 71.73 | 81.79 | 27.98 |
| 22 | 5 | 2000 (80) | 10.09 | 3.54 | 19.54 | 141.24 | 61.38 | 66.38 | 23.94 |
| 23 | 5 | 2050 (82) | 10.35 | 3.41 | 22.65 | 167.39 | 70.20 | 78.67 | 27.38 |
| 24 | 3 | 1725 (69) | 12.75 | 3.91 | 27.41 | 208.44 | 85.93 | 97.97 | 33.51 |
| Mean ± SD | 2022 ± 492 | 12.00 ± 3.09 | 3.82 ± 0.78 | 28.045 ± 12.20 | 213.50 ± 107.87 | 89.14 ± 42.54 | 100.34 ± 50.70 | 34.76 ± 16.59 | |
| DBH Class (cm) | Tree Density | Avg. DBH (cm) | Basal Area (m2 ha−1) | Biomass (Mg ha−1) | Carbon Stock (Mg C ha−1) | ||
|---|---|---|---|---|---|---|---|
| AGB | BGB | AGC | BGC | ||||
| 5–10 | 23,350 (48.12%) | 7.43 | 106.96 | 614.62 | 306.04 | 288.87 | 119.36 |
| 11–15 | 14,825 (30.55%) | 12.78 | 192.44 | 1358.05 | 603.30 | 638.28 | 235.29 |
| 16–20 | 6375 (13.14%) | 17.39 | 152.37 | 1180.71 | 491.39 | 554.93 | 191.64 |
| 21–25 | 2475 (5.10%) | 22.77 | 101.19 | 832.34 | 326.92 | 391.20 | 127.50 |
| 26–30 | 650 (1.34%) | 27.69 | 39.23 | 350.85 | 131.74 | 164.909 | 51.38 |
| 31–35 | 525 (1.08%) | 32.48 | 43.54 | 411.41 | 148.92 | 193.36 | 58.08 |
| 36–40 | 325 (0.67%) | 38.23 | 37.36 | 376.01 | 130.95 | 176.73 | 51.07 |
References
- Karimi, Z.; Abdi, E.; Deljouei, A.; Cislaghi, A.; Shirvany, A.; Schwarz, M.; Hales, T.C. Vegetation-Induced Soil Stabilization in Coastal Area: An Example from a Natural Mangrove Forest. Catena (Amst.) 2022, 216, 106410. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the Most Carbon-Rich Forests in the Tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Heider, C.; Cole, T.G.; Dwire, K.A.; Donato, D.C. Ecosystem Carbon Stocks of Micronesian Mangrove Forests. Wetlands 2011, 31, 343–352. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Purbopuspito, J.; Kauffman, J.B.; Warren, M.W.; Sasmito, S.D.; Donato, D.C.; Manuri, S.; Krisnawati, H.; Taberima, S.; Kurnianto, S. The Potential of Indonesian Mangrove Forests for Global Climate Change Mitigation. Nat. Clim. Chang. 2015, 5, 1089–1092. [Google Scholar] [CrossRef]
- NGUYEN, H.-H.; Le, T.A.; Le, T.A.; Tran, T.N.L.; Tran, T.N.L.; Nguyen, H.N.; Nguyen, H.N.; Duong, V.K.L.; Duong, V.K.L.; Bohm, S.; et al. Biomass and carbon stock estimation of coastal mangroves at hai phong coast using remote sensing and field investigation- based data. Vietnam. J. Sci. Technol. 2021, 59, 560–579. [Google Scholar] [CrossRef]
- IUCN Real Potential to Restore Mangroves: New Tool Sheds Light | IUCN. Available online: https://www.iucn.org/news/forests/201810/real-potential-restore-mangroves-new-tool-sheds-light (accessed on 27 November 2022).
- Goldberg, L.; Lagomasino, D.; Thomas, N.; Fatoyinbo, T. Global Declines in Human-Driven Mangrove Loss. Glob. Chang. Biol. 2020, 26, 5844–5855. [Google Scholar] [CrossRef]
- Spalding, M.D.; Maricé, L. (Eds.) The State of the World’s Mangroves 2021; Global Mangrove Alliance, 2021. Available online: https://www.mangrovealliance.org/wp-content/uploads/2021/07/The-State-of-the-Worlds-Mangroves-2021-FINAL.pdf (accessed on 5 June 2022).
- FAO. The World’s Mangroves 1980–2005. Available online: http://www.fao.org/3/a-a1427e.pdf (accessed on 5 June 2022).
- Dahdouh-Guebas, F.; Jayatissa, L.P.; di Nitto, D.; Bosire, J.O.; Lo Seen, D.; Koedam, N. How Effective Were Mangroves as a Defence against the Recent Tsunami? Curr. Biol. 2005, 15, R443–R447. [Google Scholar] [CrossRef] [PubMed]
- Friess, D.A.; Yando, E.S.; Abuchahla, G.M.O.; Adams, J.B.; Cannicci, S.; Canty, S.W.J.; Cavanaugh, K.C.; Connolly, R.M.; Cormier, N.; Dahdouh-Guebas, F.; et al. Mangroves Give Cause for Conservation Optimism, for Now. Curr. Biol. 2020, 30, R153–R154. [Google Scholar] [CrossRef] [PubMed]
- Gedan, K.B.; Kirwan, M.L.; Wolanski, E.; Barbier, E.B.; Silliman, B.R. The Present and Future Role of Coastal Wetland Vegetation in Protecting Shorelines: Answering Recent Challenges to the Paradigm. Clim. Chang. 2011, 106, 7–29. [Google Scholar] [CrossRef]
- Kyi Myint, K.; Myat Nwe, M.; Lay Mar, T. Study on Morphological Characters of Some Mangrove Plants in South-Eastern Ayeyarwady Delta of Myanmar. J. Aquac. Mar. Biol. 2019, 8, 118–128. [Google Scholar] [CrossRef]
- Win, S. Tin-Zar-Ni-Win Vegetative Structure and Zonal Distribution of True Mangroves in Shwe-Thaung-Yan Coastal Areas, Myanmar. J. Aquac. Mar. Biol. 2021, 10, 33–39. [Google Scholar] [CrossRef]
- Forest Department. Mangrove Data Book; Forest Department Myanmar: Naypyitaw, Myanmar, 2017. [Google Scholar]
- Eid, E.M.; Arshad, M.; Shaltout, K.H.; El-Sheikh, M.A.; Alfarhan, A.H.; Picó, Y.; Barcelo, D. Effect of the Conversion of Mangroves into Shrimp Farms on Carbon Stock in the Sediment along the Southern Red Sea Coast, Saudi Arabia. Environ. Res. 2019, 176, 108536. [Google Scholar] [CrossRef]
- Ashton, E.C. Threats to Mangroves and Conservation Strategies. In Mangroves: Biodiversity, Livelihoods and Conservation; Springer: Singapore, 2022; pp. 217–230. [Google Scholar] [CrossRef]
- Zöckler, C.; Aung, C. The Mangroves of Myanmar. In Sabkha Ecosystems: Volume VI: Asia/Pacific; Gul, B., Böer, B., Khan, M.A., Clüsener-Godt, M., Hameed, A., Eds.; Springer International Publishing: Cham, Swizerland, 2019; pp. 253–268. [Google Scholar] [CrossRef]
- Ellison, A.M. Mangrove Restoration: Do We Know Enough? Restor. Ecol. 2000, 8, 219–229. [Google Scholar] [CrossRef]
- Hai, N.T.; Dell, B.; Phuong, V.T.; Harper, R.J. Towards a More Robust Approach for the Restoration of Mangroves in Vietnam. Ann. For. Sci. 2020, 77, 18. [Google Scholar] [CrossRef]
- Thant, Y.M.; Kanzaki, M.; Ohta, S.; Than, M.M. Carbon Sequestration by Mangrove Plantations and a Natural Regeneration Stand in the Ayeyarwady Delta, Myanmar. Tropics 2012, 21, 1–10. [Google Scholar] [CrossRef]
- Ahmed, S.; Kamruzzaman, M.; Rahman, M.S.; Sakib, N.; Azad, M.S.; Dey, T. Stand Structure and Carbon Storage of a Young Mangrove Plantation Forest in Coastal Area of Bangladesh: The Promise of a Natural Solution. Nat.-Based Solut. 2022, 2, 100025. [Google Scholar] [CrossRef]
- Fromard, F.; Puig, H.; Mougin, E.; Marty, G.; Betoulle, J.L.; Cadamuro, L. Structure, above-Ground Biomass and Dynamics of Mangrove Ecosystems: New Data from French Guiana. Oecologia 1998, 115, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Cuc, N.T.K.; Hien, H.T. Stand Structure and above ground biomass of Kandelia Obovata Sheue, H.Y. Liu & J. Yong Mangrove Plantations in Northern, Viet Nam. For. Ecol. Manag. 2021, 483, 118720. [Google Scholar] [CrossRef]
- JICA. The Study on Integrated Mangrove Management through Community Participation in the Ayeyawady Delta in the Union of Myanmar: Final Report; Burma. Ministry of Forestry, Kokusai Kyōryoku Jigyōdan, Ed.; Nippon Koei Co., Ltd.: Tokyo, Japan, 2005. [Google Scholar]
- Ono, K. Locally Appropriate Management of Mangrove Ecosystem—A Case Study in the Ayeyarwady Delta, Myanmar; Yokohama National University: Yokohama, Japan, 2007. [Google Scholar]
- Brakenridge, G.R.; Syvitski, J.P.M.; Niebuhr, E.; Overeem, I.; Higgins, S.A.; Kettner, A.J.; Prades, L. Design with Nature: Causation and Avoidance of Catastrophic Flooding, Myanmar. Undefined 2017, 165, 81–109. [Google Scholar] [CrossRef]
- Webb, E.L.; Than, M.M. Optimizing Investment Strategies for Mangrove Plantations by Considering Biological and Economic Parameters. J. Coast. Conserv. 2000, 6, 181–190. [Google Scholar] [CrossRef]
- Kogo, M. Final Report on Mangrove Reforestation Feasibility Study in the Ayeyarwady Delta; MYA/90/003 Field Document No.6; UNDP/FAO: Yangon, Myanmar, 1993. [Google Scholar]
- Komiyama, A.; Poungparn, S.; Kato, S. Common Allometric Equations for Estimating the Tree Weight of Mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a Worldwide Wood Economics Spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- IPCC 2006 IPCC Guidelines for National Greenhouse Gas Inventories—IPCC. Available online: https://www.ipcc.ch/report/2006-ipcc-guidelines-for-national-greenhouse-gas-inventories/ (accessed on 17 September 2021).
- Kauffman, J.B.; Donato, D.C. Protocols for the Measurement, Monitoring and Reporting of Structure, Biomass and Carbon Stocks in Mangrove Forests; CFIOR: Bogor, Indonesia, 2012. [Google Scholar] [CrossRef]
- Waters, H. Mangrove Restoration: Letting Mother Nature Do the Work. Available online: https://ocean.si.edu/ocean-life/plants-algae/mangrove-restoration-letting-mother-nature-do-work (accessed on 14 December 2022).
- Aye, W.M.; Takeda, S. Conversion of Abandoned Paddy Fields to Productive Land through Mangrove Restoration in Myanmar’s Ayeyarwady Delta. Paddy Water Environ. 2020, 18, 417–429. [Google Scholar] [CrossRef]
- Triest, L.; del Socorro, A.; Gado, V.J.; Mazo, A.M.; Sierens, T. Avicennia Genetic Diversity and Fine-Scaled Structure Influenced by Coastal Proximity of Mangrove Fragments. Front. Mar. Sci. 2021, 8, 638. [Google Scholar] [CrossRef]
- Miah, M.A.Q.; Islam, S.A.; Habib, M.A.; Moula, M.G. Growth Performance of Avicennia Officinalis L. and the Effect of Spacing on Growth and Yield of Trees Planted in the Western Coastal Belt of Bangladesh. J. For. Res. 2014, 25, 835–838. [Google Scholar] [CrossRef]
- IUCN Ecology and Management of Mangroves—Resource | IUCN. Available online: https://www.iucn.org/resources/publication/ecology-and-management-mangroves (accessed on 27 November 2022).
- Sharma, S.; Nadaoka, K.; Nakaoka, M.; Uy, W.H.; MacKenzie, R.A.; Friess, D.A.; Fortes, M.D. Growth Performance and Structure of a Mangrove Afforestation Project on a Former Seagrass Bed, Mindanao Island, Philippines. Hydrobiologia 2017, 803, 359–371. [Google Scholar] [CrossRef]
- Menezes, M.; Berger, U.; Worbes, M. Annual Growth Rings and Long-Term Growth Patterns of Mangrove Trees from the Bragança Peninsula, North Brazil. Wetl. Ecol. Manag. 2003, 11, 233–242. [Google Scholar] [CrossRef]
- Salmo, S.G.; Lovelock, C.; Duke, N.C. Vegetation and Soil Characteristics as Indicators of Restoration Trajectories in Restored Mangroves. Hydrobiologia 2013, 720, 1–18. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; Ahmed, S.; Paul, S.; Rahman, M.M.; Osawa, A. Stand Structure and Carbon Storage in the Oligohaline Zone of the Sundarbans Mangrove Forest, Bangladesh. For. Sci. Technol. 2018, 14, 23–28. [Google Scholar] [CrossRef]
- Sitoe, A.A.; Júnior, L.; Mandlate, C.; Guedes, B.S. Biomass and Carbon Stocks of Sofala Bay Mangrove Forests. Forests 2014, 5, 1967–1981. [Google Scholar] [CrossRef]
- Ross, M.S.; Ruiz, P.L.; Telesnicki, G.J.; Meeder, J.F. Estimating Above-Ground Biomass and Production in Mangrove Communities of Biscayne National Park, Florida (U.S.A.). Wetl. Ecol. Manag. 2001, 9, 27–37. [Google Scholar] [CrossRef]
- Ahmed, S.; Kamruzzaman, M. Species-Specific Biomass and Carbon Flux in Sundarbans Mangrove Forest, Bangladesh: Response to Stand and Weather Variables. Biomass Bioenergy 2021, 153, 106215. [Google Scholar] [CrossRef]
- Azad, M.S.; Kamruzzaman, M.; Osawa, A. Quantification and Understanding of Above and Belowground Biomass in Medium Saline Zone of the Sundarbans, Bangladesh: The Relationships with Forest Attributes. J. Sustain. For. 2019, 39, 331–345. [Google Scholar] [CrossRef]
- Mizanur Rahman, M.; Nabiul Islam Khan, M.; Fazlul Hoque, A.K.; Ahmed, I. Carbon Stock in the Sundarbans Mangrove Forest: Spatial Variations in Vegetation Types and Salinity Zones. Wetl. Ecol. Manag. 2015, 23, 269–283. [Google Scholar] [CrossRef]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-Monroy, V.H.; Castañeda-Moya, E.; Thomas, N.; van der Stocken, T. Mangrove Canopy Height Globally Related to Precipitation, Temperature and Cyclone Frequency. Nat. Geosci. 2018, 12, 40–45. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, Biomass, and Productivity of Mangrove Forests: A Review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Hozumi, K.; Yoda, K.; Kokawa, S.; Kira, T. Production Ecology of Tropical Rain Forests in Southwestern Cambodia: I. Plant Biomass. Nat. Life Southeast Asia 1969, 6, 1–51. [Google Scholar]
- Ogawa, H.; Yoda, K.; Ogino, K.; Kira, T. Comparative Ecological Studies on Three Main Types of Forest Vegetation in Thailand II. Plant Biomass. Nat. Life Southeast Asia 1965, 4, 49–80. [Google Scholar]
- Ball, M.C. Salinity Tolerance in the Mangroves Aegiceras Corniculatum and Avicennia Marina. I: Water Use in Relation to Growth, Carbon Partitioning, and Salt Balance. Aust. J. Plant. Physiol. 1988, 15, 447–464. [Google Scholar] [CrossRef]
- Kajimoto, T.; Osawa, A.; Usoltsev, V.A.; Abaimov, A.P. Biomass and Productivity of Siberian Larch Forest Ecosystems. Permafr. Ecosyst. 2010, 209, 99–122. [Google Scholar] [CrossRef]
- Trissanti, V.N.; Amalo, L.F.; Handayani, L.D.W.; Nugroho, D.; Yuliani, A.R.; Mulyana, D. The Estimation of Biomass and Carbon Stocks in Mangrove Forest Ecosystem of Karawang Regency, West Java. IOP Conf. Ser. Earth Environ. Sci. 2022, 1109, 012099. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of Plant Diversity to Ecosystems: Immediate, Filter and Founder Effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Gross, J.; Flores, E.E.; Schwendenmann, L. Stand Structure and Aboveground Biomass of a Pelliciera Rhizophorae Mangrove Forest, Gulf of Monitjo Ramsar Site, Pacific Coast, Panama. Wetlands 2014, 34, 55–65. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; Ahmed, S.; Osawa, A. Biomass and Net Primary Productivity of Mangrove Communities along the Oligohaline Zone of Sundarbans, Bangladesh. For. Ecosyst. 2017, 4, 1–9. [Google Scholar] [CrossRef]
- Ragavan, P.; Kumar, S.; Kathiresan, K.; Mohan, P.M.; Jayaraj, R.S.C.; Ravichandaran, K.; Rana, T.S. Biomass and Vegetation Carbon Stock in Mangrove Forests of the Andaman Islands, India. Hydrobiologia 2021, 848, 4673–4693. [Google Scholar] [CrossRef]
- Torres, A.B.; Lovett, J.C. Using Basal Area to Estimate Aboveground Carbon Stocks in Forests: La Primavera Biosphere’s Reserve, Mexico. Forestry 2013, 86, 267–281. [Google Scholar] [CrossRef]




| No. | Species | Family | Stems ha−1 | Basal Area (m2 ha−1) | Biomass (Mg ha−1) | Carbon Stock (Mg C ha−1) | ||
|---|---|---|---|---|---|---|---|---|
| AGB | BGB | AGC | BGC | |||||
| 1. | Avicennia officinalis L. | Acanthaceae | 17,925 | 357.71 | 2731.39 | 1107.99 | 1283.75 | 432.12 |
| 2. | Avicennia marina (Forssk.) Vierh. | Acanthaceae | 11,450 | 157.72 | 1189.24 | 499.14 | 558.94 | 194.66 |
| 3. | Bruguiera gymnorhiza (L.) Lam. | Rhizophoraceae | 6725 | 21.52 | 126.25 | 65.03 | 59.34 | 25.36 |
| 4. | Bruguiera sexangula (Lour.) Poir. | Rhizophoraceae | 5750 | 32.69 | 217.30 | 103.36 | 102.13 | 40.31 |
| 5. | Rhizophora apiculata Blume. | Rhizophoraceae | 4000 | 64.20 | 594.82 | 251.74 | 279.57 | 98.18 |
| 6. | Heritiera fomes Banks. | Malvaceae | 1600 | 15.83 | 134.01 | 58.47 | 62.98 | 22.80 |
| 7. | Excoecaria agallocha L. | Euphorbiaceae | 775 | 22.16 | 123.67 | 49.87 | 58.13 | 19.45 |
| 8. | Ceriops decandra (Griff.) W.Theob. | Rhizophoraceae | 125 | 0.63 | 4.12 | 2.01 | 1.94 | 0.78 |
| 9. | Xylocarpus moluccensis (Lam.) M.Roem. | Meliaceae | 125 | 0.51 | 2.51 | 1.30 | 1.18 | 0.51 |
| 10. | Rhizophora mucronata Poir. | Rhizophoraceae | 50 | 0.12 | 0.69 | 0.37 | 0.32 | 0.14 |
| Stand Age (Year) | Avg. Stem Density (ha−1) | Mean DBH (cm) | Mean H (m) | BA (m2 ha−1) | Biomass (Mg ha−1) | Carbon Stock (Mg C ha−1) | ||
|---|---|---|---|---|---|---|---|---|
| AGB | BGB | AGC | BGC | |||||
| 3 | 2163 ± 331 | 11.77 ± 2.50 | 3.65 ± 0.65 | 30.46 ± 12.88 | 229.45 ± 105.11 | 95.83 ± 43.56 | 107.84 ± 49.40 | 37.37 ± 16.99 |
| 4 | 2006 ± 629 | 11.79 ± 3.61 | 3.70 ± 0.83 | 28.16 ± 14.46 | 224.03 ± 130.44 | 92.52 ± 51.97 | 105.29 ± 61.31 | 36.08 ± 20.27 |
| 5 | 1897 ± 501 | 12.43 ± 3.43 | 4.11 ± 0.85 | 25.52 ± 10.01 | 187.03 ± 94.63 | 79.06 ± 34.18 | 87.90 ± 44.47 | 30.83 ± 13.33 |
| Parameters | DBH (cm) | Height (m) | Stem Density (ha−1) | Basal Area (m2 ha−1) | Crown Area (m2) |
|---|---|---|---|---|---|
| Height (m) | 0.928 *** | - | |||
| Stem density (n ha−1) | −0.147 | −0.131 | - | - | |
| Basal area (m2 ha−1) | 0.830 *** | 0.694 *** | 0.372 | - | |
| Crown area (m2) | 0.907 *** | 0.845 *** | −0.336 * | 0.632 *** | - |
| AGB (Mg ha−1) | 0.824 *** | 0.671 *** | 0.340 | 0.987 *** | 0.580 *** |
| Variable | Coefficient | Standard Error | t-Statistics | p-Value |
|---|---|---|---|---|
| Intercept | 1.37 | 0.149 | 9.219 | 0.0000 *** |
| Stand basal area (m2/ha) | 1.19 | 0.449 | 26.524 | 0.0000 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aye, W.N.; Tong, X.; Li, J.; Tun, A.W. Assessing the Carbon Storage Potential of a Young Mangrove Plantation in Myanmar. Forests 2023, 14, 824. https://doi.org/10.3390/f14040824
Aye WN, Tong X, Li J, Tun AW. Assessing the Carbon Storage Potential of a Young Mangrove Plantation in Myanmar. Forests. 2023; 14(4):824. https://doi.org/10.3390/f14040824
Chicago/Turabian StyleAye, Wai Nyein, Xiaojuan Tong, Jun Li, and Aung Wunna Tun. 2023. "Assessing the Carbon Storage Potential of a Young Mangrove Plantation in Myanmar" Forests 14, no. 4: 824. https://doi.org/10.3390/f14040824