
Interspecific Differences of Stem Diameter Variations in Response to Water Conditions for Six Tree Species in Northeast China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Species Selection
2.2. Installation of the Measuring Instruments
2.3. Meteorological Data
2.4. Extraction of Stem Radial Variation and Statistical Analysis
3. Results
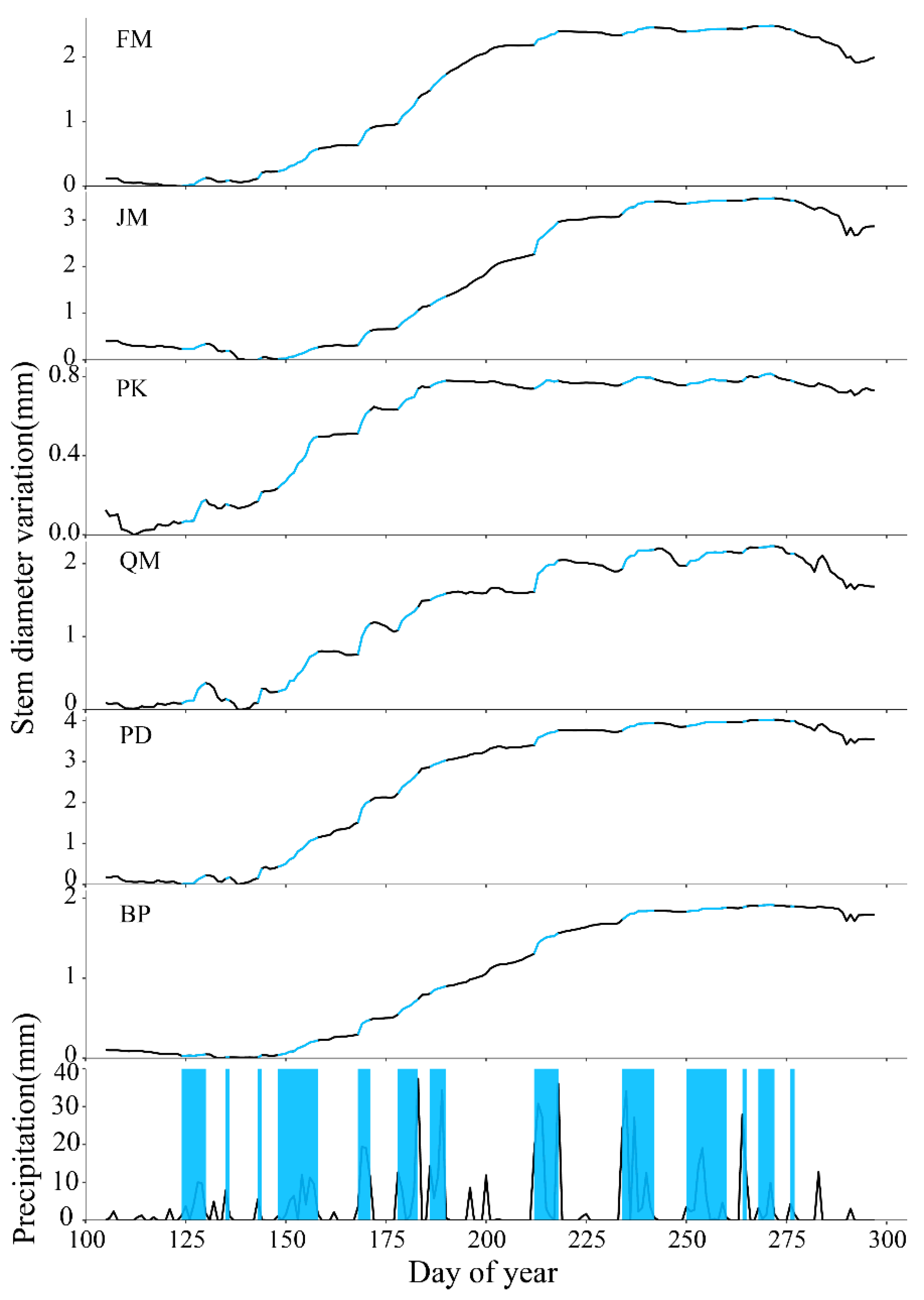
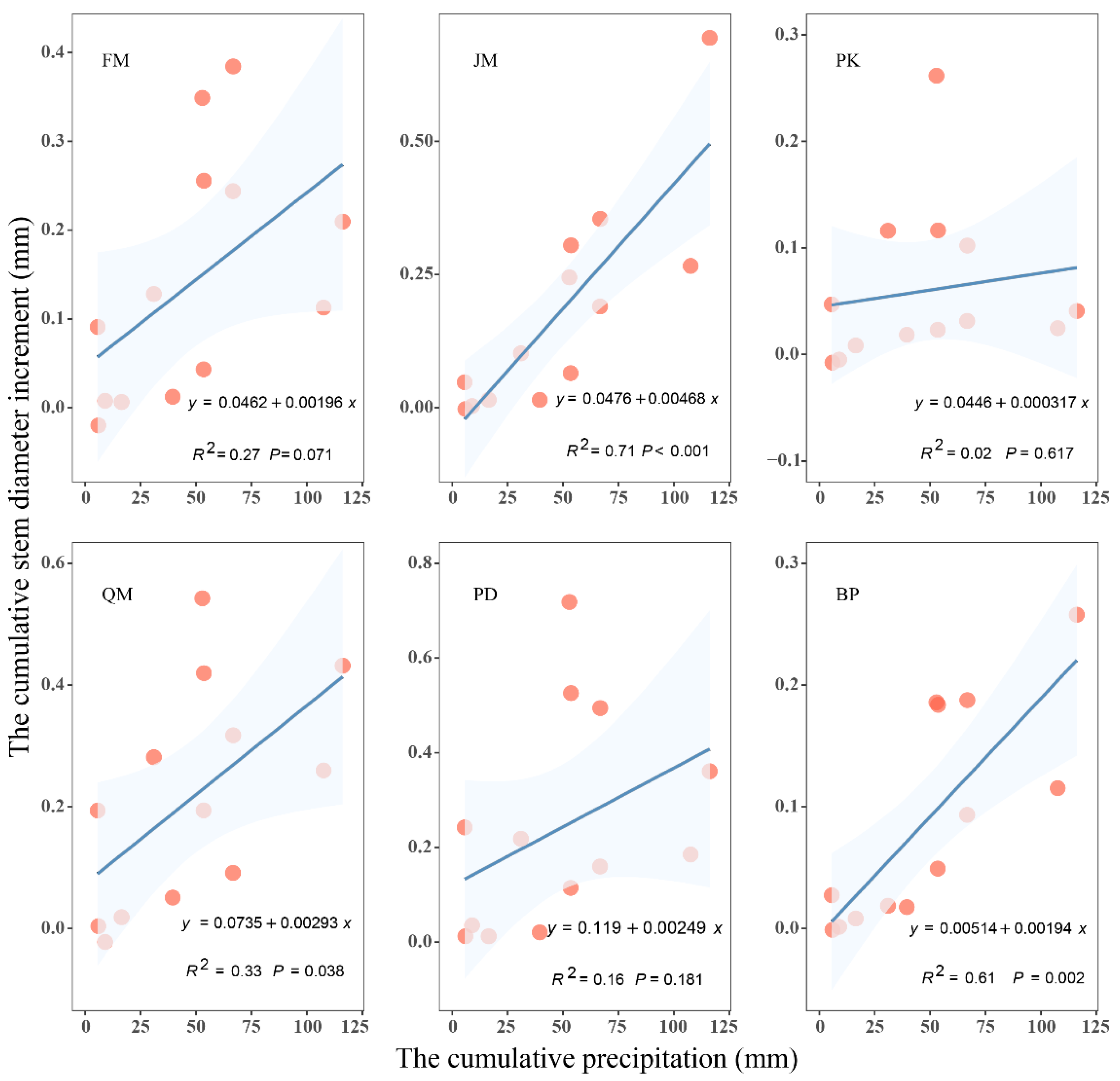
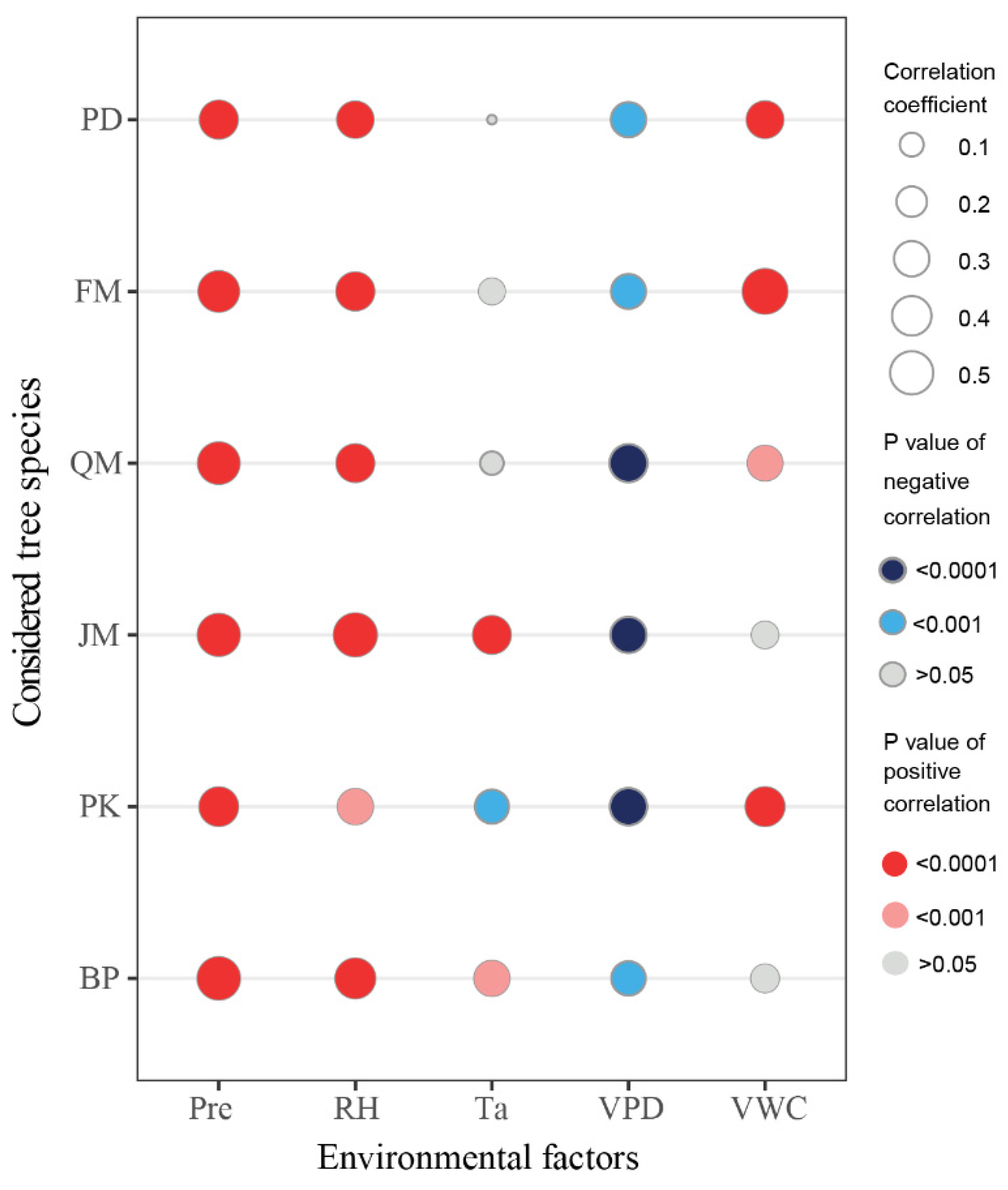
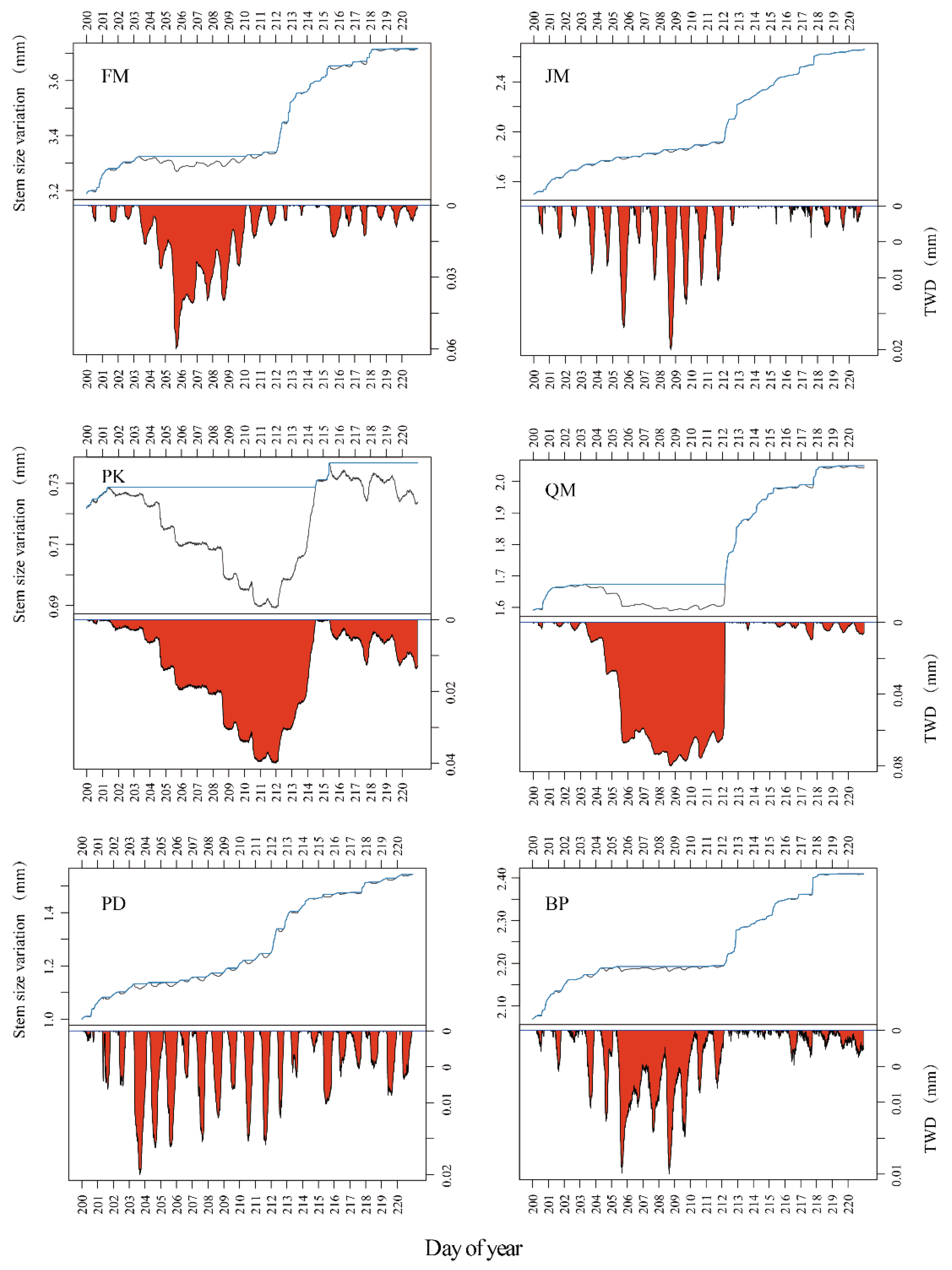
3.1. Relationship between Stem Radial Growth and Water Conditions
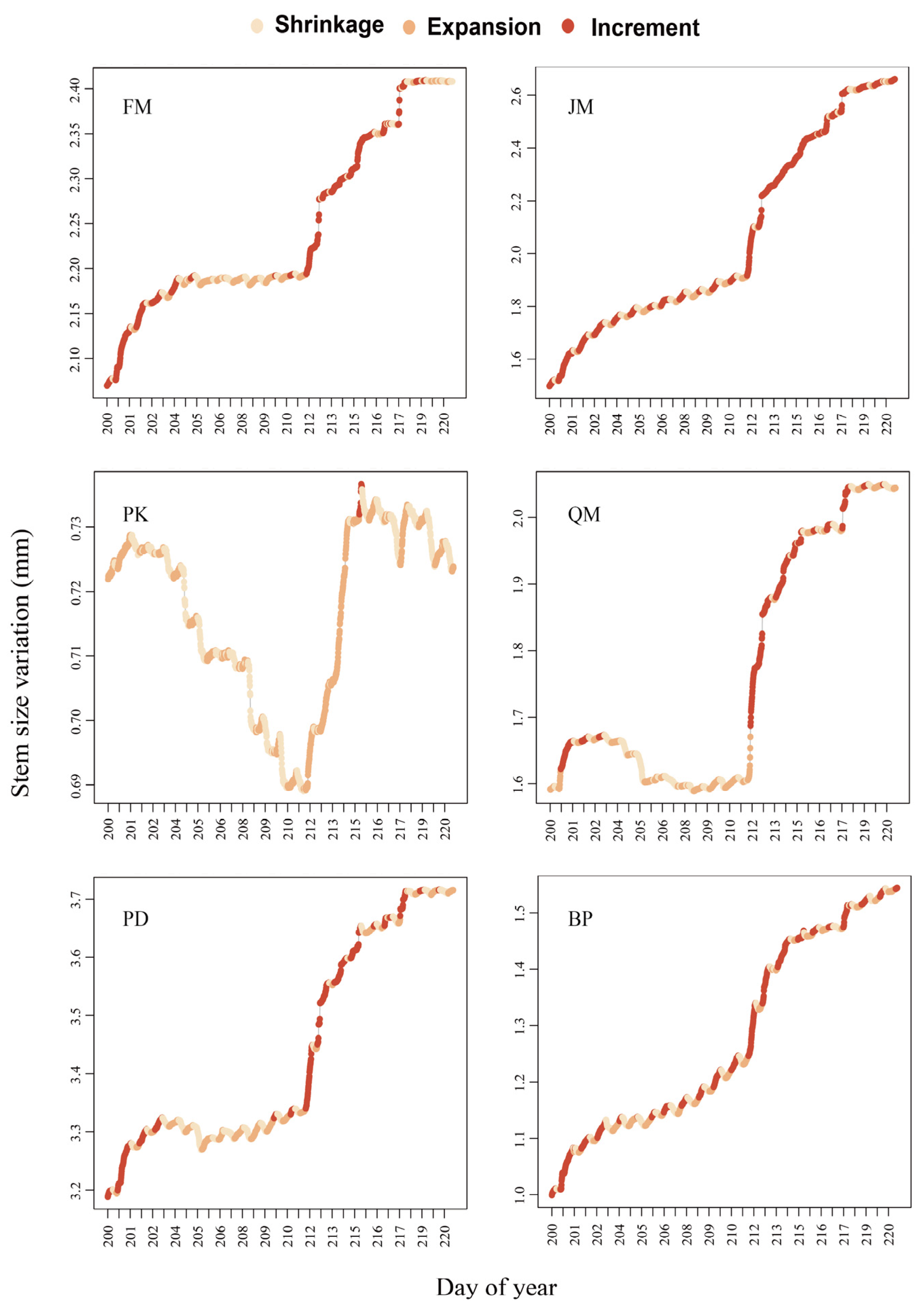
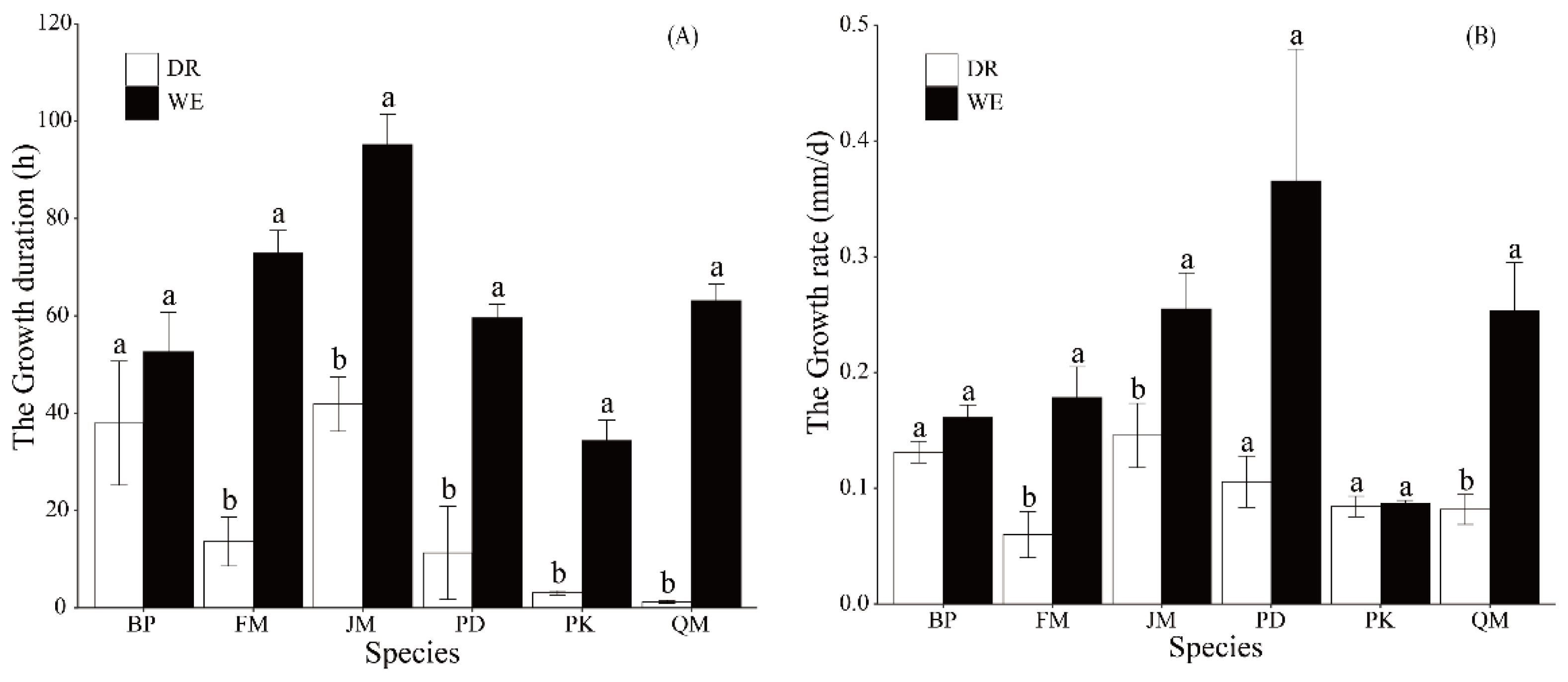
3.2. Interspecific Differences in Stem Diameter Variation Patterns under Drought Stress
4. Discussion
4.1. Water Conditions Affect Stem Radial Growth
4.2. Species-Specific Drought Response
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Pugh, T.A.M.; Lindeskog, M.; Smith, B.; Poulter, B.; Arneth, A.; Haverd, V.; Calle, L. Role of forest regrowth in global carbon sink dynamics. Proc. Natl. Acad. Sci. USA 2019, 116, 4382–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Zweifel, R.; Haeni, M.; Buchmann, N.; Eugster, W. Are trees able to grow in periods of stem shrinkage? New Phytol. 2016, 211, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Steppe, K.; Sterck, F.J.; Deslauriers, A. Diel growth dynamics in tree stems: Linking anatomy and ecophysiology. Trends Plant Sci. 2015, 20, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Yuan, Y.; Gou, X.; Zhang, T.; Zou, C.; Ji, C.; Fan, Z.; Qin, L.; Shang, H.; Li, X. Intra-annual radial growth of Schrenk spruce (Picea schrenkiana Fisch. et Mey) and its response to climate on the northern slopes of the Tianshan Mountains. Dendrochronologia 2016, 40, 36–42. [Google Scholar] [CrossRef]
- Wu, L.; Kaewmano, A.; Fu, P.; Wang, W.; Fan, Z. Intra-annual Radial Growth of Melia azedarach in a Tropical Moist Seasonal Forest and its Response to Environmental Factors in Xishuangbanna, Southwest China. Acta Ecol. Sin. 2020, 40, 6831–6840. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Urbinati, C.; Carrer, M. Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Quebec (Canada). Trees-Struct. Funct. 2003, 17, 477–484. [Google Scholar] [CrossRef]
- Martin, J.; Looker, N.; Hoylman, Z.; Jencso, K.; Hu, J. Hydrometeorology organizes intra-annual patterns of tree growth across time, space and species in a montane watershed. New Phytol. 2017, 215, 1387–1398. [Google Scholar] [CrossRef] [Green Version]
- Aldea, J.; Bravo, F.; Vazquez-Pique, J.; Ruiz-Peinado, R.; del Rio, M. Differences in stem radial variation between Pinus pinaster Ait. and Quercus pyrenaica Willd. may release inter-specific competition. For. Ecol. Manag. 2021, 481, 118779. [Google Scholar] [CrossRef]
- Etzold, S.; Sterck, F.; Bose, A.K.; Braun, S.; Buchmann, N.; Eugster, W.; Gessler, A.; Kahmen, A.; Peters, R.L.; Vitasse, Y.; et al. Number of growth days and not length of the growth period determines radial stem growth of temperate trees. Ecol. Lett. 2022, 25, 427–439. [Google Scholar] [CrossRef]
- Meng, S.-W.; Yang, F.-T.; Dai, X.-Q.; Wang, H.-M. Radial growth dynamics of Chinese fir and its response to seasonal drought. Chin. J. Appl. Ecol. 2021, 32, 3521–3530. [Google Scholar] [CrossRef]
- Sang, Y.; Wang, C.; Huo, H. Inter-specific and seasonal variations in photosynthetic capacity and water use efficiency of five temperate tree species in Northeastern China. Scand. J. For. Res. 2011, 26, 21–29. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Wang, C.K. Chinese Ecosystem Long-Term Observation and Research Dataset. Forest Ecosystem: Heilongjiang Maoershan Station (1979–2017); China Agriculture Press: Beijing, China, 2022. [Google Scholar]
- Wang, C.; Yang, J.; Zhang, Q. Soil respiration in six temperate forests in China. Glob. Chang. Biol. 2006, 12, 2103–2114. [Google Scholar] [CrossRef]
- Liu, F.; Wang, C.; Wang, X.; Zhang, J.; Zhang, Z.; Wang, J. Spatial patterns of biomass in the temperate broadleaved deciduous forest within the fetch of the Maoershan flux tower. Acta Ecol. Sin. 2016, 36, 6506–6519. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Wang, C.K.; Zhou, Z.H. Does the net primary production converge across six temperate forest types under the same climate? For. Ecol. Manag. 2019, 448, 535–542. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. An Introduction to Environmental Biophysics, 2nd ed.; Springer: New York, NY, USA, 1998. [Google Scholar]
- Downes, G.; Beadle, C.; Worledge, D. Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees 1999, 14, 102–111. [Google Scholar] [CrossRef]
- Dietrich, L.; Zweifel, R.; Kahmen, A. Daily stem diameter variations can predict the canopy water status of mature temperate trees. Tree Physiol. 2018, 38, 941–952. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Aryal, S.; Husser, M.; Grieinger, J.; Fan, Z.X.; Bruning, A. “dendRoAnalyst”: A tool for processing and analysing dendrometer data. Dendrochronologia 2020, 64, 125772. [Google Scholar] [CrossRef]
- Hu, L.; Fan, Z. Stem radial growth in response to microclimate in an Asian tropical dry karst forest. Acta Ecol. Sin. 2016, 36, 401–409. [Google Scholar] [CrossRef]
- Liu, C.; Kang, S. Assessment of trunk diameter fluctuation for irrigation schedule in apple trees. Acta Ecol. Sin. 2015, 35, 4754–4762. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Zeugin, F.; Newbery, D.M. Intra-annual radial growth and water relations of trees: Implications towards a growth mechanism. J. Exp. Bot. 2006, 57, 1445–1459. [Google Scholar] [CrossRef] [Green Version]
- Muller, B.; Pantin, F.; Genard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Anfodillo, T.; Cufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gricar, J.; Gruber, A.; King, G.M.; et al. A meta-analysis of cambium phenology and growth: Linear and non-linear patterns in conifers of the northern hemisphere. Ann. Bot. 2013, 112, 1911–1920. [Google Scholar] [CrossRef]
- Gao, J.; Yang, B.; Qin, C. Response of intra-annual stem radial growth to drought events: A case study of Pinus tabuliformis in the Helan Mountains, China. Chin. J. Appl. Ecol. 2021, 32, 3505–3511. [Google Scholar] [CrossRef]
- Raffelsbauer, V.; Spannl, S.; Peña, K.; Pucha-Cofrep, D.; Steppe, K.; Bräuning, A. Tree Circumference Changes and Species-Specific Growth Recovery after Extreme Dry Events in a Montane Rainforest in Southern Ecuador. Front. Plant Sci. 2019, 10, 342. [Google Scholar] [CrossRef]
- Aldea, J.; Bravo, F.; Vazquez-Pique, J.; Rubio-Cuadrado, A.; del Rio, M. Species-specific weather response in the daily stem variation cycles of Mediterranean pine-oak mixed stands. Agric. For. Meteorol. 2018, 256, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Ernst, V.D.M.; Jonas, P.; Marieke, V.D.M.-T.; Tobias, S.; Marko, S.; Roberto, C.-G.; Martin, W. Distinct growth phenology but similar daily stem dynamics in three co-occurring broadleaved tree species. Tree Physiol. 2018, 38, 1820–1828. [Google Scholar] [CrossRef]
- Alday, J.G.; Julio Camarero, J.; Revilla, J.; Resco de Dios, V. Similar diurnal, Seasonal and annual rhythems in radial root expansion across two coexiting Mediterranean oak species. Tree Physiol. 2020, 40, 956–968. [Google Scholar] [CrossRef] [PubMed]
- Obojes, N.; Meurer, A.; Newesely, C.; Tasser, E.; Oberhuber, W.; Mayr, S.; Tappeiner, U. Water stress limits transpiration and growth of European larch up to the lower subalpine belt in an inner-alpine dry valley. New Phytol. 2018, 220, 460–475. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Wang, C. Leaf hydraulic traits and their trade-offs for nine Chinese temperate tree species with different wood properties. Chin. J. Plant Ecol. 2016, 40, 702–710. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; Holbrook, N.M.; Zwieniecki, M.A.; Palma, B. Leaf hydraulic capacity in ferns, conifers and angiosperms: Impacts on photosynthetic maxima. New Phytol. 2005, 165, 839–846. [Google Scholar] [CrossRef]
- Li, J.; Zhai, H. Hydraulic architecture and drought resistance of woody plants. Chin. J. Appl. Ecol. 2000, 11, 301–305. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Meinzer, F.C.; Sternberg, L.D.L. Dynamic changes in hydraulic conductivity in petioles of two savanna tree species: Factors and mechanisms contributing to the refilling of embolized vessels. Plant Cell Environ. 2003, 26, 1633–1645. [Google Scholar] [CrossRef]
- Nardini, A.; Salleo, S.; Raimondo, F. Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L. Trees-Struct. Funct. 2003, 17, 529–534. [Google Scholar] [CrossRef]
- Nardini, A.; Peda, G.; La Rocca, N. Trade-offs between leaf hydraulic capacity and drought vulnerability: Morpho-anatomical bases, carbon costs and ecological consequences. New Phytol. 2012, 196, 788–798. [Google Scholar] [CrossRef]
- Gueney, A.; Zweifel, R.; Turkan, S.; Zimmermann, R.; Wachendorf, M.; Guney, C.O. Drought responses and their effects on radial stem growth of two co-occurring conifer species in the Mediterranean mountain range. Ann. For. Sci. 2020, 77, 105. [Google Scholar] [CrossRef]
- Duan, R.; Sun, H. Evaluation on Drought Tolerance of the Major Tree Species in the Eastern Mountain Region of Northeast China. Acta Bot. Boreali-Occident. Sin. 2019, 39, 156–163. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species (Code) | Habitat | Wood Property | Leaf Habit | DBH (cm) |
|---|---|---|---|---|
| Pinus koraiensis Sieb. et Zucc. (PK) | Mid slope | Non-porous | Evergreen-coniferous | 22.6 ± 0.7 |
| Fraxinus mandshurica (FM) | Valley bottom | Ring-porous | Deciduous-broadleaved | 35.4 ± 1.6 |
| Juglans mandshurica Maxim. (JM) | Valley bottom | Ring-porous | Deciduous-broadleaved | 37.7 ± 1.0 |
| Quercus mongolica Fisch. (QM) | Upper slope | Ring-porous | Deciduous-broadleaved | 34.0 ± 1.0 |
| Populous davidiana Dode. (PD) | Upper slope | Diffuse-porous | Deciduous-broadleaved | 49.6 ± 0.8 |
| Betula platyphylla Suk. (BP) | Mid slope | Diffuse-porous | Deciduous-broadleaved | 27.6 ± 1.1 |
| Period of Precipitation | Cumulative Precipitation (mm) | Proportion of Net Annual Growth (%) | |||||
|---|---|---|---|---|---|---|---|
| FM | JM | PK | QM | PD | BP | ||
| DOY 124-130 | 30.98 | 5.00 | / | 15.10 | / | / | / |
| DOY 135-136 | 8.95 | 0.31 | / | / | / | / | 0.14 |
| DOY 143-144 | 5.58 | 3.56 | 1.38 | 5.86 | 8.22 | 5.99 | 1.38 |
| DOY 148-158 | 52.82 | 13.82 | 7.03 | 34.16 | 23.44 | 17.84 | 9.70 |
| DOY 168-171 | 53.55 | 10.12 | 8.79 | 15.10 | 18.10 | 13.07 | 9.65 |
| DOY 178-183 | 66.76 | 14.11 | 9.56 | 13.09 | 13.74 | 11.79 | 9.80 |
| DOY 186-190 | 66.67 | 9.56 | 5.48 | 4.07 | 3.93 | 3.94 | 4.88 |
| DOY 212-218 | 116.15 | 8.29 | 20.13 | 5.26 | 18.65 | 8.97 | 13.52 |
| DOY 234-242 | 107.56 | 4.46 | 7.76 | 3.26 | 11.43 | 4.63 | 5.94 |
| DOY 250-260 | 53.39 | 1.73 | 1.85 | 2.87 | 8.34 | 2.83 | 2.56 |
| DOY 264-265 | 39.46 | 0.49 | 0.41 | 2.51 | 2.13 | 0.50 | 0.95 |
| DOY 268-272 | 16.42 | 0.26 | 0.48 | 1.14 | 0.77 | 0.30 | 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhang, Q. Interspecific Differences of Stem Diameter Variations in Response to Water Conditions for Six Tree Species in Northeast China. Forests 2023, 14, 805. https://doi.org/10.3390/f14040805
Wang X, Zhang Q. Interspecific Differences of Stem Diameter Variations in Response to Water Conditions for Six Tree Species in Northeast China. Forests. 2023; 14(4):805. https://doi.org/10.3390/f14040805
Chicago/Turabian StyleWang, Xueyu, and Quanzhi Zhang. 2023. "Interspecific Differences of Stem Diameter Variations in Response to Water Conditions for Six Tree Species in Northeast China" Forests 14, no. 4: 805. https://doi.org/10.3390/f14040805