
Evaluating the Legacy Effects of the Historical Predatory Seed Harvesting on the Species Composition and Structure of the Mixed Korean Pine and Broadleaf Forest from a Landscape Perspective

Abstract
:1. Introduction
2. Materials and Methods
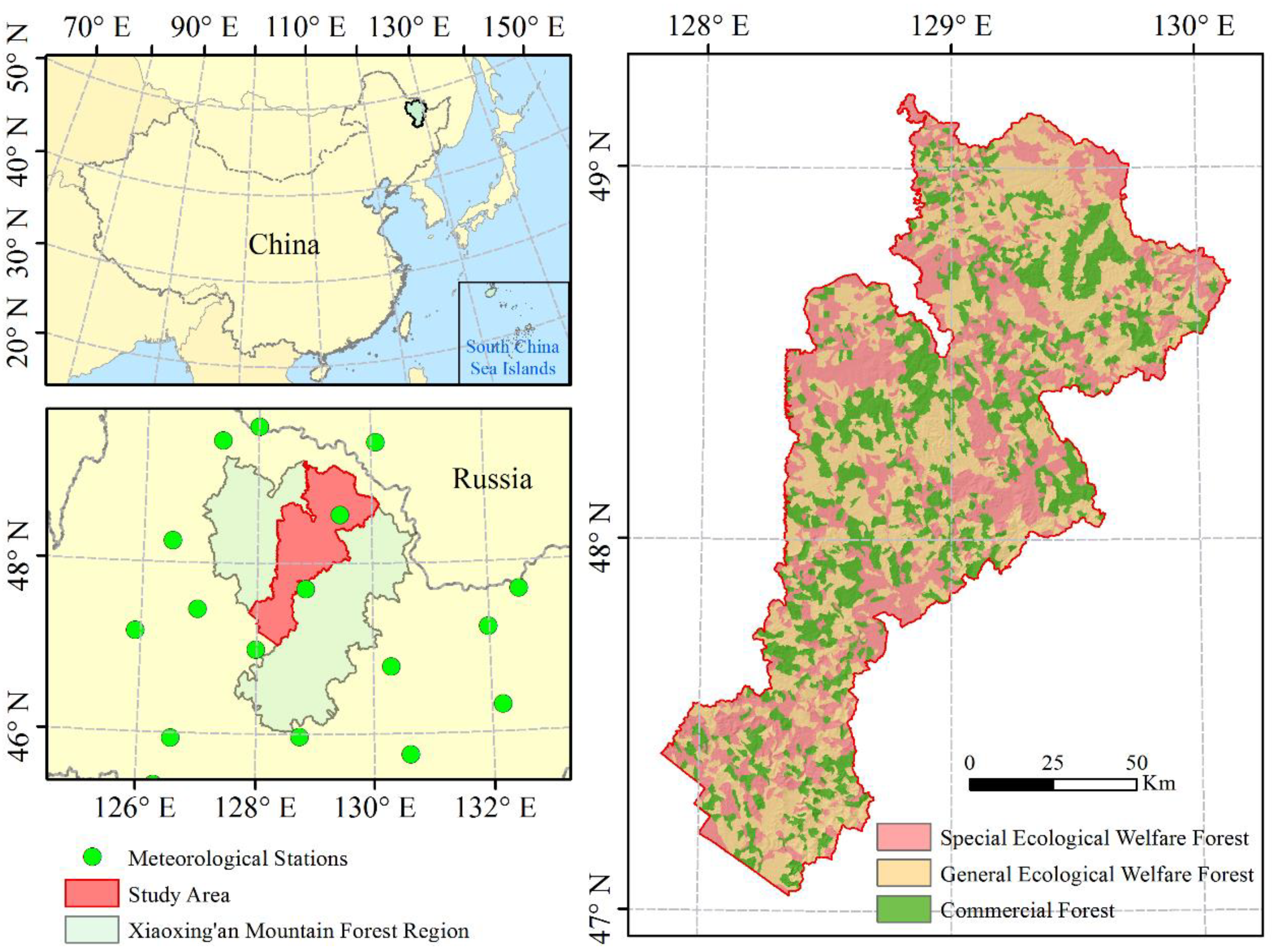
2.1. Study Area
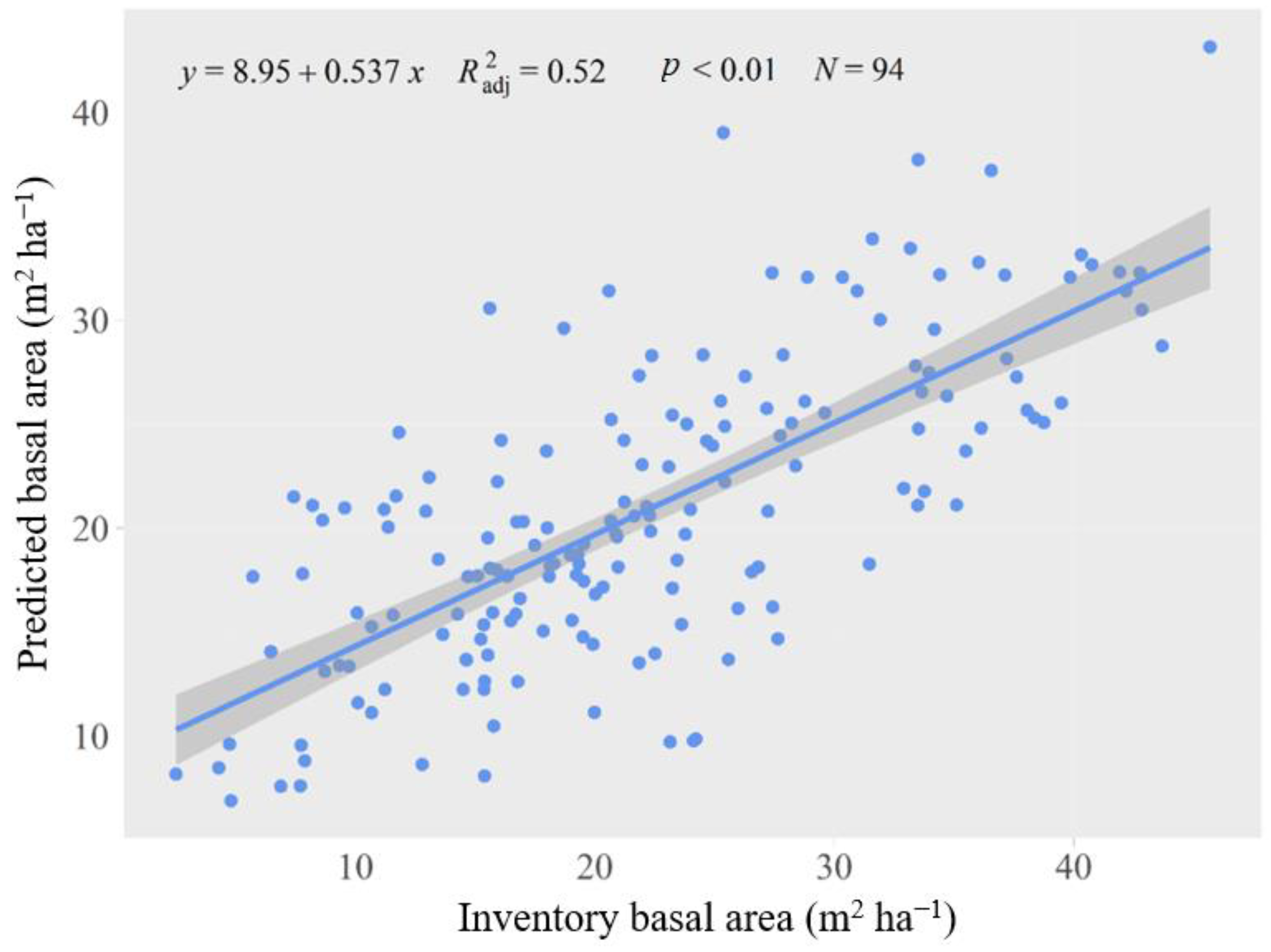
2.2. Forest Landscape Model and Parameterization
2.3. Experimental Design
2.4. Data Analysis
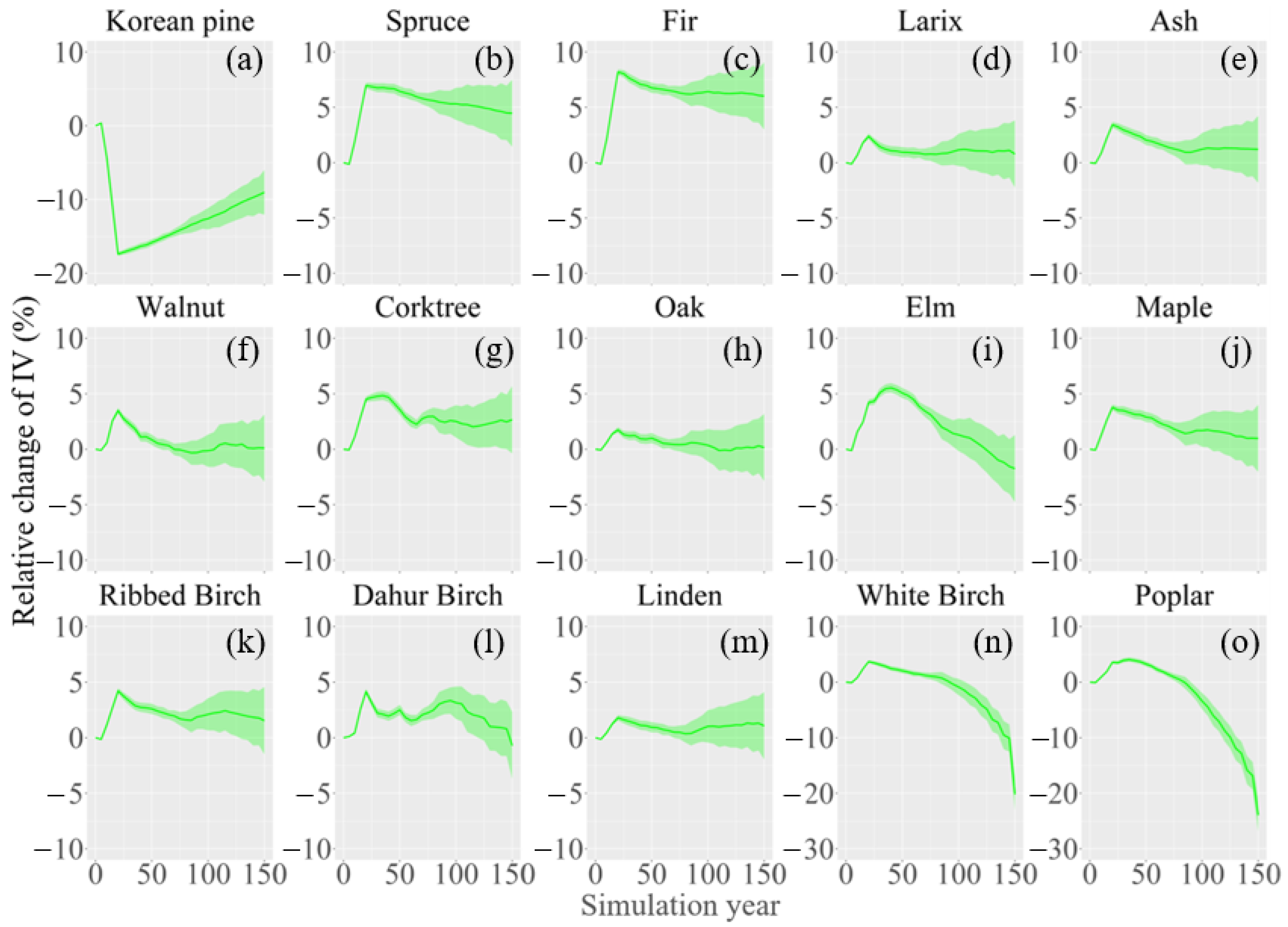
3. Results
3.1. Change in the Age Structure of Korean Pine
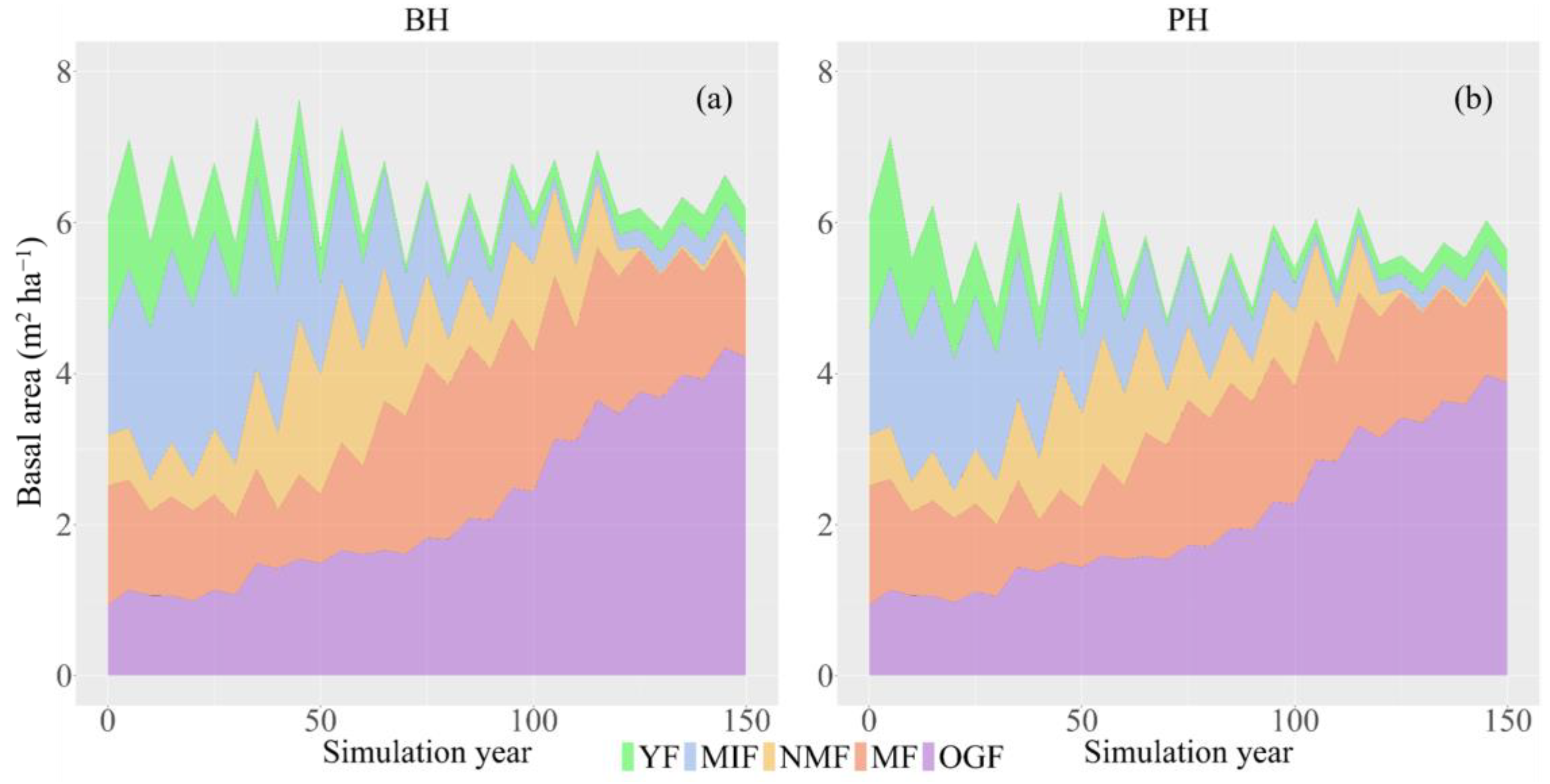
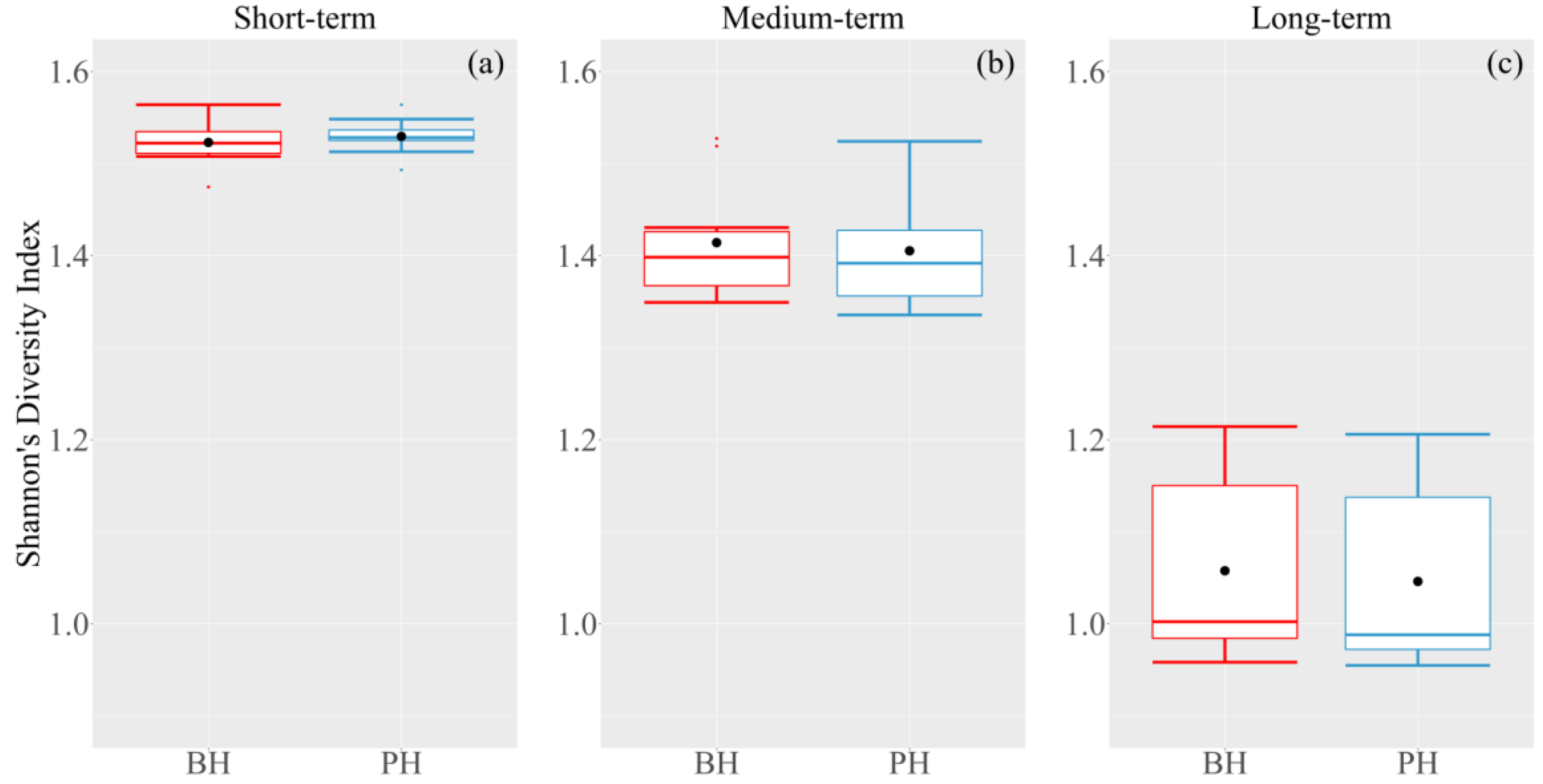
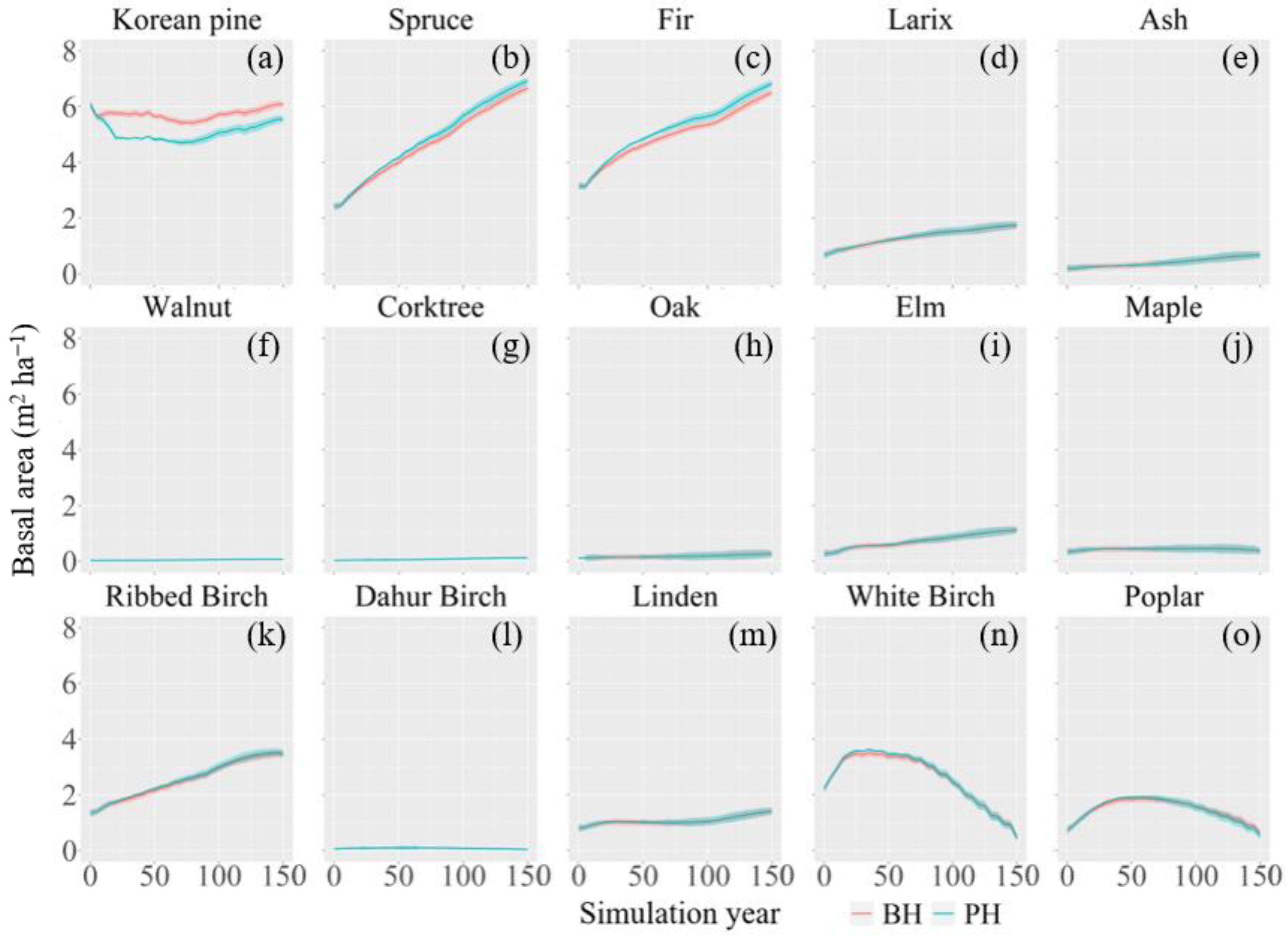
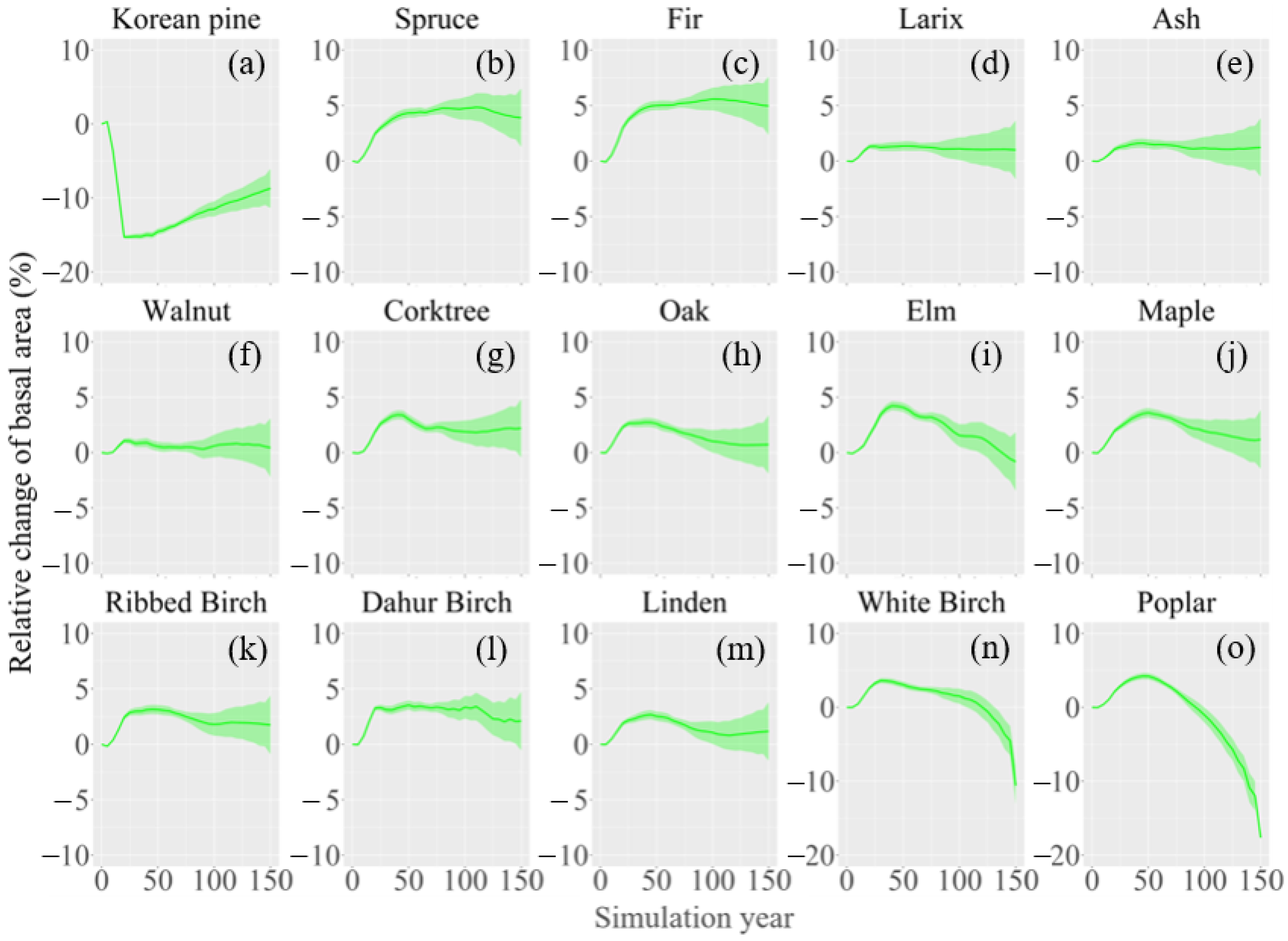
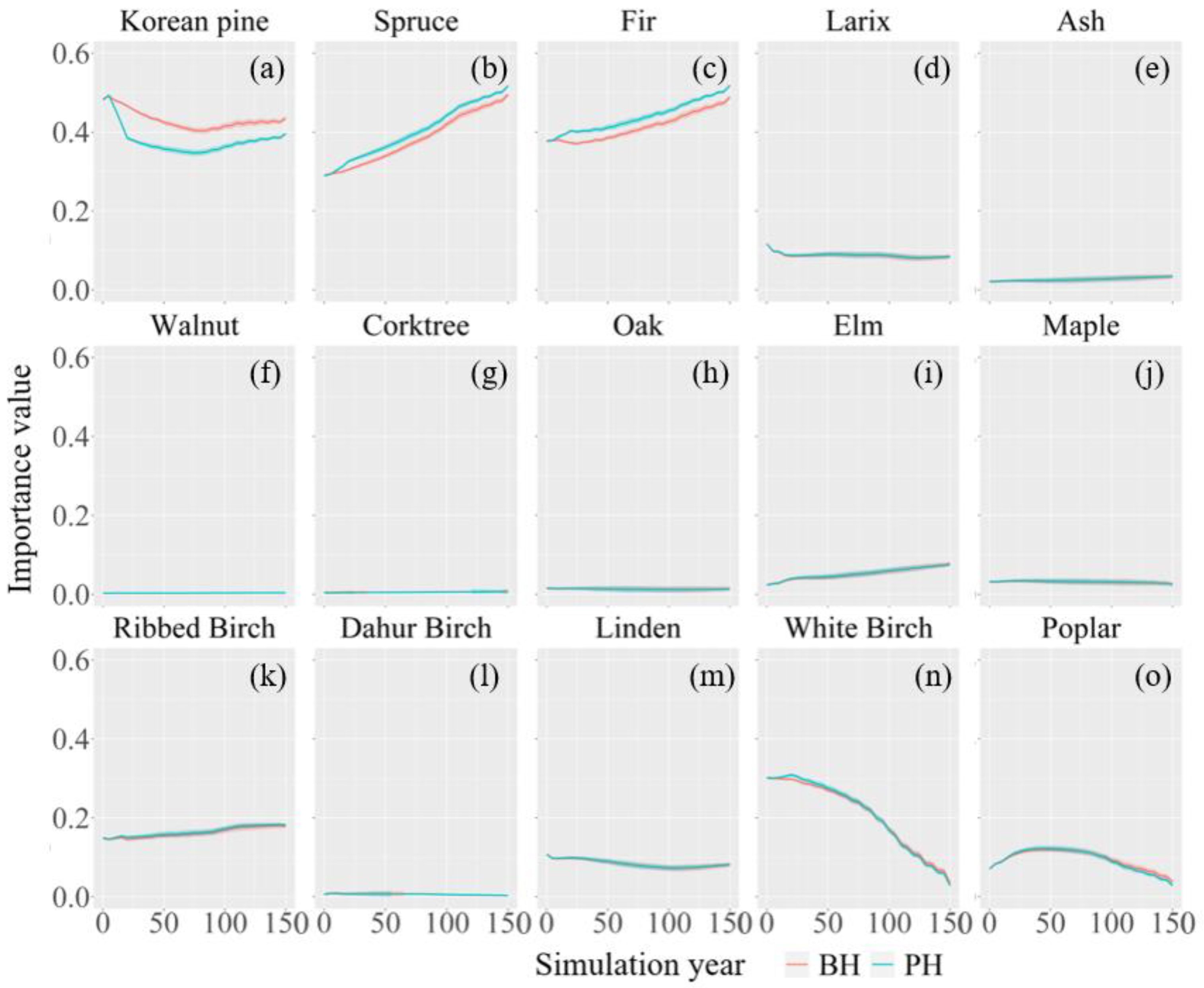
3.2. Change in Species Composition
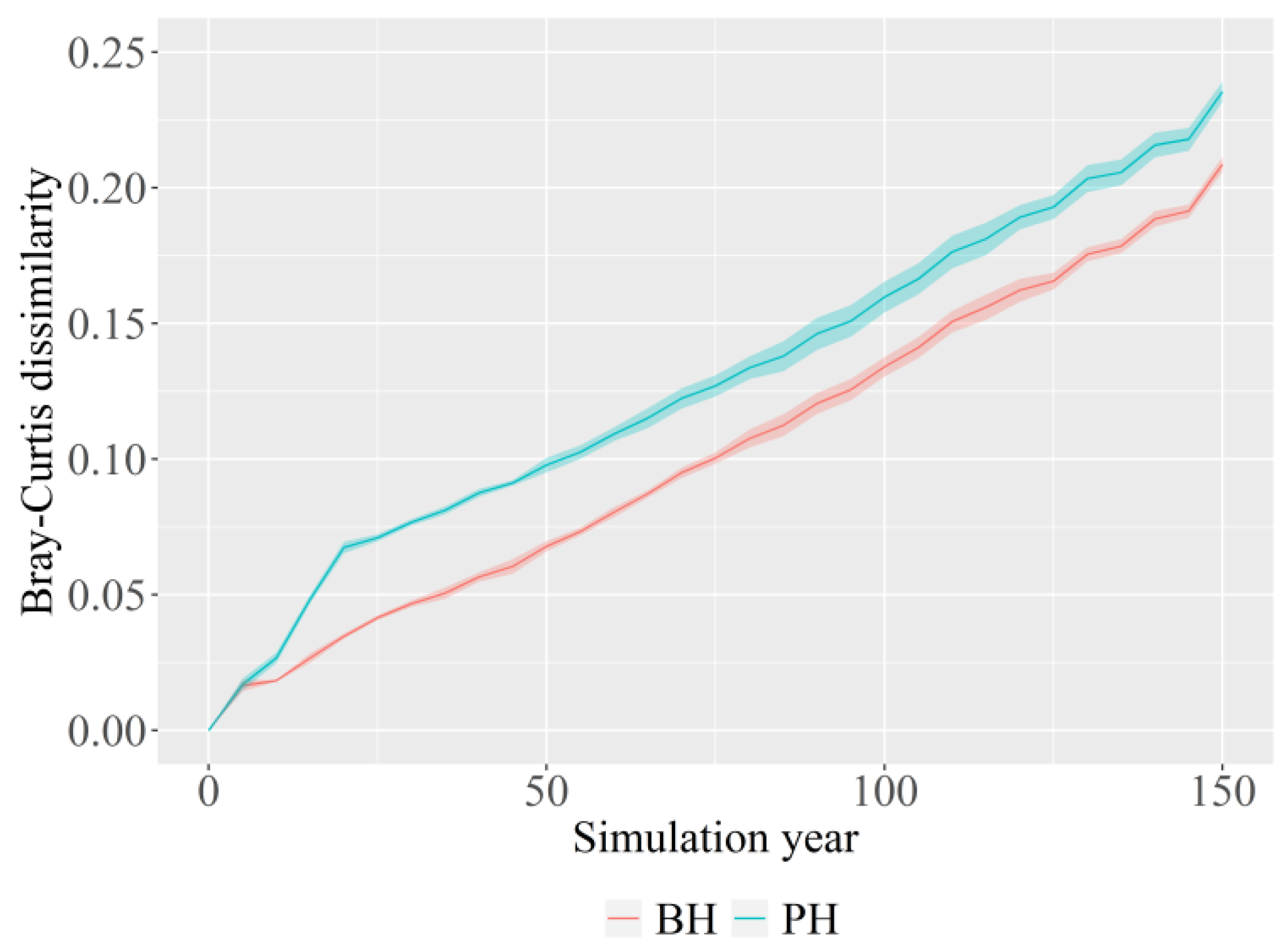
3.3. Change in Succession Trajectory
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, D.; Zhou, W.; Zhou, L.; Dai, L. Exploring the history of the management theory and technology of broad—Leaved Korean pine (Pinus koraiensis Sieb. et Zucc.) forest in Changbai Mountain Region, Northeast China. Chin. J. Appl. Ecol. 2019, 30, 1426–1434. [Google Scholar]
- Zheng, X.; Yu, D.; Wang, A.; Dai, G. Long-term Monitoring and Field Control Experiments Reveal the Structure, Function and Evolution of Typical Temperate Forests. Bull. Chin. Acad. Sci. 2019, 34, 948–961. [Google Scholar]
- Liu, K.; Liang, Y.; He, H.S.; Wang, W.J.; Huang, C.; Zong, S.; Wang, L.; Xiao, J.; Du, H. Long-term impacts of China’s new commercial harvest exclusion policy on ecosystem services and biodiversity in the temperate forests of Northeast China. Sustainability 2018, 10, 1071. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; He, H.; Dai, L.; Yang, J. Effects of human disturbances on Korean pine coverage and age structure at a landscape scale in Northeast China. Ecol. Eng. 2014, 71, 375–379. [Google Scholar] [CrossRef]
- Dai, L.; Li, S.; Zhou, W.; Qi, L.; Zhou, L.; Wei, Y.; Li, J.; Shao, G.; Yu, D. Opportunities and challenges for the protection and ecological functions promotion of natural forests in China. For. Ecol. Manag. 2018, 410, 187–192. [Google Scholar] [CrossRef]
- Ji, L.; Liu, Z.; Hao, Z.; Wang, Q.; Wang, M. Effect of Cones Picking on Broad-leaved Pinus koraiensis Forest in Changbai Mountian. Chin. J. Ecol. 2002, 21, 39–42. [Google Scholar]
- Yu, D.; Zhou, W.; Bao, Y.; Qi, L.; Zhou, L.; Dai, L. Forest management of Korean pine and broadleaf mixed forest in Northeast China since the implementation of Natural Forest Protection Project. Acta Ecol. Sin. 2015, 35, 10–17. [Google Scholar]
- Zhou, G.; Liu, Q.; Xu, Z.; Du, W.; Yu, J.; Meng, S.; Zhou, H.; Qin, L.; Shah, S. How can the shade intolerant Korean pine survive under dense deciduous canopy? For. Ecol. Manag. 2020, 457, 117735. [Google Scholar] [CrossRef]
- Liu, Z.; Ji, L.; Hao, Z.; Zhu, J.; Kang, H. Effect of cone-picking on natural regeneration of Korean pine in Changbai Mountain Nature Reserve. Chin. J. Appl. Ecol. 2004, 15, 958–962. [Google Scholar]
- Piao, Z.; Tang, L.; Swihart, R.K.; Wang, S. Human–wildlife competition for Korean pine seeds: Vertebrate responses and implications for mixed forests on Changbai Mountain, China. Ann. For. Sci. 2011, 68, 911–919. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-B.; Mou, P.; Wang, T.-M.; Ge, J. Evaluation of regeneration potential of Pinus koraiensis in mixed pine-hardwood forests in the Xiao Xing’an Mountains, China. J. For. Res. 2012, 23, 543–551. [Google Scholar] [CrossRef]
- Jin, G.; Yang, G.; Ma, J.; Li, L.; Xu, Z.; Zhao, X.; Hong, M. Effect of Anthropogenic Cone-picking on Seed Bank and Seedling Bank of Korean Pine in the Major Forest Types in Lesser Hing’an Mountains. J. Nat. Resour. 2010, 25, 1845–1854. [Google Scholar]
- Li, J.; Li, J. Regeneration and restoration of broad-leaved Korean pine forests in Lesser Xing′an Mountains of Northeast China. Acta Ecol. Sin. 2003, 23, 1268–1277. [Google Scholar]
- Wang, J.; Geoff Wang, G.; Li, R.; Sun, Y.; Yu, L.; Zhu, J.; Yan, Q. Cascading effect of source limitation on the granivore-mediated seed dispersal of Korean pine (Pinus koraiensis) in secondary forest ecosystems. Ecol. Process. 2022, 11, 1. [Google Scholar] [CrossRef]
- Zhou, Y. Vegetation in the Small Khingan Mountains of China; Science Press: Beijing, China, 1994. [Google Scholar]
- Yu, Z.; Zhao, S.; Wang, Q.; Zhai, Y. Simulation of polar-birch forest succession in broad-leaved-Korean pine forest zone in Mt. Changbai. Chin. J. Appl. Ecol. 1997, 8, 455–458. [Google Scholar]
- He, H.S.; Hao, Z.; Mladenoff, D.J.; Shao, G.; Hu, Y.; Chang, Y. Simulating forest ecosystem response to climate warming incorporating spatial effects in north-eastern China. J. Biogeogr. 2005, 32, 20432056. [Google Scholar] [CrossRef]
- He, H.S.; Gustafson, E.J.; Lischke, H. Modeling forest landscapes in a changing climate: Theory and application. Landscape Ecol. 2017, 32, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Shifley, S.R.; He, H.S.; Lischke, H.; Wang, W.J.; Jin, W.; Gustafson, E.J.; Thompson, J.R.; Thompson, F.R.; Dijak, W.D.; Yang, J. The past and future of modeling forest dynamics: From growth and yield curves to forest landscape models. Landsc. Ecol. 2017, 32, 1307–1325. [Google Scholar] [CrossRef]
- Liu, K.; He, H.; Xu, W.; Du, H.; Zong, S.; Huang, C.; Wu, M.; Tan, X.; Cong, Y. Responses of korean pine to proactive managements under climate change. Forests 2020, 11, 263. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; He, H.S.; Spetich, M.A.; Shifley, S.R.; Thompson, F.R., III; Dijak, W.D.; Wang, Q. A framework for evaluating forest landscape model predictions using empirical data and knowledge. Environ. Modell. Softw. 2014, 62, 230–239. [Google Scholar] [CrossRef]
- He, H.S.; Mladenoff, D.J. The effects of seed dispersal on the simulation of long-term forest landscape change. Ecosystems 1999, 2, 308–319. [Google Scholar] [CrossRef]
- Yoda, K. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants. XI). J. Inst. Polytech. Osaka City Univ. Ser. D. 1963, 14, 107–129. [Google Scholar]
- Wang, W.J.; He, H.S.; Fraser, F.R.T.J.S., III; Hanberry, B.B.; Dijak, W.D. Importance of succession, harvest, and climate change in determining future composition in US Central Hardwood Forests. Ecosphere 2015, 6, 277. [Google Scholar] [CrossRef]
- Xiao, J.; Liang, Y.; He, H.S.; Thompson, J.R.; Wang, W.J.; Fraser, J.S.; Wu, Z. The formulations of site-scale processes affect landscape-scale forest change predictions: A comparison between LANDIS PRO and LANDIS-II forest landscape models. Landsc. Ecol. 2017, 32, 1347–1363. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing (Version 3.2.2); R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Zhao, H.; Kang, X.; Guo, Z.; Yang, H.; Xu, M. Species interactions in spruce–fir mixed stands and implications for enrichment planting in the Changbai Mountains, China. Mt. Res. Dev. 2012, 32, 187–196. [Google Scholar] [CrossRef]
- Shao, G.; Schall, P.; Weishampel, J.F. Dynamic simulations of mixed broadleaved-Pinus koraiensis forests in the Changbaishan Biosphere Reserve of China. For. Ecol. Manag. 1994, 70, 169–181. [Google Scholar] [CrossRef]
- De Oliveira Wadt, L.H.; Faustino, C.L.; Staudhammer, C.L.; Kainer, K.A.; Evangelista, J.S. Primary and secondary dispersal of Bertholletia excelsa: Implications for sustainable harvests. For. Ecol. Manag. 2018, 415, 98–105. [Google Scholar] [CrossRef]
- Wang, J.; Wang, G.G.; Zhang, T.; Yuan, J.; Yu, L.; Zhu, J.; Yan, Q. Use of direct seeding and seedling planting to restore Korean pine (Pinus koraiensis Sieb. Et Zucc.) in secondary forests of Northeast China. For. Ecol. Manag. 2021, 493, 119243. [Google Scholar] [CrossRef]
- Freitas, M.G.; Rodrigues, S.B.; Campos-Filho, E.M.; do Carmo, G.H.P.; da Veiga, J.M.; Junqueira, R.G.P.; Vieira, D.L.M. Evaluating the success of direct seeding for tropical forest restoration over ten years. For. Ecol. Manag. 2019, 438, 224–232. [Google Scholar] [CrossRef]
- Raupp, P.P.; Ferreira, M.C.; Alves, M.; Campos-Filho, E.M.; Sartorelli, P.A.R.; Consolaro, H.N.; Vieira, D.L.M. Direct seeding reduces the costs of tree planting for forest and savanna restoration. Ecol. Eng. 2020, 148, 105788. [Google Scholar] [CrossRef]
- Atondo-Bueno, E.J.; Bonilla-Moheno, M.; López-Barrera, F. Cost-efficiency analysis of seedling introduction vs. direct seeding of Oreomunnea mexicana for secondary forest enrichment. For. Ecol. Manag. 2018, 409, 399–406. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | MT | LG | ST | MD | MDBH | MSDI | NPGS |
|---|---|---|---|---|---|---|---|
| Korean pine, Pinus koraiensis | 40 | 300 | 4 | 150 | 110 | 550 | 20 |
| Korean spruce, Picea koraiensis and Picea jezoensis | 30 | 300 | 4 | 150 | 90 | 600 | 20 |
| Khingan fir, Abies nephrolepis | 30 | 300 | 4 | 150 | 85 | 650 | 20 |
| Larch, Larix gmelinii | 20 | 300 | 2 | 300 | 95 | 650 | 30 |
| Manchurian ash, Fraxinus mandschurica | 30 | 250 | 3 | 300 | 100 | 600 | 25 |
| Manchurian walnut, Juglans mandshurica | 20 | 250 | 2 | 200 | 90 | 650 | 25 |
| Amur corktree, Phellodendron amurense | 20 | 250 | 3 | 300 | 95 | 650 | 25 |
| Mongolian oak, Quercus mongolica | 20 | 300 | 2 | 200 | 95 | 600 | 20 |
| Black elm, Ulmus davidiana | 20 | 250 | 3 | 800 | 90 | 600 | 25 |
| Mono maple, Acer mono | 20 | 200 | 3 | 200 | 60 | 700 | 25 |
| Ribbed birch, Betula costata | 20 | 250 | 3 | 800 | 90 | 650 | 25 |
| Dahur Birch, Betula dahurica | 15 | 150 | 2 | 800 | 50 | 750 | 25 |
| Amur linden, Tilia amurensis | 30 | 300 | 3 | 200 | 85 | 650 | 20 |
| White birch, Betula platyphylla | 15 | 150 | 1 | 2000 | 50 | 800 | 30 |
| Poplar, Populus davidiana | 15 | 150 | 1 | 2000 | 60 | 800 | 30 |
| Scenarios | Removal Method | Percent Area for Background Harvesting | Percent Area for Historical Predatory Seed Harvesting |
|---|---|---|---|
| Baseline harvest (BH) | Thinning from below | 5% | — |
| Predatory harvest (PH) | Thinning from below | 5% | 80% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; He, H.S.; Sun, H.; Wang, J. Evaluating the Legacy Effects of the Historical Predatory Seed Harvesting on the Species Composition and Structure of the Mixed Korean Pine and Broadleaf Forest from a Landscape Perspective. Forests 2023, 14, 402. https://doi.org/10.3390/f14020402
Liu K, He HS, Sun H, Wang J. Evaluating the Legacy Effects of the Historical Predatory Seed Harvesting on the Species Composition and Structure of the Mixed Korean Pine and Broadleaf Forest from a Landscape Perspective. Forests. 2023; 14(2):402. https://doi.org/10.3390/f14020402
Chicago/Turabian StyleLiu, Kai, Hong S. He, Hang Sun, and Jingfeng Wang. 2023. "Evaluating the Legacy Effects of the Historical Predatory Seed Harvesting on the Species Composition and Structure of the Mixed Korean Pine and Broadleaf Forest from a Landscape Perspective" Forests 14, no. 2: 402. https://doi.org/10.3390/f14020402