
Effects of Polyploidy on Physiological Performance of Acclimatized Solanum betaceum Cav. Plants under Water Deficit
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of Shoot Cultures
2.2. Plant Material and Experimental Design
2.3. Growth and Plant Water Potential
2.4. Leaf gas-Exchange Measurements
2.5. Photosynthetic Pigments and Chlorophyll a Fluorescence Analysis
2.6. Lipid Peroxidation and Proline Content
2.7. Total Soluble Sugars and Starch
2.8. Statistical Analysis
3. Results
3.1. Acclimatization of Micropropagated Plants
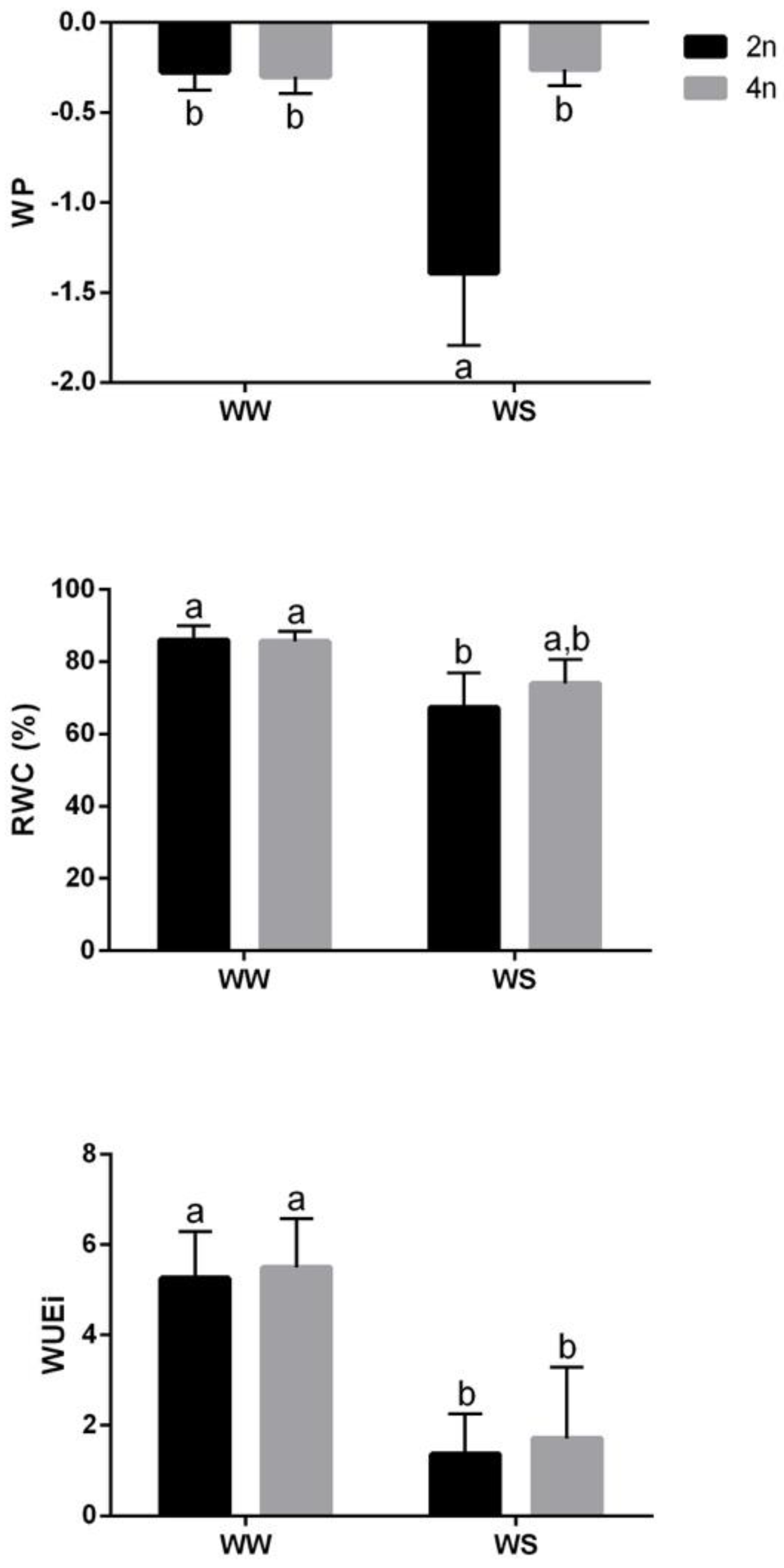
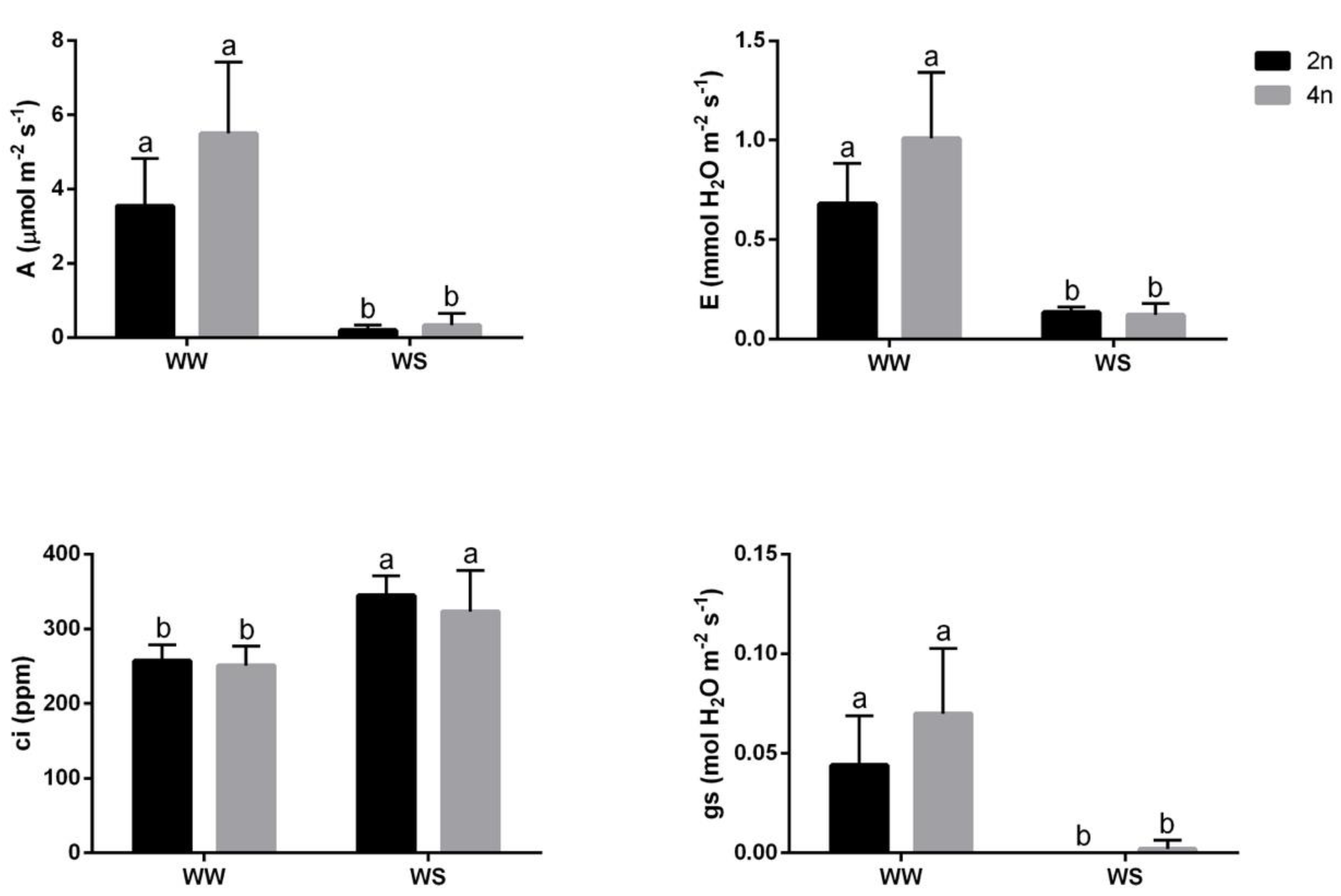
3.2. Responses of Diploid and Tetraploid Plants under Water Deficit
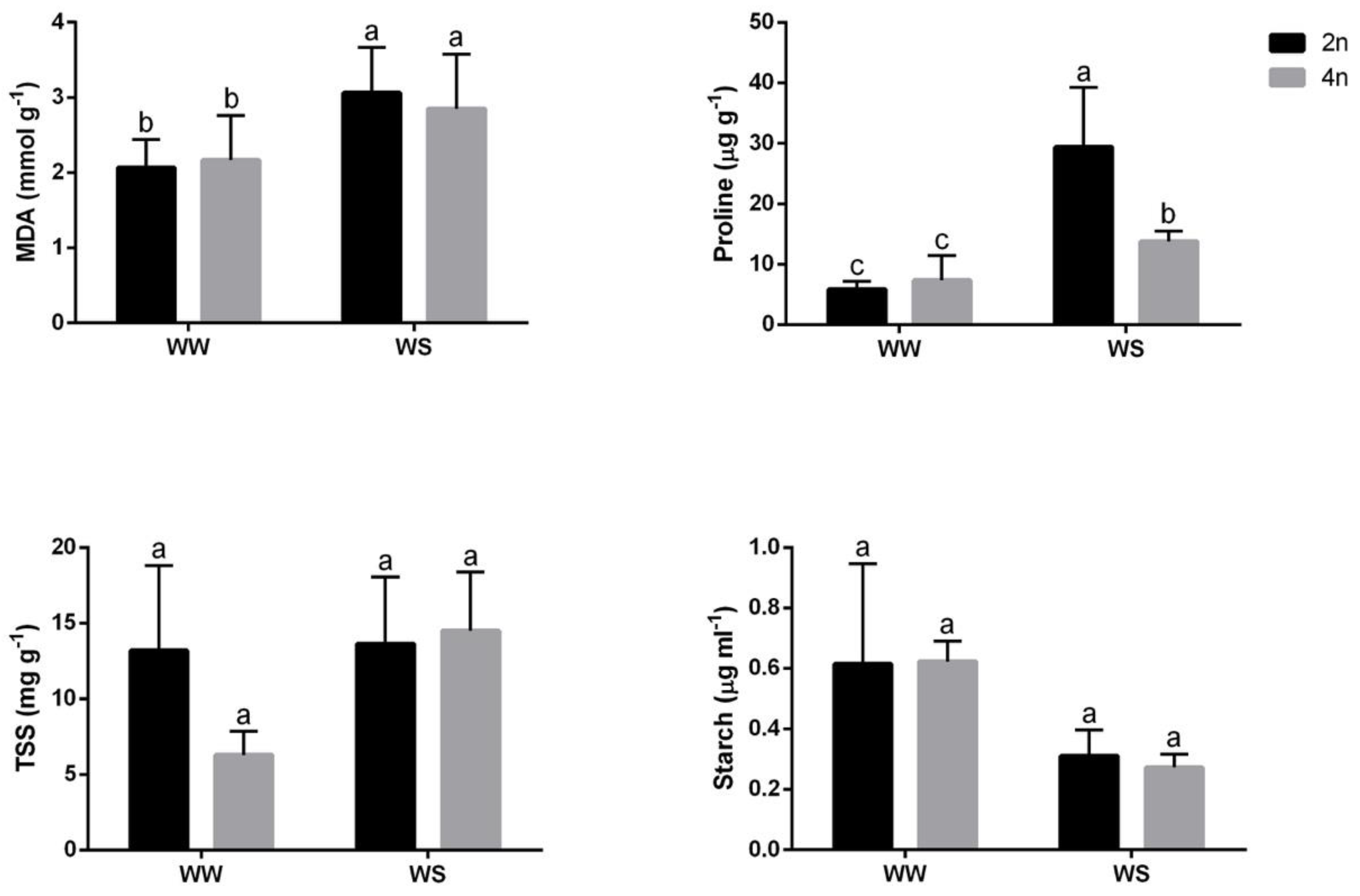
3.3. Changes in Proline, MDA and Carbohydrates Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meadows, L.R. Growing Tamarillo Relatives. The New Zealand Home Garden. 2002. Available online: www.naturalhub.com (accessed on 18 May 2021).
- Mertz, C.; Brat, P.; Caris-Veyrat, C.; Günata, Z. Characterization and thermal lability of carotenoids and vitamin C of tamarillo fruit (Solanum betaceum Cav.). Food Chem. 2009, 119, 653–659. [Google Scholar] [CrossRef]
- Noor Atiqah, A.A.K.; Maisarah, A.M.; Asmah, R. Comparison of antioxidant properties of tamarillo (Cyphomandra betacea), cherry tomato (Solanum lycopersicum var. cerasiforme) and tomato (Lycopersicon esculentum). Int. Food Res. J. 2014, 21, 2355–2362. [Google Scholar]
- Osorio, S.; Alba, R.; Nikoloski, Z.; Kochevenko, A.; Fernie, A.R.; Giovannoni, J.J. Integrative comparative analyses of transcript and metabolite profiles from pepper and tomato ripening and development stages uncovers species-specific patterns of network regulatory behavior. Plant Physiol. 2012, 159, 1713–1729. [Google Scholar] [CrossRef] [Green Version]
- Cohen, D.; Elliot, D. Micropropagation methods for blueberries and tamarillos. Comb. Proc. Int. Plant Prop. Soc. 1979, 29, 177–179. [Google Scholar]
- Bakshi, P.; Kour, G.; Ahmed, R. Tamarillo (Cyphomandra betacea). In Underutilized Fruit Crops: Importance and Cultivation, 1st ed.; Ghosh, S.N., Ed.; Jaya Publications: Delhi, India, 2016; pp. 1271–1294. [Google Scholar]
- Correia, S.; Canhoto, J.M. Biotechnology of tamarillo (Cyphomandra betacea): From in vitro cloning to genetic transformation. Sci. Hortic. 2012, 148, 161–168. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Balting, D.F.; AghaKouchak, A.; Lohmann, G.; Ionita, M. Northern Hemisphere drought risk in a warming climate. Npj Clim. Atmos. Sci. 2021, 4, 61. [Google Scholar] [CrossRef]
- Trojak-Goluch, A.; Kawka-Lipińska, M.; Wielgusz, K.; Praczyk, M. Polyploidy in industrial crops: Applications and perspectives in plant breeding. Agronomy 2021, 11, 2574. [Google Scholar] [CrossRef]
- Leitch, A.R.; Leitch, I.J. Genomic plasticity and the diversity of polyploid plants. Science 2008, 320, 481–483. [Google Scholar] [CrossRef]
- Ntuli, N.; Zobolo, A.M. Effect of water stress on growth of colchicine induced polyploid Coccinia palmata and Lagenaria sphaerica plants. Afr. J. Biotechnol. 2008, 7, 3548–3652. [Google Scholar]
- Liu, S.; Chen, S.; Chen, Y.; Guan, Z.; Yin, D.; Chen, F. In vitro induced tetraploid of Dendranthema nankingense (Nakai) Tzvel. shows an improved level of abiotic stress tolerance. Sci. Hortic. 2011, 127, 411–419. [Google Scholar] [CrossRef]
- Radia, L.; Julie, O.; Yann, Q.; Yann, F.; Stéphane, H.; Raphael, M.; Liliane, B.; Jérémie, S. Improved response of triploid citrus varieties to water deficit is related to anatomical and cytological properties. Plant Physiol. Biochem. 2021, 162, 762–775. [Google Scholar]
- del Pozo, J.C.; Ramirez-Parra, E. Deciphering the molecular bases for drought tolerance in Arabidopsis autotetraploids. Plant Cell Environ. 2014, 37, 2722–2737. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.M.; Huang, Q.C.; Qin, G.Y.; Zhao, S.P.; Zhou, J.G. Different drought-stress responses in photosynthesis and reactive oxygen metabolism between autotetraploid and diploid rice. Photosynthetica 2014, 52, 193–202. [Google Scholar] [CrossRef]
- Pringle, G.; Murray, B. Polyploidy and aneuploidy in the tamarillo, Cyphomandra betacea (Cav.) Sendt. (Solanaceae). I. Spontaneous polyploidy and features of the euploids. Plant Breed. 1992, 108, 132–138. [Google Scholar] [CrossRef]
- Antunes, P. Indução de plantas tetraplóides através de tratamento com agentes c-mitóticos no tamarilho (Cyphomandra betacea) e no medronheiro (Arbutus unedo). Master’s Thesis, University of Coimbra, Coimbra, Portugal, 2010. [Google Scholar]
- Tossi, V.E.; Martínez Tosar, L.J.; Laino, L.E.; Iannicelli, J.; Regalado, J.J.; Escandón, A.S.; Baroli, I.; Causin, H.F.; Pitta-Álvarez, S.I. Impact of polyploidy on plant tolerance to abiotic and biotic stresses. Front. Plant Sci. 2022, 13, 869423. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Correia, B.; Pintó-Marijuan, M.; Neves, L.; Brossac, R.; Dias, M.C.; Costa, A.; Castro, B.; Araújo, C.; Santosa, C.; Chaves, M.M.; et al. Water stress and recovery in the performance of two Eucalyptus globulus clones: Physiological and biochemical profiles. Physiol. Plant. 2014, 150, 580–592. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Hodges, J.M. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Khedr, A.; Abbas, M.; Wahid, A.; Quick, W.; Abogadallah, G. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J. Exp. Bot. 2003, 54, 2553–2562. [Google Scholar] [CrossRef] [Green Version]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Mitsuru, O.; Takuro, S.; Toshiaki, T. Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci. Plant Nutr. 1991, 37, 117–128. [Google Scholar]
- Van de Peer, Y.; Ashman, T.-L.; Soltis, P.S.; Soltis, D.E. Polyploidy: An evolutionary and ecological force in stressful times. Plant Cell 2021, 33, 11–26. [Google Scholar] [CrossRef]
- Maherali, H.; Walden, A.E.; Husband, B.C. Genome duplication and the evolution of physiological responses to water stress. New Phytol. 2009, 184, 721–731. [Google Scholar] [CrossRef]
- Li, W.-D.; Biswas, D.K.; Xu, H.; Xu, C.-Q.; Wang, X.-Z.; Liu, J.-K.; Jiang, G.-M. Photosynthetic responses to chromosome doubling in relation to leaf anatomy in Lonicera japonica subjected to water stress. Funct. Plant Biol. 2009, 36, 783–792. [Google Scholar] [CrossRef]
- Acquaah, G. Principles of plant genetics and breeding. In Polyploidy in Plant Breeding; Wiley-Blackwell: Malden, MA, USA, 2007; Volume 13, pp. 130–214. [Google Scholar]
- Song, C.; Liu, S.; Xiao, J.; Zhou, Y.; Qin, Q.; Zhang, C.; Liu, Y. Polyploid organisms. Science China Life. Science 2012, 55, 301–311. [Google Scholar]
- Huang, M.; Deng, X.; Zhao, Y.; Zhou, S.; Inanaga, S.; Yamada, S.; Tanaka, K. Water and nutrient use efficiency in diploid, tetraploid and hexaploid wheats. J. Integr. Plant Biol. 2007, 49, 706–715. [Google Scholar] [CrossRef]
- Souza, R.; Machado, E.; Silva, J.; Lagoa, A.; Silveira, J. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Correia, M.; Osório, M.; Osório, J.; Barrote, I.; Martins, M.; David, M. Influence of transient shade periods on the effects of drought on photosynthesis, carbohydrate accumulation and lipid peroxidation in sunflower leaves. Environ. Exp. Bot. 2006, 58, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought–from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Bio Technol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Fàbregas, N.; Fernie, A.R. The metabolic response to drought. J. Exp. Bot. 2019, 70, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Rosa, M.; Prado, C.; Podazza, G.; Roque, I.; Juan, A.G.; Mirna, H.; Prado, F.E. Soluble sugars--metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Pavlík, M.; Pavlíková, D.; Staszková, L.; Neuberg, M.; Kaliszová, R. The effect of arsenic contamination on aminoacids metabolism in Spinacia oleracea L. Ecotoxicol. Environ. Saf. 2010, 73, 1309–1313. [Google Scholar] [CrossRef]
- Živanović, B.; Milić Komić, S.; Tosti, T.; Vidović, M.; Prokić, L.; Veljović Jovanović, S. Leaf soluble sugars and free amino acids as important components of abscisic acid—Mediated drought response in tomato. Plants 2020, 9, 1147. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, J.; Chen, L.; Yang, D.; Shen, J.; Li, J.; Liston, A.; Ashman, T.-L.; Dong, M. ABA-regulated ploidy-related genes and non-structural carbon accumulation may underlie cold tolerance in tetraploid Fragaria moupinensis. Environ. Exp. Bot. 2020, 179, 104232. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of sweet pepper with stress tolerance-inducing compounds alleviates salinity stress oxidative damage by mediating the physio-biochemical activities and antioxidant systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Ploidy | Total Biomass (DW, g) | Root Lenght (cm) | Shoot Lenght (cm) | Plant Height (cm) |
|---|---|---|---|---|---|
| WW | 2n | 5.15 ± 1.18 a | 23.63 ± 4.44 a | 23.00 ± 2.63 a | 46.63 ± 5.34 a |
| 4n | 4.77 ± 1.18 a | 20.82 ± 1.27 a | 18.58 ± 1.17 b | 39.40 ± 1.10 b | |
| WS | 2n | 5.48 ± 1.12 a | 20.37 ± 0.84 a | 20.50 ± 1.51 a,b | 40.87 ± 1.63 b |
| 4n | 4.12 ± 0.84 a | 21.73 ± 2.24 a | 18.50 ± 0.96 b | 40.23 ± 2.46 b |
| Treatment | Ploidy | Chlorophyll a Fluorescence | Pigment Content (µmol mL−1) | |||
|---|---|---|---|---|---|---|
| ϕPSII | Fv/Fm | Chl a | Chl b | Car | ||
| WW | 2n | 0.69 ± 0.02 a | 0.80 ± 0.01 a | 0.0095 ± 0.0015 a | 0.0033 ± 0.0006 a | 0.0043 ± 0.0007 a |
| 4n | 0.58 ± 0.10 a | 0.82 ± 0.01 a | 0.0093 ± 0.0013 a | 0.0033 ± 0.0004 a | 0.0042 ± 0.0005 a | |
| WS | 2n | 0.49 ± 0.10 b | 0.79 ± 0.01 a | 0.0102 ± 0.0008 a | 0.0035 ± 0.0003 a | 0.0049 ± 0.0004 a |
| 4n | 0.46 ± 0.09 b | 0.80 ± 0.02 a | 0.0084 ± 0.0004 a | 0.0030 ± 0.0002 a | 0.0041 ± 0.0004 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, S.; Braga, A.; Martins, J.; Correia, B.; Pinto, G.; Canhoto, J. Effects of Polyploidy on Physiological Performance of Acclimatized Solanum betaceum Cav. Plants under Water Deficit. Forests 2023, 14, 208. https://doi.org/10.3390/f14020208
Correia S, Braga A, Martins J, Correia B, Pinto G, Canhoto J. Effects of Polyploidy on Physiological Performance of Acclimatized Solanum betaceum Cav. Plants under Water Deficit. Forests. 2023; 14(2):208. https://doi.org/10.3390/f14020208
Chicago/Turabian StyleCorreia, Sandra, Ana Braga, João Martins, Barbara Correia, Glória Pinto, and Jorge Canhoto. 2023. "Effects of Polyploidy on Physiological Performance of Acclimatized Solanum betaceum Cav. Plants under Water Deficit" Forests 14, no. 2: 208. https://doi.org/10.3390/f14020208