
Climate–Growth Relationships in Laurus azorica—A Dominant Tree in the Azorean Laurel Forest
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
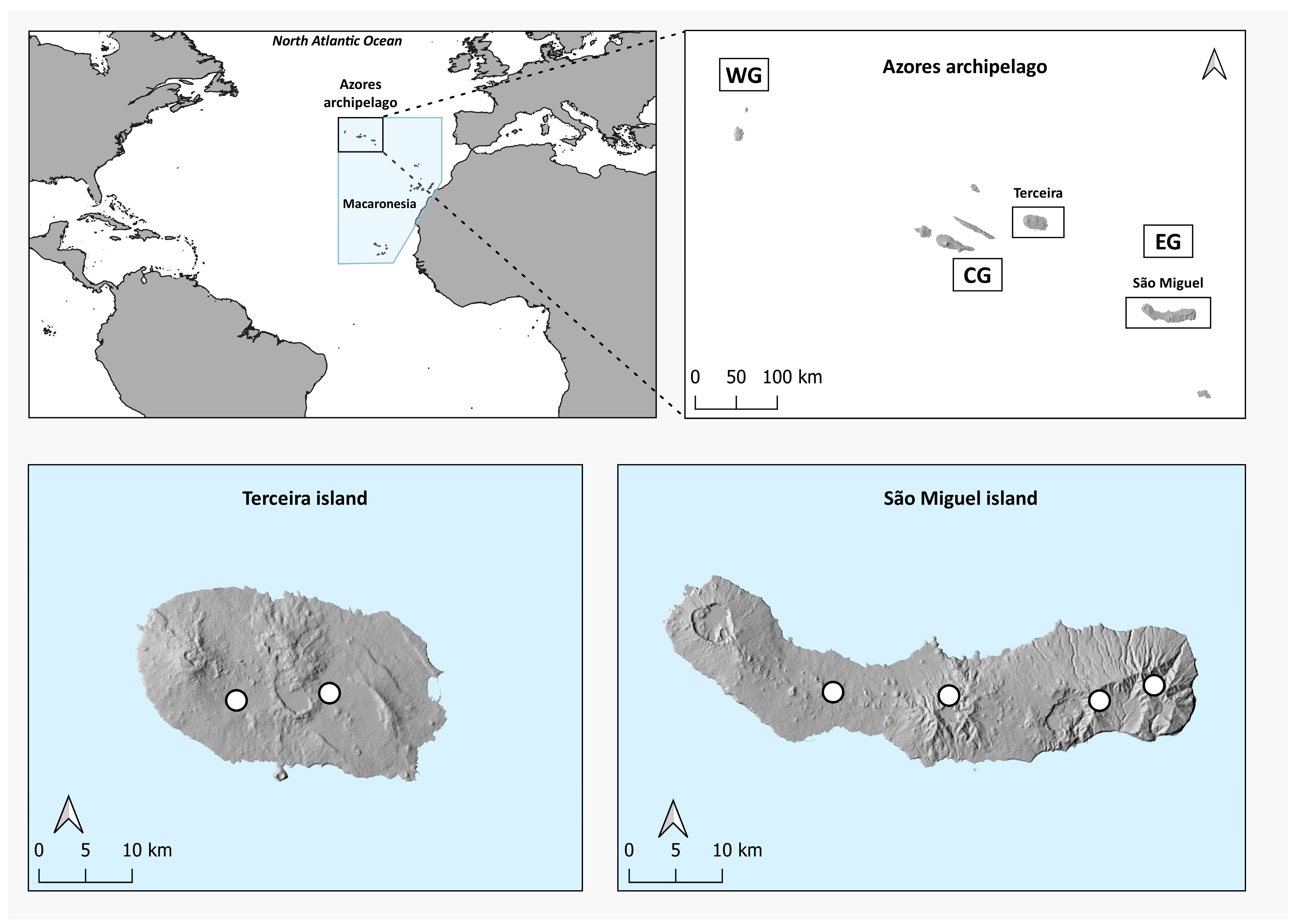
2.1. Study Area
2.2. Target Species—Laurus azorica
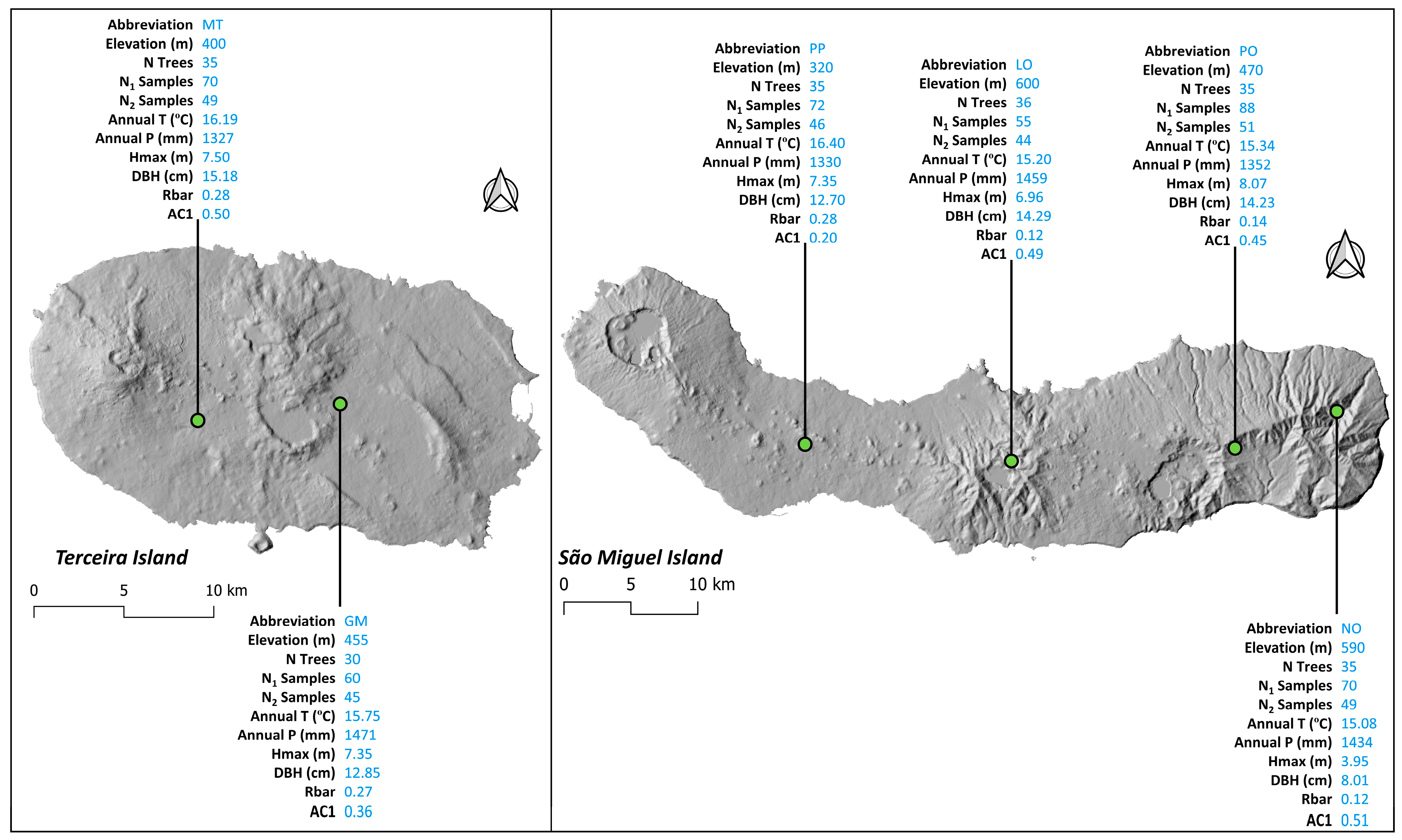
2.3. Sample Collection and Data Preparation
2.4. Tree-Ring Data Collection and Analysis
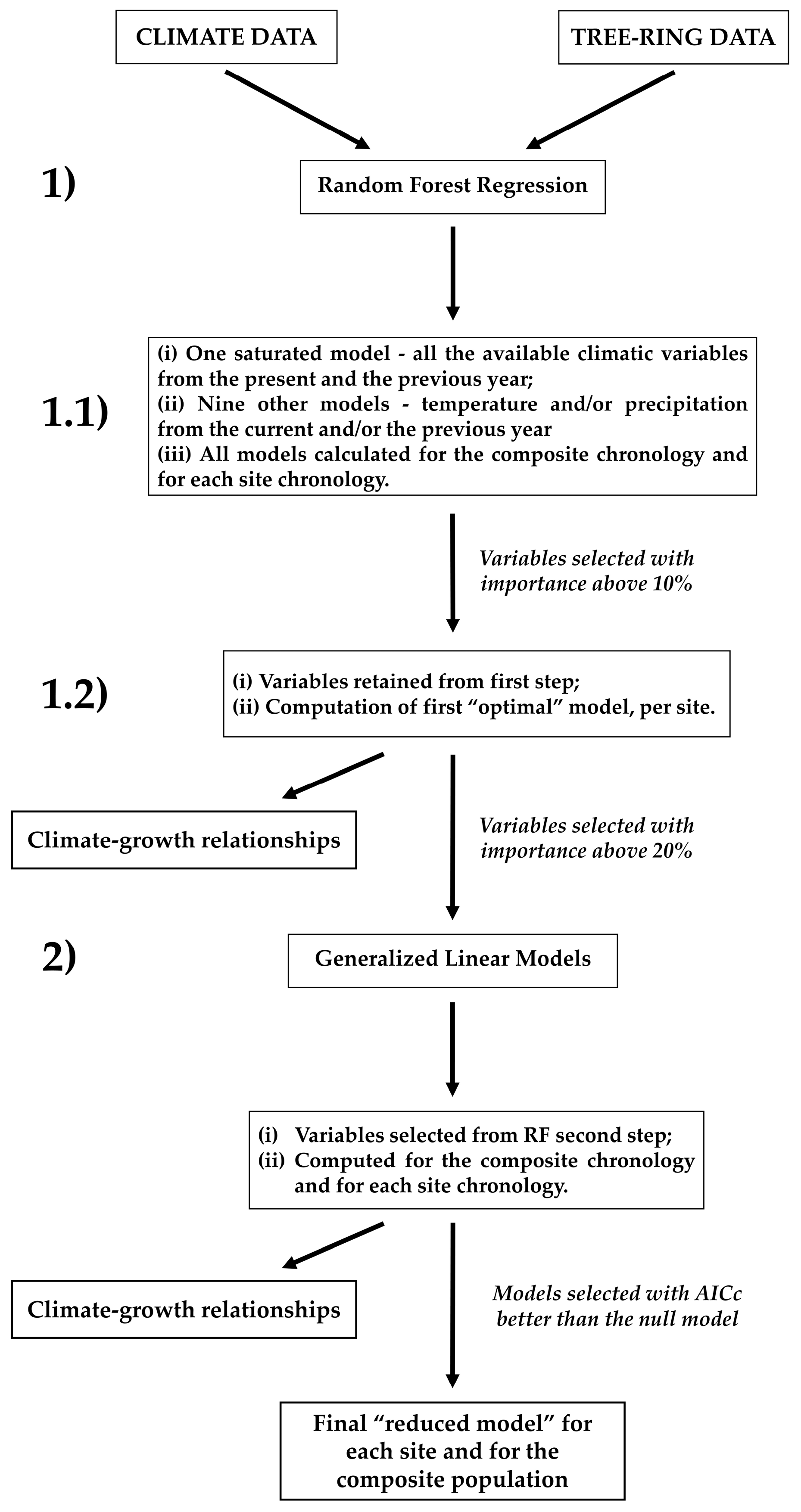
2.5. Dendroclimatic Models
2.5.1. Climatic Variables
2.5.2. Modeling Approaches
2.5.3. Model Implementation and Variable Selection
3. Results
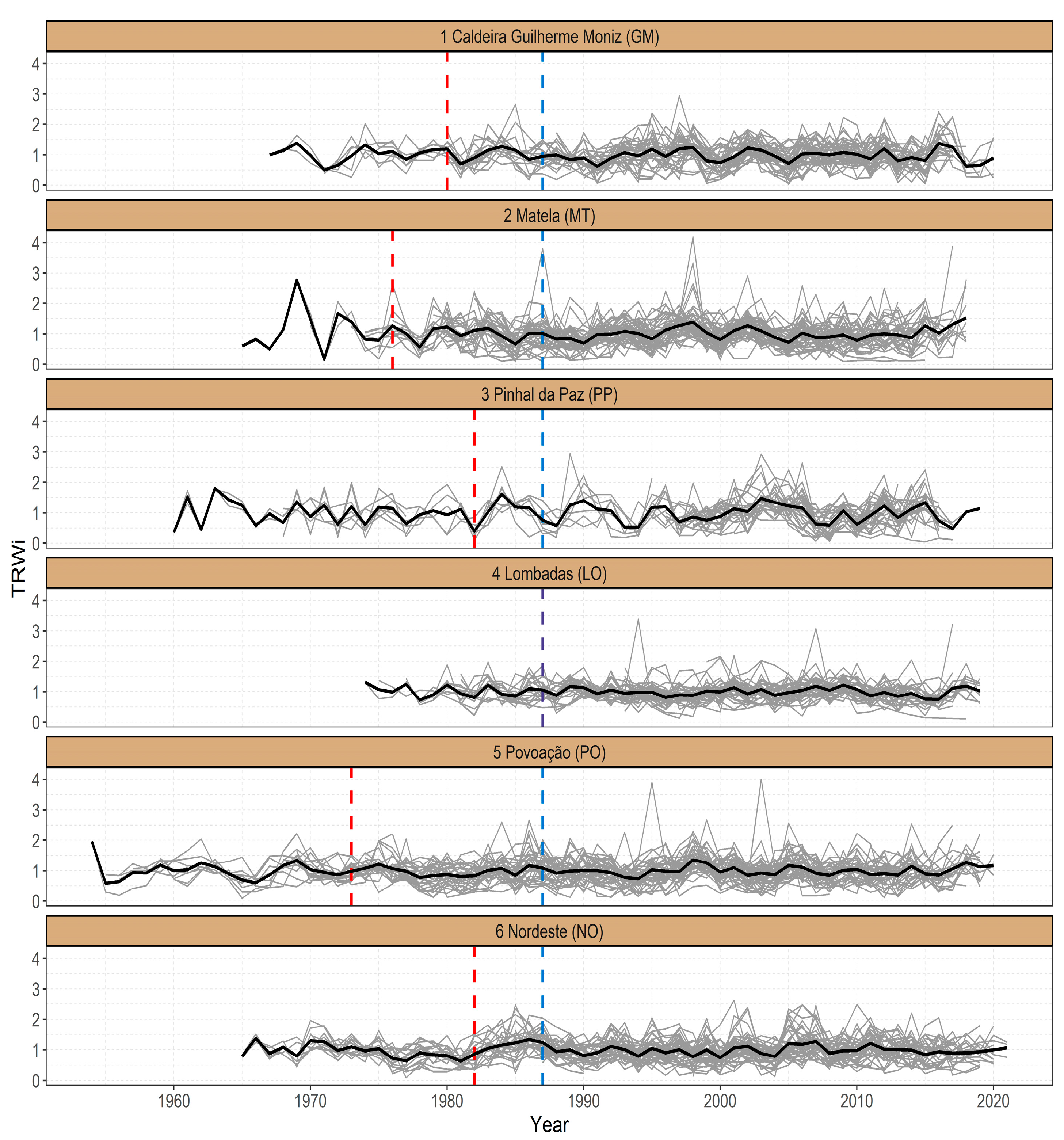
3.1. Site Chronologies
3.2. Climate–Growth Relationships
4. Discussion
4.1. Chronology Evaluation
4.2. Modeling Constraints
4.3. Climate–Growth Relationships—Temperature Effect
4.4. Climate–Growth Relationships—Precipitation Effect
4.5. Climate–Growth Relationships—Previous Year Effect
4.6. Climate–Growth Relationships—Differences among Sites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elias, R.B.; Gil, A.; Silva, L.; Fernández-Palacios, J.M.; Azevedo, E.B.; Reis, F. Natural Zonal Vegetation of the Azores Islands: Characterization and Potential Distribution. Phytocoenologia 2016, 46, 107–123. [Google Scholar] [CrossRef]
- Pavão, D.C.; Elias, R.B.; Silva, L. Comparison of Discrete and Continuum Community Models: Insights from Numerical Ecology and Bayesian Methods Applied to Azorean Plant Communities. Ecol. Modell. 2019, 402, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Mueller-Dombois, D.; Boehmer, H.J. Origin of the Hawaiian Rainforest and Its Transition States in Long-Term Primary Succession. Biogeosciences 2013, 10, 5171–5182. [Google Scholar] [CrossRef] [Green Version]
- Hanz, D.M.; Cutts, V.; Barajas-Barbosa, M.P.; Algar, A.C.; Beierkuhnlein, C.; Fernández-Palacios, J.M.; Field, R.; Kreft, H.; Steinbauer, M.J.; Weigelt, P.; et al. Climatic and Biogeographical Drivers of Functional Diversity in the Flora of the Canary Islands. Glob. Ecol. Biogeogr. 2022, 31, 1313–1331. [Google Scholar] [CrossRef]
- Kier, G.; Kreft, H.; Tien, M.L.; Jetz, W.; Ibisch, P.L.; Nowicki, C.; Mutke, J.; Barthlott, W. A Global Assessment of Endemism and Species Richness across Island and Mainland Regions. Proc. Natl. Acad. Sci. USA 2009, 106, 9322–9327. [Google Scholar] [CrossRef] [Green Version]
- Weigelt, P.; Jetz, W.; Kreft, H. Bioclimatic and Physical Characterization of the World’s Islands. Proc. Natl. Acad. Sci. USA 2013, 110, 15307–15312. [Google Scholar] [CrossRef] [Green Version]
- Zimowski, M.; Leuschner, H.H.; Gärtner, H.; Bergmeier, E. Age and Diversity of Mediterranean Dwarf Shrublands: A Dendrochronological Approach along an Altitudinal Gradient on Crete. J. Veg. Sci. 2014, 25, 122–134. [Google Scholar] [CrossRef]
- Battipaglia, G.; De Micco, V.; Brand, W.A.; Linke, P.; Aronne, G.; Saurer, M.; Cherubini, P. Variations of Vessel Diameter and Δ13C in False Rings of Arbutus unedo L. Reflect Different Environmental Conditions. New Phytol. 2010, 188, 1099–1112. [Google Scholar] [CrossRef] [Green Version]
- Copenheaver, C.A.; Gärtner, H.; Schäfer, I.; Vaccari, F.P.; Cherubini, P. Drought-Triggered False Ring Formation in a Mediterranean Shrub. Botany 2010, 88, 545–555. [Google Scholar] [CrossRef]
- Vieira, J.; Campelo, F.; Nabais, C. Dendrochronology of Maritime Pine in the Middle of the Atlantic Ocean. Dendrochronologia 2017, 45, 73–80. [Google Scholar] [CrossRef]
- Pavão, D.; Jevšenak, J.; Petrillo, M.; Camarinho, R.; Rodrigues, A.; Silva, L.B.; Elias, R.B.; Silva, L. Dendrochronological Potential of the Azorean Endemic Gymnosperm Juniperus brevifolia (Seub.) Antoine. Dendrochronologia 2022, 71, 125901. [Google Scholar] [CrossRef]
- Christopoulou, A.; Gmińska-Nowak, B.; Özarslan, Y.; Ważny, T. Aegean Trees and Timbers: Dendrochronological Survey of the Island of Symi. Forests 2020, 11, 1266. [Google Scholar] [CrossRef]
- Fernández de Castro, A.G.; Rozas, V.; Fuertes-Aguilar, J.; Moreno-Saiz, J.C. Demographic and Dendrochronological Evidence Reveals Highly Endangered Status of a Paleoendemic Woody Mallow from the Canary Islands. Biodivers. Conserv. 2020, 29, 469–485. [Google Scholar] [CrossRef]
- Matos, B.; Silva, L.B.; Camarinho, R.; Rodrigues, A.S.; Rego, R.; Câmara, M.; Silva, L. Linking Dendrometry and Dendrochronology in the Dominant Azorean Tree Laurus azorica (Seub.) Franco. Forests 2019, 10, 538. [Google Scholar] [CrossRef] [Green Version]
- Génova, M.; Ortega, P.; Sadornil, E. The Effects of Fire on Pinus sylvestris L. as Determined by Dendroecological Analysis (Sierra de Gredos, Spain). IForest 2022, 15, 171–178. [Google Scholar] [CrossRef]
- Jonsson, S.; Gunnarson, B.; Criado, C. Drought Is the Major Limiting Factor for Tree-Ring Growth of High-Altitude Canary Island Pines on Tenerife. Geogr. Ann. Ser. A Phys. Geogr. 2002, 84, 51–71. [Google Scholar] [CrossRef]
- Pérez-De-Lis, G.; García-González, I.; Rozas, V.; Arévalo, J.R. Effects of Thinning Intensity on Radial Growth Patterns and Temperature Sensitivity in Pinus canariensis Afforestations on Tenerife Island, Spain. Ann. For. Sci. 2011, 68, 1093–1104. [Google Scholar] [CrossRef] [Green Version]
- Rozas, V.; García-González, I.; Pérez-De-Lis, G.; Arévalo, J.R. Local and Large-Scale Climatic Factors Controlling Tree-Ring Growth of Pinus Canariensis on an Oceanic Island. Clim. Res. 2013, 56, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Rozas, V.; Pérez-de-Lis, G.; García-González, I.; Arévalo, J.R. Contrasting Effects of Wildfire and Climate on Radial Growth of Pinus canariensis on Windward and Leeward Slopes on Tenerife, Canary Islands. Trees–Struct. Funct. 2011, 25, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Palacios, J.M.; Arévalo, J.R.; Balguerías, E.; Barone, R.; de Nascimento, L.; Elias, R.B.; Delgado, J.D.; Fernández-Lugo, S.; Méndez, J.; Naranjo Cigala, A.; et al. La Laurisilva. In Canarias, Madeira y Azores; Macaronesia Editorial: Santa Cruz de Tenerife, Spain, 2017. [Google Scholar]
- Vieira, J.; Carvalho, A.; Campelo, F. Tree Growth Under Climate Change: Evidence From Xylogenesis Timings and Kinetics. Front. Plant Sci. 2020, 11, 90. [Google Scholar] [CrossRef]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Fournier, M. Kinetics of Tracheid Development Explain Conifer Tree-Ring Structure. New Phytol. 2014, 203, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Giguère-Croteau, C.; Boucher, É.; Bergeron, Y.; Girardin, M.P.; Drobyshev, I.; Silva, L.C.R.; Hélie, J.F.; Garneau, M. North America’s Oldest Boreal Trees Are More Efficient Water Users Due to Increased [CO2], but Do Not Grow Faster. Proc. Natl. Acad. Sci. USA 2019, 116, 2749–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchand, W.; Girardin, M.P.; Hartmann, H.; Depardieu, C.; Isabel, N.; Gauthier, S.; Boucher, É.; Bergeron, Y. Strong Overestimation of Water-Use Efficiency Responses to Rising CO2 in Tree-Ring Studies. Glob. Chang. Biol. 2020, 26, 4538–4558. [Google Scholar] [CrossRef] [PubMed]
- Baldocchi, D.; Penuelas, J. The Physics and Ecology of Mining Carbon Dioxide from the Atmosphere by Ecosystems. Glob. Chang. Biol. 2019, 25, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Wang, X.; Liu, T.; Guo, L.; Wang, B.; Khan, A. Impact of Competition on the Growth of Pinus tabulaeformis in Response to Climate on the Loess Plateau of China. Plant Ecol. 2022, 223, 353–368. [Google Scholar] [CrossRef]
- Luo, Y.; McIntire, E.J.B.; Boisvenue, C.; Nikiema, P.P.; Chen, H.Y.H. Climatic Change Only Stimulated Growth for Trees under Weak Competition in Central Boreal Forests. J. Ecol. 2020, 108, 36–46. [Google Scholar] [CrossRef]
- Azevedo, E.M.V.B. Condicionantes Dinâmicas Do Clima Do Arquipélago Dos Açores. Elementos Para o Seu Estudo. Açoreana 2001, 9, 309–317. [Google Scholar]
- Couto, M.A.G. Atlas Climático de Los Archipiélagos de Canarias, Madeira y Azores Atlas Climático Dos Arquipélagos Das Canárias, Da Madeira e Dos Açores; Aemet: Madrid, Spain, 2012; p. 79. [Google Scholar]
- Borges Silva, L.C.; Pavão, D.C.; Elias, R.B.; Moura, M.; Ventura, M.A.; Silva, L. Taxonomic, Structural Diversity and Carbon Stocks in a Gradient of Island Forests. Sci. Rep. 2022, 12, 1–16. [Google Scholar] [CrossRef]
- Reis-Avila, G.; Oliveira, J.M. Lauraceae: A Promising Family for the Advance of Neotropical Dendrochronology. Dendrochronologia 2017, 44, 103–116. [Google Scholar] [CrossRef]
- Schweingruber, F.H.; Börner, A.; Schulze, E.-D. Atlas of Stem Anatomy in Herbs, Shrubs and Trees—Volume 2; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642320434-0. [Google Scholar]
- Balabasquer, L.D.V. Dendroecología de Laurus novocanariensis En Cinco Localidades de Tenerife, Islas Canarias; Universidad de La Laguna: La Laguna, Spain, 2021. [Google Scholar]
- García-López, M.A.; Rozas, V.; Olano, J.M.; Sangüesa-Barreda, G.; García-Hidalgo, M.; Gómez-González, S.; López-Rubio, R.; Fernández-Palacios, J.M.; García-González, I.; García-Cervigón, A.I. Tree-Ring Distinctness, Dating Potential and Climatic Sensitivity of Laurel Forest Tree Species in Tenerife Island. Dendrochronologia 2022, 76, 126011. [Google Scholar] [CrossRef]
- Granato-Souza, D.; Adenesky-Filho, E.; Esemann-Quadros, K. Dendrochronology and Climatic Signals in the Wood of Nectandra oppositifolia from a Dense Rain Forest in Southern Brazil. J. For. Res. 2019, 30, 545–553. [Google Scholar] [CrossRef]
- Malheiro, A. Geological Hazards in the Azores Archipelago: Volcanic Terrain Instability and Human Vulnerability. J. Volcanol. Geotherm. Res. 2006, 156, 158–171. [Google Scholar] [CrossRef]
- Louvat, P.; Allègre, C.J. Riverine Erosion Rates on Sao Miguel Volcanic Island, Azores Archipelago. USDA For. Serv.-Gen. Tech. Rep. RMRS-GTR 1998, 148, 177–200. [Google Scholar] [CrossRef]
- Fontes, J.C.; Pereira, L.S.; Smith, R.E. Runoff and Erosion in Volcanic Soils of Azores: Simulation with OPUS. Catena 2004, 56, 199–212. [Google Scholar] [CrossRef]
- Wunder, J. Technical Report: Age Estimation of Pittosporum undulatum from São Miguel, Azores, Portugal; Wunder Consulting: Pfäffikon, Switzerland, 2010. [Google Scholar]
- Teixeira, A.; Mir, C.; Borges Silva, L.; Hahndorf, I.; Silva, L. Invasive Woodland Resources in the Azores: Biomass Availability for 100% Renewable Energy Supply in Graciosa Island. In Proceedings of the 23rd European Biomass Conference and Exhibition, Vienna, Austria, 1–4 June 2015. [Google Scholar]
- Borges Silva, L.; Teixeira, A.; Alves, M.; Elias, R.B.; Silva, L. Tree Age Determination in the Widespread Woody Plant Invader Pittosporum undulatum. For. Ecol. Manag. 2017, 400, 457–467. [Google Scholar] [CrossRef]
- Borges Silva, L.; Lourenço, P.; Teixeira, A.; Azevedo, E.B.; Alves, M.; Elias, R.B.; Silva, L. Biomass Valorization in the Management of Woody Plant Invaders: The Case of Pittosporum undulatum in the Azores. Biomass Bioenergy 2018, 109, 155–165. [Google Scholar] [CrossRef]
- Pavão, D.; Jevšenak, J.; Engblom, J.; Silva, L.B.; Elias, R.B.; Silva, L. Tree Growth-Climate Relationship in the Azorean Holly in a Temperate Humid Forest with Low Thermal Amplitude. Dendrochronologia 2023, 77, 126050. [Google Scholar] [CrossRef]
- Câmara, M.I.M. Tree Age Estimation in an Invaded Natural Forest in São Miguel Island, Azores; Universidade dos Açores: Ponta Delgada, Portugal, 2016. [Google Scholar]
- Rego, R.; Borges Silva, L.; Medeiros, F.; Porteiro, J.; Silva, L. Ecological Characterization as the First Step Towards the Conservation of Natural Unprotected Areas: A Case Study in the Azores. In Proceedings of the European Meeting of Phytosociology, Biogeography and Syntaxonomy of the Atlantic Regions, Praia, Cape Verde, 5–7 November 2017. [Google Scholar]
- Schweingruber, F.H.; Börner, A.; Schulze, E.-D. Atlas of Stem Anatomy in Herbs, Shrubs and Trees—Volume 1; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 9783642116377. [Google Scholar]
- Buttó, V.; Millan, M.; Rossi, S.; Delagrange, S. Contrasting Carbon Allocation Strategies of Ring-Porous and Diffuse-Porous Species Converge Toward Similar Growth Responses to Drought. Front. Plant Sci. 2021, 12, 2990. [Google Scholar] [CrossRef]
- García González, I.; Eckstein, D. Climatic Signal of Earlywood Vessels of Oak on a Maritime Site. Tree Physiol. 2003, 23, 497–504. [Google Scholar] [CrossRef]
- Miranda, J.M.; Luis, J.F.; Lourenço, N.; Goslin, J. Distributed Deformation Close to the Azores Triple “Point”. Mar. Geol. 2014, 355, 27–35. [Google Scholar] [CrossRef]
- Forjaz, V.H.; Tavares, J.M.; Azevedo, E.M.V.B.; Nunes, J.C. Atlas Básico Dos Açores; Observatório Vulcanológico e Geotérmico dos Açores: Lagoa, Portugal, 2004. [Google Scholar]
- Costa, A.C.G.; Hildenbrand, A.; Marques, F.O.; Sibrant, A.L.R.; Santos de Campos, A. Catastrophic Flank Collapses and Slumping in Pico Island during the Last 130 Kyr (Pico-Faial Ridge, Azores Triple Junction). J. Volcanol. Geotherm. Res. 2015, 302, 33–46. [Google Scholar] [CrossRef]
- Ramalho, R.S.; Helffrich, G.; Madeira, J.; Cosca, M.; Thomas, C.; Quartau, R.; Hipólito, A.; Rovere, A.; Hearty, P.J.; Ávila, S.P. Emergence and Evolution of Santa Maria Island (Azores)—The Conundrum of Uplifted Islands Revisited. Bull. Geol. Soc. Am. 2017, 129, 372–391. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.E. Erosão e Degradação Do Solo Agrícola. Bol. Da Com. Regul. Dos Cereais 1952, 15, 25–44. [Google Scholar]
- Pinheiro, J. Caracterização Geral Dos Solos Da Ilha Terceira (Açores) Que Se Enquadram Na Ordem Andisol. An. Do Inst. Super. Agron. 1999, 47, 99–117. [Google Scholar]
- Ricardo, R.P.; Madeira, M.A.V.; Medina, J.M.B.; Marques, M.M.; Furtado, F.A.S. Esboço Pedológico Da Ilha de S. Miguel (Açores). An. do Inst. Super. Agron. 1977, 275–385. [Google Scholar]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Quinet, A.; Andreata, R.H.P. Lauraceae Jussieu Na Reserva Ecológica de Macaé de Cima, Município de Nova Friburgo, Rio de Janeiro, Brasil. Rodriguesia 2002, 53, 59–121. [Google Scholar] [CrossRef]
- Marques, C.A. Economic Importance of Family Lauraceae Linal. Floresta E Ambient. 2001, 8, 195–206. [Google Scholar]
- Silva, L.; Beech, E. Laurus azorica. The IUCN Red List of Threatened Species 2017: E.T38397A81868030; IUCN Global Species Programme Red List Unit: Cambridge, UK, 2017. [Google Scholar]
- DRRF. Plano de Gestão Florestal Do Perímetro Florestal e Matas Regionais Da Ilha de São Miguel; Ponta Delgada: Região Autónoma dos Açores, Portugal, 2017. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; James, H.S., Ed.; University of Arizona Press: Tucson, AZ, USA, 2010; Volume 26. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Schweingruber, F.H. Tree Rings: Basics and Applications of Dendrochronology; Kluwer: Dordrecht, The Netherlands, 1988. [Google Scholar]
- Buras, A.; Wilmking, M. Correcting the Calculation of Gleichläufigkeit. Dendrochronologia 2015, 34, 29–30. [Google Scholar] [CrossRef]
- Bunn, A.G. A Dendrochronology Program Library in R (DplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Bontemps, J.D.; Esper, J. Statistical Modelling and RCS Detrending Methods Provide Similar Estimates of Long-Term Trend in Radial Growth of Common Beech in North-Eastern France. Dendrochronologia 2011, 29, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Kairiukstis, L. Methods of Dendrochronology: Applications in the Environmental Science; Cook, E.R., Kairiukstis, L.A., Eds.; Springer-Science+Business Media B.V.: Berlin/Heidelberg, Germany, 1990; ISBN 978-90-481-4060-2. [Google Scholar]
- Esper, J.; Schneider, L.; Smerdon, J.E.; Schöne, B.R.; Büntgen, U. Signals and Memory in Tree-Ring Width and Density Data. Dendrochronologia 2015, 35, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Brienen, R.J.W.; Schöngart, J.; Zuidema, P.A. Tree Rings in the Tropics: Insights into the Ecology and Climate Sensitivity of Tropical Trees. In Tropical Tree Physiology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 439–461. ISBN 9783319274225. [Google Scholar]
- Cook, E.R. A Time Sires Analysis Approach to Tree Ring Standardization; Univeristy of Arizona: Tucson, Arizona, 1985. [Google Scholar]
- Cook, E.R.; Pederson, N. Uncertainty, Emergence, and Statistics in Dendrochronology; Springer: Dordrecht, The Netherlands, 2011; ISBN 9781402057250. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series with Applications in Dendroclimatology and Hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Buras, A. A Comment on the Expressed Population Signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, P.; Kessler, M. Climatologies at High Resolution for the Earth Land Surface Areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [Green Version]
- Karger, D.N.; Zimmermann, N.E. CHELSAcruts–High Resolution Temperature and Precipitation Timeseries for the 20th Century and Beyond. EnviDat 2018. [Google Scholar] [CrossRef]
- Jevšenak, J.; Džeroski, S.; Zavadlav, S.; Levanič, T. A Machine Learning Approach to Analyzing the Relationship between Temperatures and Multi-Proxy Tree-Ring Records. Tree-Ring Res. 2018, 74, 210–224. [Google Scholar] [CrossRef]
- Sahour, H.; Gholami, V.; Torkaman, J.; Vazifedan, M.; Saeedi, S. Random Forest and Extreme Gradient Boosting Algorithms for Streamflow Modeling Using Vessel Features and Tree-Rings. Environ. Earth Sci. 2021, 80, 1–14. [Google Scholar] [CrossRef]
- Cui, J.P.; Qureshi, S.; Harris, A.; Jim, C.Y.; Wang, H.F. Venerable Trees of Tropical Chinese Wuzhishan City: Distribution Patterns and Drivers. Urban Ecosyst. 2022, 25, 1765–1776. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical Temperatures for Xylogenesis in Conifers of Cold Climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Archer, K.J.; Kimes, R.V. Empirical Characterization of Random Forest Variable Importance Measures. Comput. Stat. Data Anal. 2008, 52, 2249–2260. [Google Scholar] [CrossRef]
- Li, X.; Chen, G.; Qin, W.; Wang, X.; Liu, H.; Wang, P. Differences in Responses of Tree-Ring Δ13C in Angiosperms and Gymnosperms to Climate Change on a Global Scale. For. Ecol. Manag. 2021, 492, 119247. [Google Scholar] [CrossRef]
- Jevšenak, J.; Skudnik, M. A Random Forest Model for Basal Area Increment Predictions from National Forest Inventory Data. For. Ecol. Manag. 2021, 479, 118601. [Google Scholar] [CrossRef]
- Lucas, T.C.D. A Translucent Box: Interpretable Machine Learning in Ecology. Ecol. Monogr. 2020, 90, e01422. [Google Scholar] [CrossRef]
- Ávila, S.P.; Cordeiro, R.; Madeira, P.; Silva, L.; Medeiros, A.; Rebelo, A.C.; Melo, C.; Neto, A.I.; Haroun, R.; Monteiro, A.; et al. Global Change Impacts on Large-Scale Biogeographic Patterns of Marine Organisms on Atlantic Oceanic Islands. Mar. Pollut. Bull. 2018, 126, 101–112. [Google Scholar] [CrossRef]
- Parelho, C.; Rodrigues, A.; Barreto, M. do C.; Cruz, J.V.; Rasche, F.; Silva, L.; Garcia, P. Bioaccumulation and Potential Ecotoxicological Effects of Trace Metals along a Management Intensity Gradient in Volcanic Pasturelands. Chemosphere 2021, 273, 128601. [Google Scholar] [CrossRef]
- Barreiro, A.; Fox, A.; Jongen, M.; Melo, J.; Musyoki, M.; Vieira, A.; Zimmermann, J.; Carlsson, G.; Cruz, C.; Lüscher, A.; et al. Soil Bacteria Respond to Regional Edapho-Climatic Conditions While Soil Fungi Respond to Management Intensity in Grasslands along a European Transect. Appl. Soil Ecol. 2022, 170, 104264. [Google Scholar] [CrossRef]
- Sheppard, P.R. Dendroclimatology: Extracting Climate from Trees. Wiley Interdiscip. Rev. Clim. Chang. 2010, 1, 343–352. [Google Scholar] [CrossRef]
- Johnson, J.B.; Omland, K.S. Model Selection in Ecology and Evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. Maximum Likelihood Identification of Gaussian Autoregressive Moving Average Models. Biometrika 1973, 60, 255–265. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Springer Science + Businees Media, LLC: New York, NY, USA, 2007. [Google Scholar]
- Brienen, R.J.W.; Zuidema, P.A. Relating Tree Growth to Rainfall in Bolivian Rain Forests: A Test for Six Species Using Tree Ring Analysis. Oecologia 2005, 146, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, C.I.; Camarero, J.J.; Gusmán, A.A. Site-Dependent Growth Responses to Climate in Two Major Tree Species from Tropical Dry Forests of Southwest Ecuador. Dendrochronologia 2018, 52, 11–19. [Google Scholar] [CrossRef]
- Granato-Souza, D.; Adenesky-Filho, E.; Barbosa, A.C.M.C.; Esemann-Quadros, K. Dendrochronological Analyses and Climatic Signals of Alchornea triplinervia in Subtropical Forest of Southern Brazil. Austral Ecol. 2018, 43, 385–396. [Google Scholar] [CrossRef]
- Vasconcellos, T.J.D.; Tomazello-Filho, M.; Callado, C.H. Dendrochronology and Dendroclimatology of Ceiba speciosa (A. St.-Hil.) Ravenna (Malvaceae) Exposed to Urban Pollution in Rio de Janeiro City, Brazil. Dendrochronologia 2019, 53, 104–113. [Google Scholar] [CrossRef]
- Ford, K.R.; Breckheimer, I.K.; Franklin, J.F.; Freund, J.A.; Kroiss, S.J.; Larson, A.J.; Theobald, E.J.; HilleRisLambers, J. Competition Alters Tree Growth Responses to Climate at Individual and Stand Scales. Can. J. For. Res. 2017, 47, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Wood Growth in Pure and Mixed Quercus ilex L. Forests: Drought Influence Depends on Site Conditions. Front. Plant Sci. 2019, 10, 397. [Google Scholar] [CrossRef]
- Bhuyan, U.; Zang, C.; Vicente-Serrano, S.M.; Menzel, A. Exploring Relationships among Tree-Ring Growth, Climate Variability, and Seasonal Leaf Activity on Varying Timescales and Spatial Resolutions. Remote Sens. 2017, 9, 526. [Google Scholar] [CrossRef] [Green Version]
- Lopatin, E.; Kolström, T.; Spiecker, H. Determination of Forest Growth Trends in Komi Republic (Northwestern Russia): Combination of Tree-Ring Analysis and Remote Sensing Data. Boreal Environ. Res. 2006, 11, 341–353. [Google Scholar]
- Babst, F.; Bodesheim, P.; Charney, N.; Friend, A.D.; Girardin, M.P.; Klesse, S.; Moore, D.J.P.; Seftigen, K.; Björklund, J.; Bouriaud, O.; et al. When Tree Rings Go Global: Challenges and Opportunities for Retro- and Prospective Insight. Quat. Sci. Rev. 2018, 197, 1–20. [Google Scholar] [CrossRef]
- Arlot, S.; Genuer, R. Analysis of Purely Random Forests Bias. arXiv 2014, arXiv:1407.3939. [Google Scholar]
- Painsky, A.; Rosset, S. Cross-Validated Variable Selection in Tree-Based Methods Improves Predictive Performance. IEEE Trans. Pattern Anal. Mach. Intell. 2017, 39, 2142–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piryonesi, S.M.; El-Diraby, T.E. Role of Data Analytics in Infrastructure Asset Management: Overcoming Data Size and Quality Problems. J. Transp. Eng. Part B Pavements 2020, 146, 04020022. [Google Scholar] [CrossRef]
- Fox, E.W.; Hill, R.A.; Leibowitz, S.G.; Olsen, A.R.; Thornbrugh, D.J.; Weber, M.H. Assessing the Accuracy and Stability of Variable Selection Methods for Random Forest Modeling in Ecology. Environ. Monit. Assess. 2017, 189, 316. [Google Scholar] [CrossRef]
- Zheng, B.; Agresti, A. Summarizing the Predictive Power of a Generalized Linear Model. Stat. Med. 2000, 19, 1771–1781. [Google Scholar] [CrossRef]
- Dobson, A.J.; Barnett, A.G. An Introduction to Generalized Linear Models, 4th ed.; Dobson, A.J., Barnett, A.G., Eds.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 978-1-138-74168-3. [Google Scholar]
- Cermák, J.; Jiménez, M.S.; González-Rodríguez, A.M.; Morales, D. Laurel Forests in Tenerife, Canary Islands: II. Efficiency of the Water Conducting System in Laurus azorica Trees. Trees–Struct. Funct. 2002, 16, 538–546. [Google Scholar] [CrossRef]
- Tixier, A.; Gambetta, G.A.; Godfrey, J.; Orozco, J.; Zwieniecki, M.A. Non-Structural Carbohydrates in Dormant Woody Perennials; The Tale of Winter Survival and Spring Arrival. Front. For. Glob. Chang. 2019, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.Y.S.; Young, D.J.N.; Latimer, A.M.; Buckley, T.N.; Magney, T.S. Importance of the Legacy Effect for Assessing Spatiotemporal Correspondence between Interannual Tree-Ring Width and Remote Sensing Products in the Sierra Nevada. Remote Sens. Environ. 2021, 265, 112635. [Google Scholar] [CrossRef]
- Körner, C. Carbon Limitation in Trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Paradigm Shift in Plant Growth Control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Prada, S.; Menezes de Sequeira, M.; Figueira, C.; da Silva, M.O. Fog Precipitation and Rainfall Interception in the Natural Forests of Madeira Island (Portugal). Agric. For. Meteorol. 2009, 149, 1179–1187. [Google Scholar] [CrossRef]
- Dias, E.; Melo, C. Factors Influencing the Distribution of Azorean Mountain Vegetation: Implications for Nature Conservation. Biodivers. Conserv. 2010, 19, 3311–3326. [Google Scholar] [CrossRef]
- Dawadi, B.; Liang, E.; Tian, L.; Devkota, L.P.; Yao, T. Pre-Monsoon Precipitation Signal in Tree Rings of Timberline Betula utilis in the Central Himalayas. Quat. Int. 2013, 283, 72–77. [Google Scholar] [CrossRef]
- Thapa, U.K.; Shah, S.K.; Gaire, N.P.; Bhuju, D.R. Spring Temperatures in the Far-Western Nepal Himalaya since AD 1640 Reconstructed from Picea smithiana Tree-Ring Widths. Clim. Dyn. 2015, 45, 2069–2081. [Google Scholar] [CrossRef]
- Schäfer, H. Flora of the Azores–A Field Guide; 2nd ed.; Margraf Publishers: Weikersheim, Duitsland, 2005. [Google Scholar]
- Zani, D.; Crowther, T.W.; Mo, L.; Renner, S.S.; Zohner, C.M. Increased Growing-Season Productivity Drives Earlier Autumn Leaf Senescence in Temperate Trees. Science 2020, 37, 1066–1071. [Google Scholar] [CrossRef]
- Babst, F.; Bouriaud, O.; Poulter, B.; Trouet, V.; Girardin, M.P.; Frank, D.C. Twentieth Century Redistribution in Climatic Drivers of Global Tree Growth. Sci. Adv. 2019, 5, eaat4313. [Google Scholar] [CrossRef]
- Harvey, J.E.; Smiljanić, M.; Scharnweber, T.; Buras, A.; Cedro, A.; Cruz-García, R.; Drobyshev, I.; Janecka, K.; Jansons, Ā.; Kaczka, R.; et al. Tree Growth Influenced by Warming Winter Climate and Summer Moisture Availability in Northern Temperate Forests. Glob. Chang. Biol. 2020, 26, 2505–2518. [Google Scholar] [CrossRef]
- Weijers, S.; Pape, R.; Löffler, J.; Myers-Smith, I.H. Contrasting Shrub Species Respond to Early Summer Temperatures Leading to Correspondence of Shrub Growth Patterns. Environ. Res. Lett. 2018, 13, 034005. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Y.; Zhao, S.; Jiao, L.; Wen, Y. Relationships between Tree Age and Climate Sensitivity of Radial Growth in Different Drought Conditions of Qilian Mountains, Northwestern China. Forests 2018, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Biondi, F. Are Climate-Tree Growth Relationships Changing in North-Central Idaho, U.S.A.? Arct. Antarct. Alp. Res. 2000, 32, 111–116. [Google Scholar] [CrossRef]
- Fritts, H.C. Growth-Rings of Trees: Their Correlation with Climate. Science 1966, 154, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Qaderi, M.M.; Martel, A.B.; Dixon, S.L. Environmental Factors Influence Plant Vascular System and Water Regulation. Plants 2019, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Oliver, M.; Fullana-Pericàs, M.; Bota, J.; Flexas, J. Adjustments in Photosynthesis and Leaf Water Relations Are Related to Changes in Cell Wall Composition in Hordeum vulgare and Triticum aestivum Subjected to Water Deficit Stress. Plant Sci. 2021, 311, 111015. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.Q.; Durgante, F.M.; Wittmann, F.; Teresa, M.; Piedade, F.; Ricardo, D.; Rodriguez, O.; Tomazello, M.; Pia, F.; Schöngart, J. Minimum Temperature and Evapotranspiration in Central Amazonian Floodplains Limit Tree Growth of Nectandra amazonum (Lauraceae). Trees 2021, 35, 1367–1384. [Google Scholar] [CrossRef]
- Sun, M.; Li, J.; Cao, R.; Tian, K.; Zhang, W.; Yin, D.; Zhang, Y. Climate-Growth Relations of Abies Georgei along an Altitudinal Gradient in Haba Snow Mountain, Southwestern China. Forests 2021, 12, 1569. [Google Scholar] [CrossRef]
- Camarero, J.J.; Rubio-Cuadrado, A. Relating Climate, Drought and Radial Growth in Broadleaf Mediterranean Tree and Shrub Species: A New Approach to Quantify Climate-Growth Relationships. Forests 2020, 11, 1250. [Google Scholar] [CrossRef]
- Roibu, C.C.; Sfecla, V.; Mursa, A.; Ionita, M.; Nagavciuc, V.; Chiriloaei, F.; Lesan, I.; Popa, I. The Climatic Response of Tree Ring Width Components of Ash (Fraxinus excelsior L.) and Common Oak (Quercus robur L.) from Eastern Europe. Forests 2020, 11, 600. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Herrmann, V.; Rollinson, C.R.; Gonzalez, B.; Gonzalez-Akre, E.B.; Pederson, N.; Alexander, M.R.; Allen, C.D.; Alfaro-Sánchez, R.; Awada, T.; et al. Joint Effects of Climate, Tree Size, and Year on Annual Tree Growth Derived from Tree-Ring Records of Ten Globally Distributed Forests. Glob. Chang. Biol. 2022, 28, 245–266. [Google Scholar] [CrossRef]
- Boninsegna, J.A.; Argollo, J.; Aravena, J.C.; Barichivich, J.; Christie, D.; Ferrero, M.E.; Lara, A.; Le Quesne, C.; Luckman, B.H.; Masiokas, M.; et al. Dendroclimatological Reconstructions in South America: A Review. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 281, 210–228. [Google Scholar] [CrossRef]
- Babst, F.; Poulter, B.; Trouet, V.; Tan, K.; Neuwirth, B.; Wilson, R.; Carrer, M.; Grabner, M.; Tegel, W.; Levanic, T.; et al. Site- and Species-Specific Responses of Forest Growth to Climate across the European Continent. Glob. Ecol. Biogeogr. 2013, 22, 706–717. [Google Scholar] [CrossRef]
- Hacket-Pain, A.J.; Friend, A.D.; Lageard, J.G.A.; Thomas, P.A. The Influence of Masting Phenomenon on Growth-Climate Relationships in Trees: Explaining the Influence of Previous Summers’ Climate on Ring Width. Tree Physiol. 2015, 35, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Wu, G.; Liu, X.; Chen, T.; Wang, B.; Hudson, A.; Trouet, V. Age-Related Climate Response of Tree-Ring Δ13C and Δ18O From Spruce in Northwestern China, With Implications for Relative Humidity Reconstructions. J. Geophys. Res. Biogeosci. 2020, 125, 1–18. [Google Scholar] [CrossRef]
- Timofeeva, G.; Treydte, K.; Bugmann, H.; Rigling, A.; Schaub, M.; Siegwolf, R.; Saurer, M. Long-Term Effects of Drought on Tree-Ring Growth and Carbon Isotope Variability in Scots Pine in a Dry Environment. Tree Physiol. 2017, 37, 1028–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaMarche, V.C.; Stockton, C.W. Chronologies from Temperature-Sensitive Bristlecone Pines at Upper Treeline in Western United States. Tree-Ring Bull. 1974, 34, 21–45. [Google Scholar]
- Szejner, P.; Wright, W.E.; Belmecheri, S.; Meko, D.; Leavitt, S.W.; Ehleringer, J.R.; Monson, R.K. Disentangling Seasonal and Interannual Legacies from Inferred Patterns of Forest Water and Carbon Cycling Using Tree-Ring Stable Isotopes. Glob. Chang. Biol. 2018, 24, 5332–5347. [Google Scholar] [CrossRef]
- Smith, M.G.; Miller, R.E.; Arndt, S.K.; Kasel, S.; Bennett, L.T. Whole-Tree Distribution and Temporal Variation of Non-Structural Carbohydrates in Broadleaf Evergreen Trees. Tree Physiol. 2018, 38, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Kannenberg, S.A.; Maxwell, J.T.; Pederson, N.; D’Orangeville, L.; Ficklin, D.L.; Phillips, R.P. Drought Legacies Are Dependent on Water Table Depth, Wood Anatomy and Drought Timing across the Eastern US. Ecol. Lett. 2019, 22, 119–127. [Google Scholar] [CrossRef]
- Liu, W.; Su, J.; Li, S.; Lang, X.; Huang, X. Non-Structural Carbohydrates Regulated by Season and Species in the Subtropical Monsoon Broad-Leaved Evergreen Forest of Yunnan Province, China. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Monserud, R.A.; Marshall, J.D. Time-Series Analysis of Δ13C from Tree Rings. I. Time Trends and Autocorrelation. Tree Physiol. 2001, 21, 1087–1102. [Google Scholar] [CrossRef] [Green Version]
- McCarroll, D.; Whitney, M.; Young, G.H.F.; Loader, N.J.; Gagen, M.H. A Simple Stable Carbon Isotope Method for Investigating Changes in the Use of Recent versus Old Carbon in Oak. Tree Physiol. 2017, 37, 1021–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primicia, I.; Camarero, J.J.; Janda, P.; Čada, V.; Morrissey, R.C.; Trotsiuk, V.; Bače, R.; Teodosiu, M.; Svoboda, M. Age, Competition, Disturbance and Elevation Effects on Tree and Stand Growth Response of Primary Picea abies Forest to Climate. For. Ecol. Manag. 2015, 354, 77–86. [Google Scholar] [CrossRef]
- Lu, K.; Chen, N.; Zhang, C.; Dong, X.; Zhao, C. Drought Enhances the Role of Competition in Mediating the Relationship between Tree Growth and Climate in Semi-Arid Areas of Northwest China. Forests 2019, 10, 804. [Google Scholar] [CrossRef] [Green Version]
- Moser-Reischl, A.; Rahman, M.A.; Pauleit, S.; Pretzsch, H.; Rötzer, T. Growth Patterns and Effects of Urban Micro-Climate on Two Physiologically Contrasting Urban Tree Species. Landsc. Urban Plan. 2019, 183, 88–99. [Google Scholar] [CrossRef]
- Wang, B.; Yu, P.; Yu, Y.; Wan, Y.; Wang, Y.; Zhang, L.; Wang, S.; Wang, X.; Liu, Z.; Xu, L. Effects of Canopy Position on Climate-Growth Relationships of Qinghai Spruce in the Central Qilian Mountains, Northwestern China. Dendrochronologia 2020, 64, 125756. [Google Scholar] [CrossRef]
- Yang, J.; Cooper, D.J.; Li, Z.; Song, W.; Zhang, Y.; Zhao, B.; Han, S.; Wang, X. Differences in Tree and Shrub Growth Responses to Climate Change in a Boreal Forest in China. Dendrochronologia 2020, 63, 125744. [Google Scholar] [CrossRef]
- Foster, J.R.; Finley, A.O.; D’Amato, A.W.; Bradford, J.B.; Banerjee, S. Predicting Tree Biomass Growth in the Temperate-Boreal Ecotone: Is Tree Size, Age, Competition, or Climate Response Most Important? Glob. Chang. Biol. 2016, 22, 2138–2151. [Google Scholar] [CrossRef]
- Rollinson, C.R.; Kaye, M.W.; Canham, C.D. Interspecific Variation in Growth Responses to Climate and Competition of Five Eastern Tree Species. Ecology 2016, 97, 1003–1011. [Google Scholar] [CrossRef]
- van der Maaten-Theunissen, M.; Bouriaud, O. Climate-Growth Relationships at Different Stem Heights in Silver Fir and Norway Spruce. Can. J. For. Res. 2012, 42, 958–969. [Google Scholar] [CrossRef]
- Mazza, G.; Gallucci, V.; Manetti, M.C.; Urbinati, C. Climate-Growth Relationships of Silver Fir (Abies alba Mill.) in Marginal Populations of Central Italy. Dendrochronologia 2014, 32, 181–190. [Google Scholar] [CrossRef]
- Misi, D.; Puchałka, R.; Pearson, C.; Robertson, I.; Koprowski, M. Differences in the Climate-Growth Relationship of Scots Pine: A Case Study from Poland and Hungary. Forests 2019, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Henderson, M.; Liu, B.; Yan, H. Directional Variability in Response of Pinus koraiensis Radial Growth to Climate Change. Forests 2021, 12, 1684. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDMs: Species Distribution Models That Account for Local Adaptation and Phenotypic Plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef] [PubMed]
- Kumarathunge, D.P.; Medlyn, B.E.; Drake, J.E.; Tjoelker, M.G.; Aspinwall, M.J.; Battaglia, M.; Cano, F.J.; Carter, K.R.; Cavaleri, M.A.; Cernusak, L.A.; et al. Acclimation and Adaptation Components of the Temperature Dependence of Plant Photosynthesis at the Global Scale. New Phytol. 2019, 222, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J.; Goad, D.M.; Gross, B.L.; Muñoz, P.R.; Olsen, K.M. Genetic Trade-Offs Underlie Divergent Life History Strategies for Local Adaptation in White Clover. Mol. Ecol. 2022, 31, 3742–3760. [Google Scholar] [CrossRef]
- Ren, L.; Guo, X.; Liu, S.; Yu, T.; Guo, W.; Wang, R.; Ye, S.; Lambertini, C.; Brix, H.; Eller, F. Intraspecific Variation in Phragmites australis: Clonal Adaption of Functional Traits and Phenotypic Plasticity Vary with Latitude of Origin. J. Ecol. 2020, 108, 2531–2543. [Google Scholar] [CrossRef]
- Souto-Herrero, M.; Rozas, V.; García-González, I. Earlywood Vessels and Latewood Width Explain the Role of Climate on Wood Formation of Quercus pyrenaica Willd. across the Atlantic-Mediterranean Boundary in NW Iberia. For. Ecol. Manag. 2018, 425, 126–137. [Google Scholar] [CrossRef]
- García-González, I.; Fonti, P. Selecting Earlywood Vessels to Maximize Their Environmental Signal. Tree Physiol. 2006, 26, 1289–1296. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate Variable | Year | Variable |
|---|---|---|
| Temperature | Current | Mean annual temperature |
| Mean annual minimum temperature | ||
| Mean annual maximum temperature | ||
| Mean winter temperature | ||
| Mean spring temperature | ||
| Mean summer temperature | ||
| Mean autumn temperature | ||
| Mean winter minimum temperature | ||
| Mean spring minimum temperature | ||
| Mean summer minimum temperature | ||
| Mean autumn minimum temperature | ||
| Mean winter maximum temperature | ||
| Mean spring maximum temperature | ||
| Mean summer maximum temperature | ||
| Mean autumn maximum temperature | ||
| Monthly mean temperature | ||
| Monthly minimum temperature | ||
| Monthly maximum temperature | ||
| Previous | Mean annual temperature | |
| Mean annual minimum temperature | ||
| Mean annual maximum temperature | ||
| Monthly mean temperature | ||
| Monthly minimum temperature | ||
| Monthly maximum temperature | ||
| Precipitation | Current | Annual precipitation |
| Winter precipitation | ||
| Spring precipitation | ||
| Summer precipitation | ||
| Autumn precipitation | ||
| Monthly precipitation | ||
| Previous | Annual precipitation | |
| Monthly precipitation |
| Number | Model | Current | Previous | Both |
|---|---|---|---|---|
| Random Forest Step 1 | ||||
| 1 | Saturated model (All variables) | x | ||
| 2 | Partially saturated model (All variables) | x | ||
| 3 | Partially saturated model (All variables) | x | ||
| 4 | Temperature model | x | ||
| 5 | Temperature model | x | ||
| 6 | Temperature model | x | ||
| 7 | Precipitation model | x | ||
| 8 | Precipitation model | x | ||
| 9 | Precipitation model | x | ||
| Random Forest Step 2 | ||||
| 10 | Optimal model (Selected variables) | x | x | x |
| GLMs | ||||
| 11 | Reduced model (Selected variables) | x | x | x |
| ClimVar | CC | GM | MT | PP | LO | PO | NO | ClimVar | CC | GM | MT | PP | LO | PO | NO |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T4 | + | TminPre6 | + | ||||||||||||
| T6 | - | - | TminPre7 | - | |||||||||||
| T10 | - | TminPre10 | + | ||||||||||||
| Tmin3 | + | TminPre12 | + | ||||||||||||
| Tmin5 | + | + | TmaxPre6 | + | - | + | |||||||||
| Tmin10 | + | - | TmaxPre10 | + | |||||||||||
| Tmax3 | - | P1 | + | ||||||||||||
| Tmax5 | - | - | P9 | + | |||||||||||
| Tmax6 | Ppre4 | + | |||||||||||||
| Tpre2 | - | Ppre5 | - | ||||||||||||
| Tpre6 | + | Ppre8 | + | ||||||||||||
| Tpre10 | + | Ppre10 | + | ||||||||||||
| TminPre2 | - |
| Chronology | Model | Coefficients | AIC | AICc | Null Model AIC | R2 |
|---|---|---|---|---|---|---|
| CC | Tmin5 *** | +0.23618 | 317.02 | 317.234 | 349.084 | 0.171 |
| TmaxPre6 ** | +0.11627 | |||||
| GM | Tmax6 *** | −0.29387 | 15.592 | 17.074 | 27.623 | 0.386 |
| TmaxPre6 ** | −0.22347 | |||||
| MT | Ppre8 * | +0.002231 | 11.588 | 12.445 | 14.284 | 0.125 |
| PP | Tmin5 *** | +0.6203 | 70.255 | 71.736 | 81.250 | 0.366 |
| TminPre6 * | +0.4827 | |||||
| LO | T6 * | −0.0882183 | −13.663 | −12.182 | −8.901 | 0.230 |
| Ppre5 * | −0.0008316 | |||||
| PO | TminPre7 * | −0.14009 | −7.3785 | −5.897 | 7.716 | 0.442 |
| TminPre10 *** | +0.12668 | |||||
| NO | Tpre6 * | +0.05826 | −45.015 | −43.534 | −37.001 | 0.304 |
| TminPre2 ** | −0.06556 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavão, D.C.; Jevšenak, J.; Silva, L.B.; Elias, R.B.; Silva, L. Climate–Growth Relationships in Laurus azorica—A Dominant Tree in the Azorean Laurel Forest. Forests 2023, 14, 166. https://doi.org/10.3390/f14020166
Pavão DC, Jevšenak J, Silva LB, Elias RB, Silva L. Climate–Growth Relationships in Laurus azorica—A Dominant Tree in the Azorean Laurel Forest. Forests. 2023; 14(2):166. https://doi.org/10.3390/f14020166
Chicago/Turabian StylePavão, Diogo C., Jernej Jevšenak, Lurdes Borges Silva, Rui Bento Elias, and Luís Silva. 2023. "Climate–Growth Relationships in Laurus azorica—A Dominant Tree in the Azorean Laurel Forest" Forests 14, no. 2: 166. https://doi.org/10.3390/f14020166