
Forest Insect Outbreak Dynamics: Fractal Properties, Viscous Fingers, and Holographic Principle

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
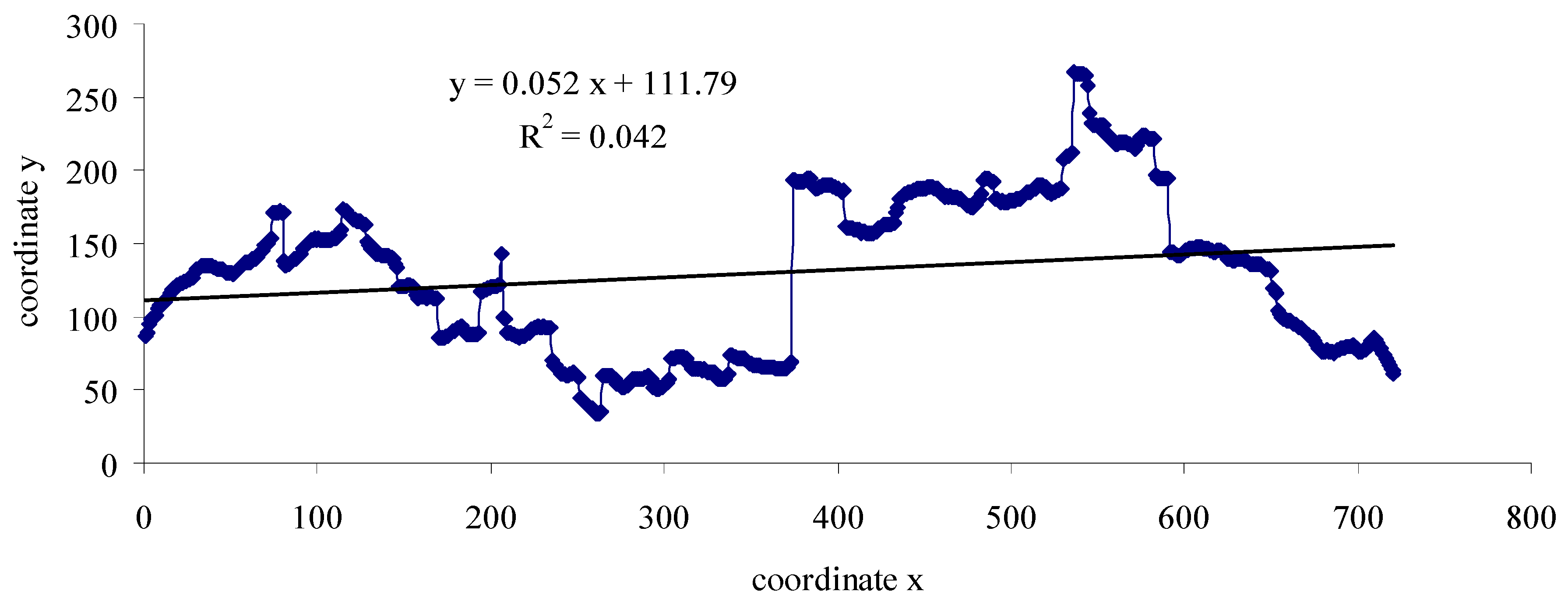
2.3. Data Analysis
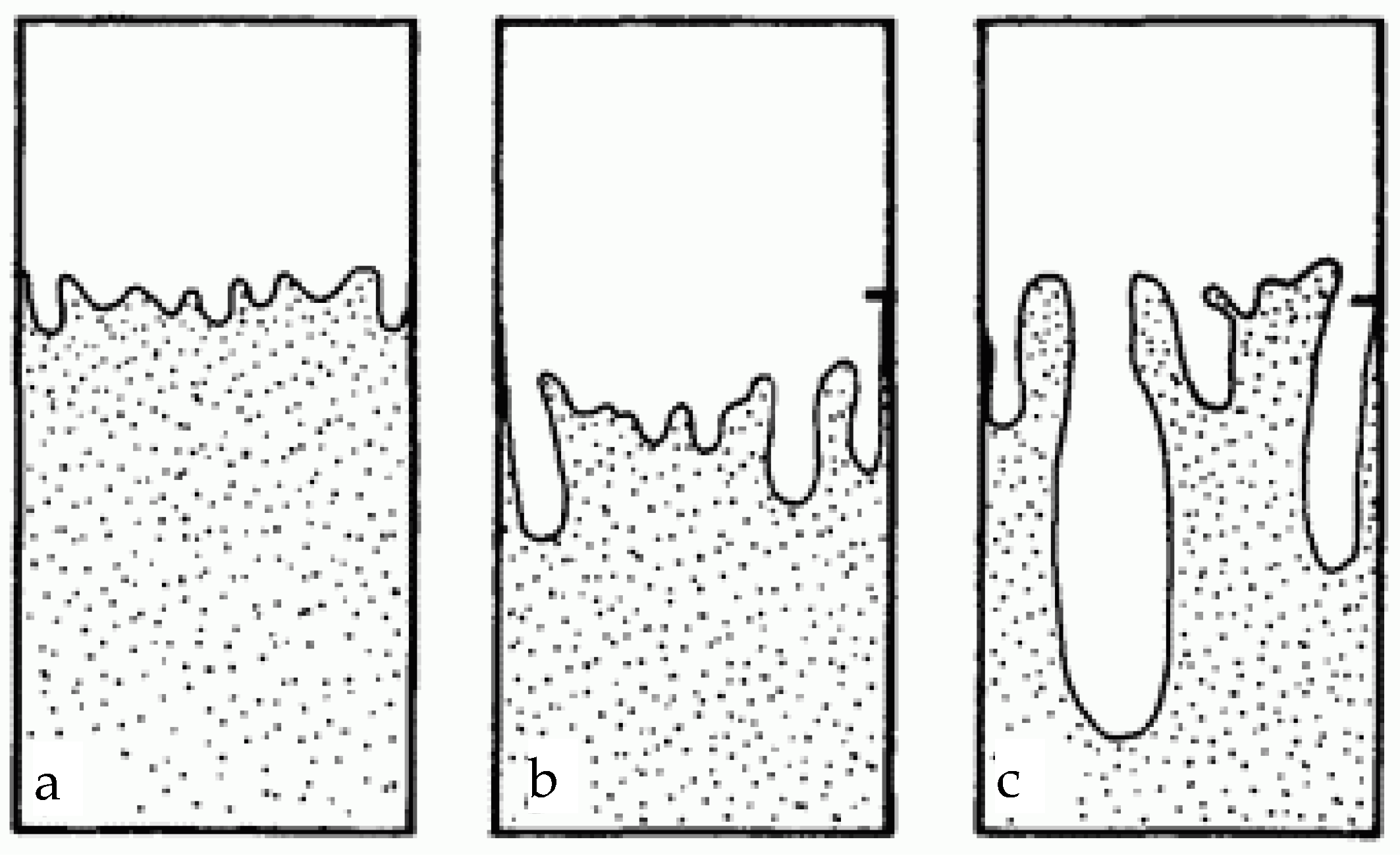
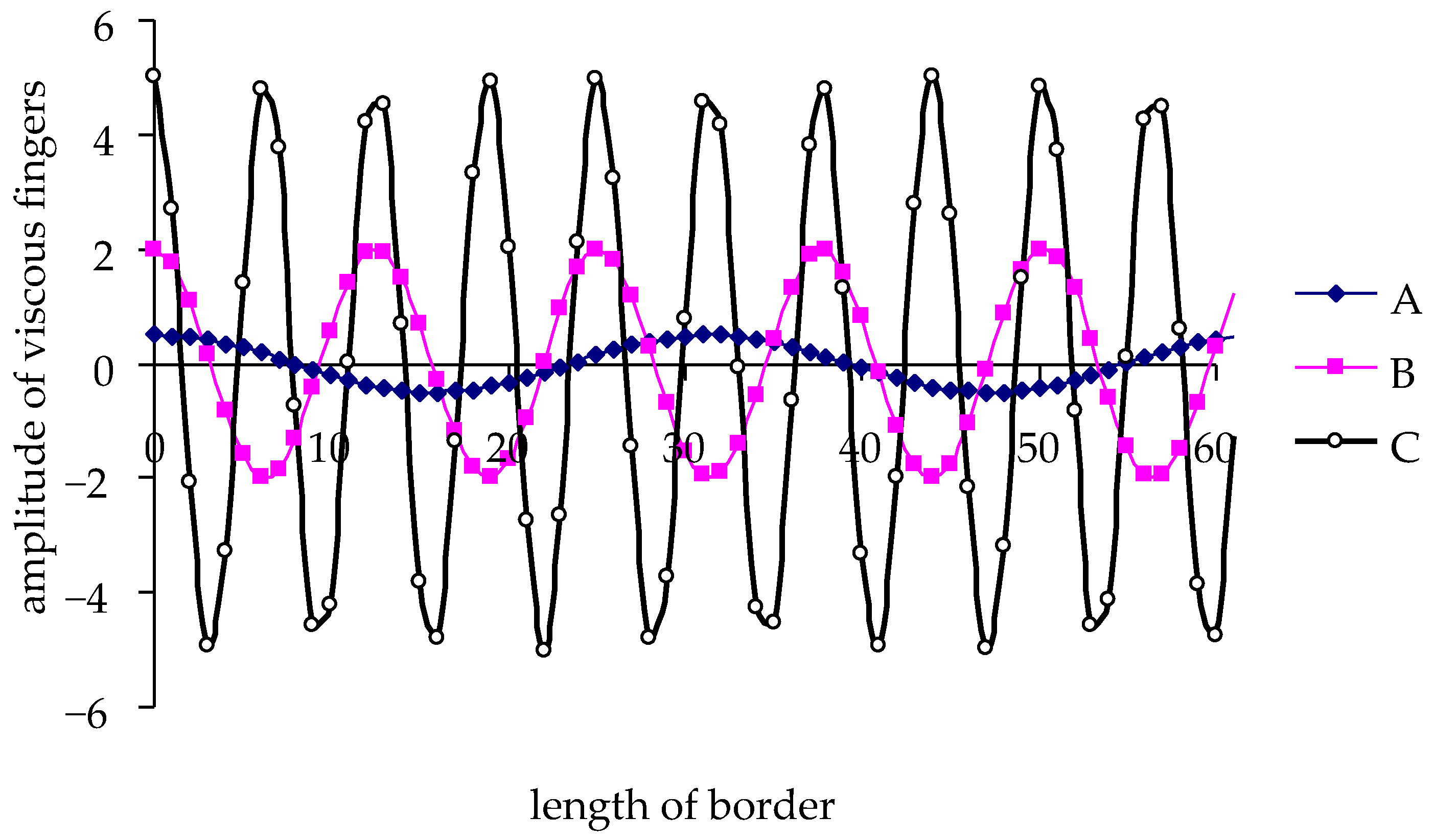
- (A) a boundary with a small number of “viscous fingers” and small deviations from the average characteristics of the interface (type a in Figure 1);
- (B) a boundary with an intermediate number of “viscous fingers” and moderate deviations from the average interface characteristics (type b in Figure 1);
- (C) a boundary with a large number of “viscous fingers” and strong deviations from the average characteristics of the interface (type c in Figure 1);
- (D) a boundary of random stationary shape.
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Isaev, A.S.; Khlebopros, R.G.; Kiselev, V.V.; Kondakov, Y.P.; Nedorezov, L.V.; Soukhovolsky, V.G. Forest Insects Population Dynamics; Publishing House of the Eurasian Entomological Journal: Novosibirsk, Russia, 2009; 115p. [Google Scholar]
- Isaev, A.S.; Soukhovolsky, V.G.; Tarasova, O.V.; Palnikova, E.N.; Kovalev, A.V. Forest Insect Population Dynamics, Outbreaks and Global Warming Effects; Wiley and Sons: New York, NY, USA, 2017; 286p. [Google Scholar]
- Taylor, L.R. Assessing and interpreting the spatial distributions of insect population. Ann. Rev. Entomol. 1984, 29, 321–357. [Google Scholar] [CrossRef]
- Sutton, A.; Tardif, J. Dendrochronological reconstruction of forest tent caterpillar outbreaks in time and space, western Manitoba, Canada. Can. J. For. Res. 2007, 37, 1643–1657. [Google Scholar] [CrossRef]
- Cooke, B.J.; Roland, J. Trembling aspen responses to drought and defoliation by forest tent caterpillar and reconstruction of recent outbreaks in Ontario. Can. J. For. Res. 2007, 37, 1586–1598. [Google Scholar] [CrossRef]
- Cooke, B.J.; MacQuarrie, C.J.K.; Lorenzetti, F. The dynamics of forest tent caterpillar outbreaks across east-central Canada. Ecography 2012, 35, 422–435. [Google Scholar] [CrossRef]
- Charbonneau, D.; Lorenzetti, F.; Doyon, F.; Mauffette, Y. The influence of stand and landscape characteristics on forest tent caterpillar (Malacosoma disstria) defoliation dynamics: The case of the 1999–2002 outbreak in northwestern Quebec. Can. J. For. Res. 2012, 42, 1827–1836. [Google Scholar] [CrossRef]
- Robert, L.E.; Sturtevant, B.R.; Cooke, B.J.; James, P.M.; Fortin, M.J.; Townsend, P.A.; Wolter, P.T.; Kneeshaw, D. Landscape host abundance and configuration regulate periodic outbreak behavior in spruce budworm (Choristoneura fumiferana Clem.). Ecography 2018, 41, 1556–1571. [Google Scholar] [CrossRef]
- Anderson, T.M.; Dragicevic, S. Network-agent based model for simulating the dynamic spatial network structure of complex ecological systems. Ecol. Model. 2018, 389, 19–32. [Google Scholar] [CrossRef]
- Barbour, D.A. Synchronous fluctuations in spatially separated populations of cyclic forest insects. In Population Dynamics of Forest Insects; Watt, A.D., Leather, S.R., Hunter, M.D., Kidd, N.A.C., Eds.; Intercept Limited: Andover, UK, 1990; pp. 339–346. [Google Scholar]
- Wolter, P.T.; Townsend, P.A.; Sturtevant, B.R.; Kingdon, C.C. Remote sensing of the distribution and abundance of host species for spruce budworm in Northern Minnesota and Ontario. Remote Sens. Environ. 2008, 112, 3971–3982. [Google Scholar] [CrossRef]
- Schroeder, M. Fractals, Chaos, and Power Laws; W.H. Freeman and Company: New York, NY, USA, 1991; 429p. [Google Scholar]
- Prozorov, S.S. The silk moth in fir forests of Siberia. Proc. SibLTI. Krasn. 1952, 3, 93–132. (In Russian) [Google Scholar]
- Boldaruyev, V.O. Population Dynamics of the Siberian Silk Moth and Its Parasites; Buryat Publishers: Ulan-Ude, Russia, 1969; 162p. (In Russian) [Google Scholar]
- Epova, V.I.; Pleshanov, A.S. Zones of Severity of Phyllophagous Insects in Asian Russia; Nauka: Novosibirsk, Russia, 1995; 147p. (In Russian) [Google Scholar]
- Kolomiyets, N.G. Parasites and Predators of the Siberian Silk Moth; Nauka: Novosibirsk, Russia, 1962; 172p. (In Russian) [Google Scholar]
- Kondakov, Y.P. Siberian silk moth outbreaks. In Population Ecology of Forest Animals in Siberia; Nauka: Novosibirsk, Russia, 1974; pp. 206–265. (In Russian) [Google Scholar]
- Rozhkov, A.S. Outbreak of the Siberian Silk Moth and Insect Control Measures; Nauka: Moscow, Russia, 1965; 178p. (In Russian) [Google Scholar]
- Rozhkov, A.S. Siberian Silk Moth; Nauka: Moscow, Russia, 1963; 175p. (In Russian) [Google Scholar]
- Soukhovolsky, V.G.; Tarasova, O.V.; Kovalev, A.V. A modeling of critical events in forest insects populations). Rus. J. Gen. Biol. 2020, 81, 374–386. (In Russian) [Google Scholar]
- Yurchenko, G.I.; Turova, G.I. Siberian and White-Striped Silkworms in the Far East; DalNIILCH Puplishers: Habarovsk, Russia, 2007; 98p. (In Russian) [Google Scholar]
- Pavlov, I.; Litovka, Y.; Golubev, D.; Astapenko, S.; Chromogin, P. New Outbreak of Dendrolimus sibiricus Tschetv. in Siberia (2012–2017): Monitoring, Modeling and Biological Control. Contemp. Probl. Ecol. 2018, 11, 406–419. [Google Scholar] [CrossRef]
- Broadbent, S.R.; Hammersley, J.M. Percolation processes. I. Crystals and mazes. Proc. Camb. Philos. Soc. 1957, 53, 629–641. [Google Scholar] [CrossRef]
- Kesten, H. Percolation Theory for Mathematicians; Birkhauser: Boston, MA, USA, 1982; 423p. [Google Scholar]
- Feder, J. Fractals; Plenum Press: New York, NY, USA, 1988; 283p. [Google Scholar]
- Jensen, I. Enumerations of lattice animals and trees. J. Stat. Phys. 2001, 102, 865–881. [Google Scholar] [CrossRef]
- Hsu, H.P.; Nadler, W.; Grassberger, P. Simulations of lattice animals and trees. J. Phys. A Math. Gen. 2005, 38, 775–806. [Google Scholar] [CrossRef]
- Kleman, M.; Lavrentovich, O.D. Soft Matter Physics: An Introduction; Springer: New York, NY, USA, 2003; 637p. [Google Scholar]
- Kessler, D.A.; Levine, H. The theory of Saffman-Taylor fingers. Phys. Rev. A Gen. Phys. 1986, 33, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Constantin, P.; Pugh, M. Global solutions for small data to the Hele-Shaw problem. Appear. Nonlinearity 1993, 6, 393–416. [Google Scholar] [CrossRef]
- Ceniceros, H.; Hou, T.Y. The singular perturbation of surface tension in Hele-Shaw flows. J. Fluid Mech. 2000, 409, 251–272. [Google Scholar] [CrossRef]
- Hooft, G. Canonical Quantization of Gravitating Point Particles in 2 + 1 Dimensions. Class. Quantum Gravity 1993, 10, 1653–1664. [Google Scholar] [CrossRef]
- Susskind, L. The world as a hologram. J. Math. Phys. 1995, 36, 6377–6396. [Google Scholar] [CrossRef]
- Susskind, L. The Black Hole War; Little, Brown: New York, NY, USA, 2008; 470p. [Google Scholar]
- Anderson, T.W. Statistical Analysis of Time Series; Wiley: New York, NY, USA, 1971; 704p. [Google Scholar]
- Meddens, A.J.H.; Hicke, J.A.; Vierling, L.A.; Hudak, A.T. Evaluating methods to detect bark beetle-caused tree mortality using single-date and multi-date Landsat imagery. Remote Sens. Environ. 2013, 132, 49–58. [Google Scholar] [CrossRef]
- Anees, A.; Aryal, J. Near-Real Time Detection of Beetle Infestation in Pine Forests Using MODIS Data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 3713–3723. [Google Scholar] [CrossRef]
- Meigs, G.W.; Kennedy, R.E.; Cohen, W.B. A Landsat time series approach to characterize bark beetle and defoliator impacts on tree mortality and surface fuels in conifer forests. Remote Sens. Environ. 2011, 115, 3707–3718. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R.; Hostert, P. Remote sensing of forest insect disturbances: Current state and future directions. Int. J. Appl. Earth Obs. Geoinf. 2017, 60, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Latifi, H.; Dahms, T.; Beudert, B.; Heurich, M.; Kübert, C.; Dech, S. Synthetic RapidEye data used for the detection of area-based spruce tree mortality induced by bark beetles. GIScience Remote Sens. 2018, 55, 839–859. [Google Scholar] [CrossRef]
- Spruce, J.P.; Hicke, J.A.; Hargrove, W.W.; Grulke, N.E.; Meddens, A.J.H. Use of MODIS NDVI Products to Map Tree Mortality Levels in Forests Affected by Mountain Pine Beetle Outbreaks. Forests 2019, 10, 811. [Google Scholar] [CrossRef]
- Bárta, V.; Lukeš, P.; Homolová, L. Early detection of bark beetle infestation in Norway spruce forests of Central Europe using Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2021, 100, 102335. [Google Scholar] [CrossRef]
- Dash, J.P.; Watt, M.S.; Pearse, G.D.; Heaphy, M.; Dungey, H.S. Assessing very high-resolution UAV imagery for monitoring forest health during a simulated disease outbreak. ISPRS J. Photogramm. Remote Sens. 2017, 131, 1–14. [Google Scholar] [CrossRef]
- Näsi, R.; Honkavaara, E.; Blomqvist, M.; Lyytikäinen-Saarenmaa, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Holopainen, M. Remote sensing of bark beetle damage in urban forests at individual tree level using a novel hyperspectral camera from UAV and aircraft. Urban For. Urban Green 2018, 30, 72–83. [Google Scholar] [CrossRef]
- Klouček, T.; Komárek, J.; Surový, P.; Hrach, K.; Janata, P.; Vašíček, B. The Use of UAV Mounted Sensors for Precise Detection of Bark Beetle Infestation. Remote Sens. 2019, 11, 1561. [Google Scholar] [CrossRef]
- Gray, D.R.; MacKinnon, W.E. Outbreak patterns of the spruce budworm and their impact in Canada. For. Chronicle. 2006, 82, 550–561. [Google Scholar] [CrossRef]
- Foster, J.R.; Townsend, P.A.; Mladenoff, D.J. Spatial dynamics of gypsy moth defoliation outbreak and dependence of habitat characteristics. Landscape Ecol. 2013, 28, 1307–1320. [Google Scholar] [CrossRef]
- Meigs, G.W.; Kennedy, R.E.; Gray, A.N.; Gregory, M.J. Spatiotemporal dynamics of recent mountain pine beetle and western spruce budworm outbreaks across the Pacific Northwest Region, USA. For. Ecol. Manag. 2015, 339, 71–86. [Google Scholar] [CrossRef]
- Senf, C.; Campbell, E.M.; Pflugmacher, D.; Wulder, M.A.; Hostert, P. A multi-scale analysis of western spruce budworm outbreak dynamics. Landsc. Ecol. 2017, 32, 501–514. [Google Scholar] [CrossRef]
- Sproull, G.J.; Adamus, M.; Szewczyk, J.; Kersten, G.; Szwagrzyk, J. Fine-scale spruce mortality dynamics driven by bark beetle disturbance in Babia Góra National Park. Poland. Eur. J. For. Res. 2016, 135, 507–517. [Google Scholar] [CrossRef]
- Fernández, A.; Fort, H. Catastrophic phase transitions and early warnings in a spatial ecological model. J. Stat. Mech. Theor. Exp. 2009, P09014. [Google Scholar] [CrossRef]
- Lewis, M.A.; Nelson, W.; Xu, C. A structured threshold model for mountain pine beetle outbreak. Bull. Math. Biol. 2010, 72, 565–589. [Google Scholar] [CrossRef]
- Fahse, L.; Heurich, M. Simulation and analysis of outbreaks of bark beetle infestations and their management at the stand level. Ecol. Model. 2011, 222, 1833–1846. [Google Scholar] [CrossRef]
- Overbeck, M.; Schmidt, M. Modelling infestation risk of Norway spruce by Ips typographus (L.) in the Lower Saxon Harz Mountains (Germany). For. Ecol. Manag. 2012, 266, 115–125. [Google Scholar] [CrossRef]
- Kautz, M.; Dworschak, K.; Gruppe, A.; Schopf, R. Quantifying spatio-temporal dispersion of bark beetle infestations in epidemic and non-epidemic conditions. For. Ecol. Manag. 2011, 262, 598–608. [Google Scholar] [CrossRef]
- Stadelmann, G.; Bugmann, H.; Wermelinger, B.; Meier, F.; Bigler, C. A predictive framework to assess spatio-temporal variability of infestations by the European spruce bark beetle. Ecography 2013, 36, 1208–1217. [Google Scholar] [CrossRef]
- Kärvemo, S.; Van Boeckel, T.P.; Gilbert, M.; Grégoire, J.C.; Schroeder, M. Large-scale risk mapping of an eruptive bark beetle—Importance of forest susceptibility and beetle pressure. For. Ecol. Manag. 2014, 318, 158–166. [Google Scholar] [CrossRef]
- Seidl, R.; Müller, J.; Hothorn, T.; Bässler, C.; Heurich, M.; Kautz, M. Small beetle, large-scale drivers: How regional and landscape factors affect outbreaks of the european spruce bark beetle. J. Appl. Ecol. 2016, 53, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Urban, D.; Keitt, T. Landscape connectivity: A graph-theoretic perspective. Ecology 2001, 82, 1205–1218. [Google Scholar] [CrossRef]
- Riley, J.R. Remote sensing in entomology. Annu. Rev. Entomol. 1989, 34, 247–271. [Google Scholar] [CrossRef]
- Nansen, C.; Elliott, N. Remote sensing and reflectance profiling in entomology. Annu. Rev. Entomol. 2016, 61, 139–158. [Google Scholar] [CrossRef]
- Latchininsky, A.V. Locusts and remote sensing: A review. J. Appl. Remote Sens. 2013, 7, 075099. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, Y.; Pu, R.; Gonzalez-Moreno, P.; Yuan, L.; Wu, K.; Huang, W. Monitoring plant diseases and pests through remote sensing technology: A review. Comput. Electron. Agric. 2019, 165, 104943. [Google Scholar] [CrossRef]
- Filho, I.F.H.; Heldens, W.B.; Kong, Z.; de Lange, E.S. Drones: Innovative technology for use in precision pest management. J. Econ. Entomol. 2020, 113, 1–25. [Google Scholar] [CrossRef]
- Spruce, J.P.; Sader, S.; Ryan, R.E.; Smoot, J.; Kuper, P.; Ross, K.; Prados, D.; Russell, J.; Gasser, G.; McKellip, R.; et al. Assessment of MODIS NDVI time series data products for detecting forest defoliation by gypsy moth outbreaks. Remote Sens. Environ. 2011, 115, 427–437. [Google Scholar] [CrossRef]
- Olsson, P.O.; Lindström, J.; Eklundh, L. Near real-time monitoring of insect induced defoliation in subalpine birch forests with MODIS derived NDVI. Remote Sens. Environ. 2016, 181, 42–53. [Google Scholar] [CrossRef]
- Eklundh, L.; Johansson, T.; Solberg, S. Mapping insect defoliation in scots pine with MODIS time-series data. Remote Sens. Environ. 2009, 113, 1566–1573. [Google Scholar] [CrossRef]
- Hart, S.J.; Veblen, T.T. Detection of spruce beetle-induced tree mortality using high- and medium-resolution remotely sensed imagery. Remote Sens. Environ. 2015, 168, 134–145. [Google Scholar] [CrossRef]
- Bryk, M.; Kołodziej, B.; Pliszka, R. Changes of Norway spruce health in the Białowieża forest (CE Europe) in 2013–2019 during a bark beetle infestation, studied with Landsat imagery. Forests 2021, 12, 34. [Google Scholar] [CrossRef]
- de Beurs, K.M.; Townsend, P.A. Estimating the effect of gypsy moth defoliation using MODIS. Remote Sens. Environ. 2008, 112, 3983–3990. [Google Scholar] [CrossRef]
- Zhou, G.; Liebhold, A.M. Forecasting the spatial dynamics of gypsy moth outbreaks using cellular transition models. Landsc. Ecol. 1995, 10, 177–189. [Google Scholar] [CrossRef]
- Logan, J.A.; White, P.; Bentz, B.J.; Powell, J.A. Model analysis of spatial patterns in mountain pine beetle outbreaks. Theor. Popul. Biol. 1998, 53, 236–255. [Google Scholar] [CrossRef]
- Fisher, R.A. The Wave of Advance of Advantageous Genes. Ann. Eugen. 1937, 7, 355–369. [Google Scholar] [CrossRef]
- Kolmogorov, A.N.; Petrovsky, I.G.; Piskunov, N.S. A study of the equation of diffusion linked to the matter increase and its application to one biological problem. Bull. MSU Ser. A Math. Mech. 1937, 16, 1–16. (In Russian) [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Border’s Form | Parameters | |
|---|---|---|
| s | Fmax | |
| A | 0.354 | 0.032 |
| B | 1.420 | 0.081 |
| C | 3.571 | 0.161 |
| D1 | 0.263 | 0.339 |
| D2 | 0.897 | 0.258 |
| Outbreak | Variables | Coeff. | Std. Err. | t-Test | p-Value |
|---|---|---|---|---|---|
| Yeniseisk district, 2015. | ln s0 | −0.587 | 0.115 | −5.115 | 0.000 |
| α | 1.330 | 0.024 | 55.705 | 0.000 | |
| adjR2 | 0.979 | ||||
| F | 3103 | ||||
| D | 1.50 | ||||
| Yeniseisk district, 2016. | ln s0 | −0.788 | 0.046 | −17.311 | 0.000 |
| α | 1.362 | 0.011 | 122.561 | 0.000 | |
| adjR2 | 0.992 | ||||
| F | 15,021 | ||||
| D | 1.47 | ||||
| Irbey district, 2020. | ln s0 | −0.844 | 0.033 | −25.653 | 0.00 |
| α | 1.428 | 0.0084 | 169.523 | 0.00 | |
| adjR2 | 0.99 | ||||
| F | 28,737 | ||||
| D | 1.40 | ||||
| Irbey district, 2020. | ln s0 | −0.701 | 0.026 | −27.447 | 0.00 |
| α | 1.374 | 0.007 | 194.701 | 0.00 | |
| adjR2 | 0.986 | ||||
| F | 37,908 | ||||
| D | 1.455 | ||||
| Parameter | Year | |||
|---|---|---|---|---|
| 2018 | 2019 | 2020 | 2021 | |
| Frequency of the spectrum maximum, fmax, Hz | 0 | 0.0077 | 0.0031 | 0.0002 |
| Standard deviation σ of “viscous fingers” pixels | 0 | 57, 2 | 37.5 | 20.2 |
| Area S of outbreaks in % relative to the total area of the territory | 0 | 17, 52 | 45.56 | 47.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soukhovolsky, V.; Kovalev, A.; Tarasova, O.; Ivanova, Y. Forest Insect Outbreak Dynamics: Fractal Properties, Viscous Fingers, and Holographic Principle. Forests 2023, 14, 2459. https://doi.org/10.3390/f14122459
Soukhovolsky V, Kovalev A, Tarasova O, Ivanova Y. Forest Insect Outbreak Dynamics: Fractal Properties, Viscous Fingers, and Holographic Principle. Forests. 2023; 14(12):2459. https://doi.org/10.3390/f14122459
Chicago/Turabian StyleSoukhovolsky, Vladislav, Anton Kovalev, Olga Tarasova, and Yulia Ivanova. 2023. "Forest Insect Outbreak Dynamics: Fractal Properties, Viscous Fingers, and Holographic Principle" Forests 14, no. 12: 2459. https://doi.org/10.3390/f14122459