
Inverted Soil Mounding as a Restoration Approach of Seismic Lines in Boreal Peatlands: Implications on Plant and Arthropod Abundance and Diversity

Abstract
:1. Introduction
2. Materials and Methods
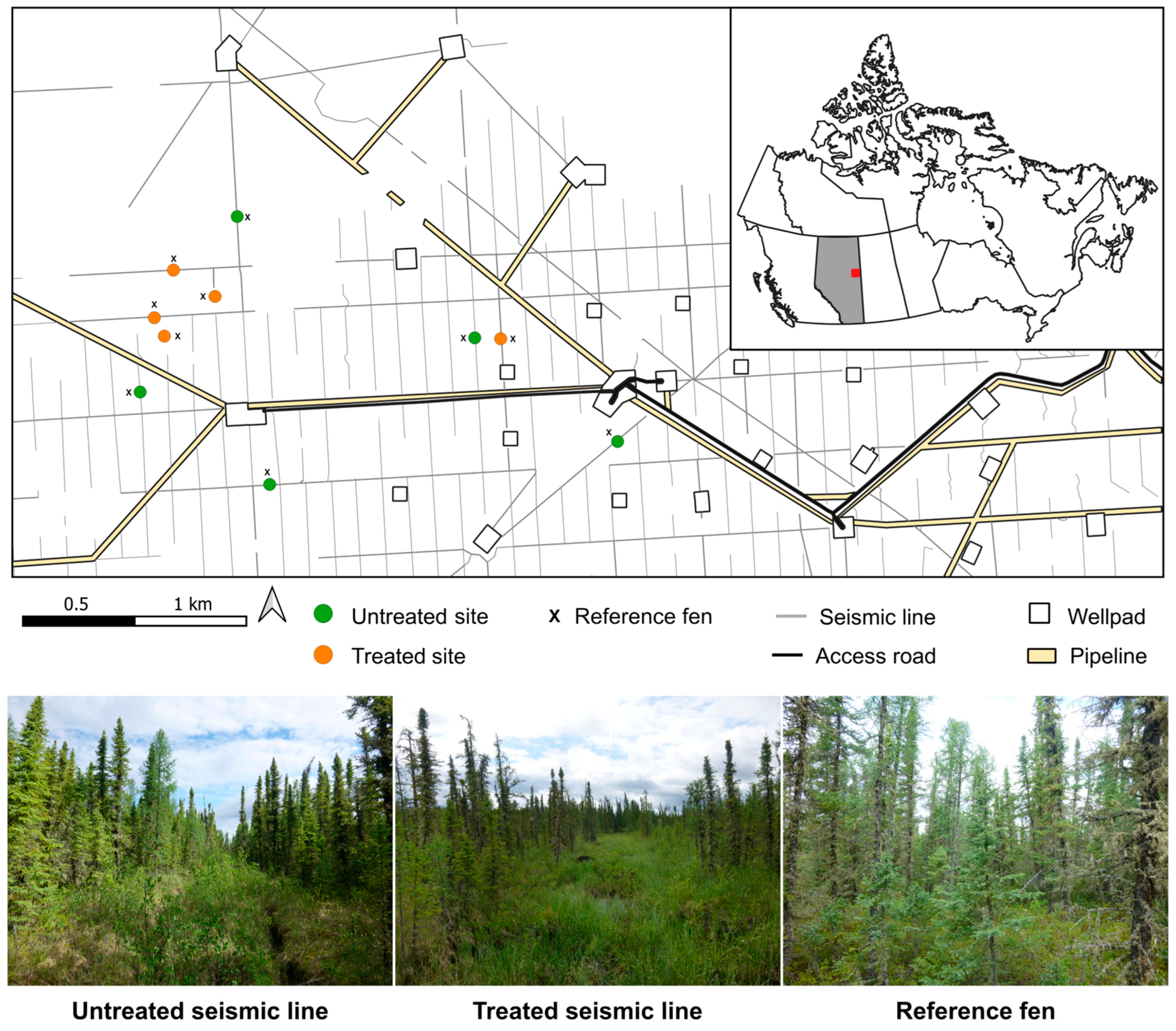
2.1. Study Area
2.2. Data Collection
2.2.1. Understory Vegetation
2.2.2. Ground-Dwelling Arthropods
2.3. Data Analyses
2.3.1. Richness, Diversity, and Abundance
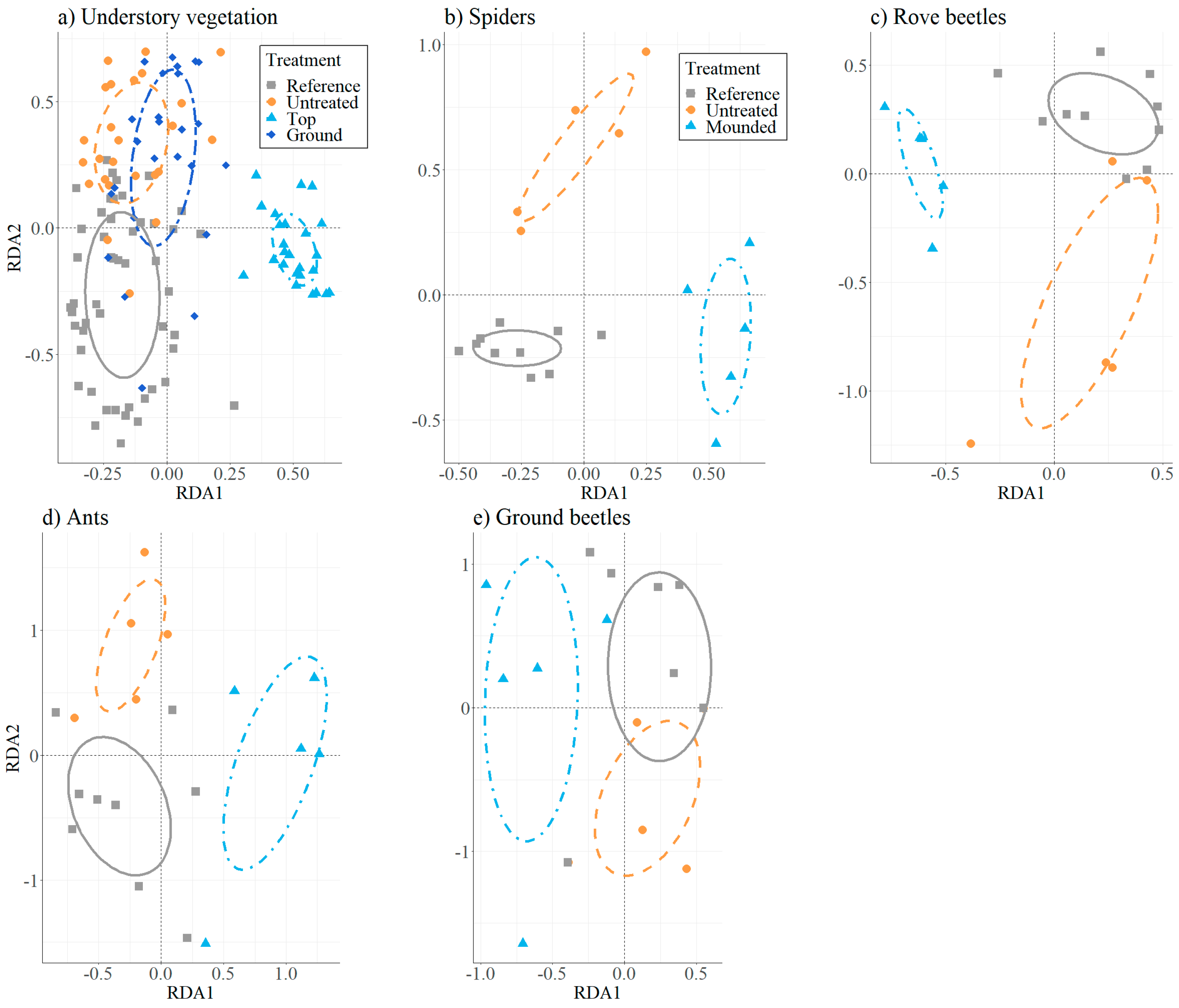
2.3.2. Species Composition and Indicator Species
3. Results
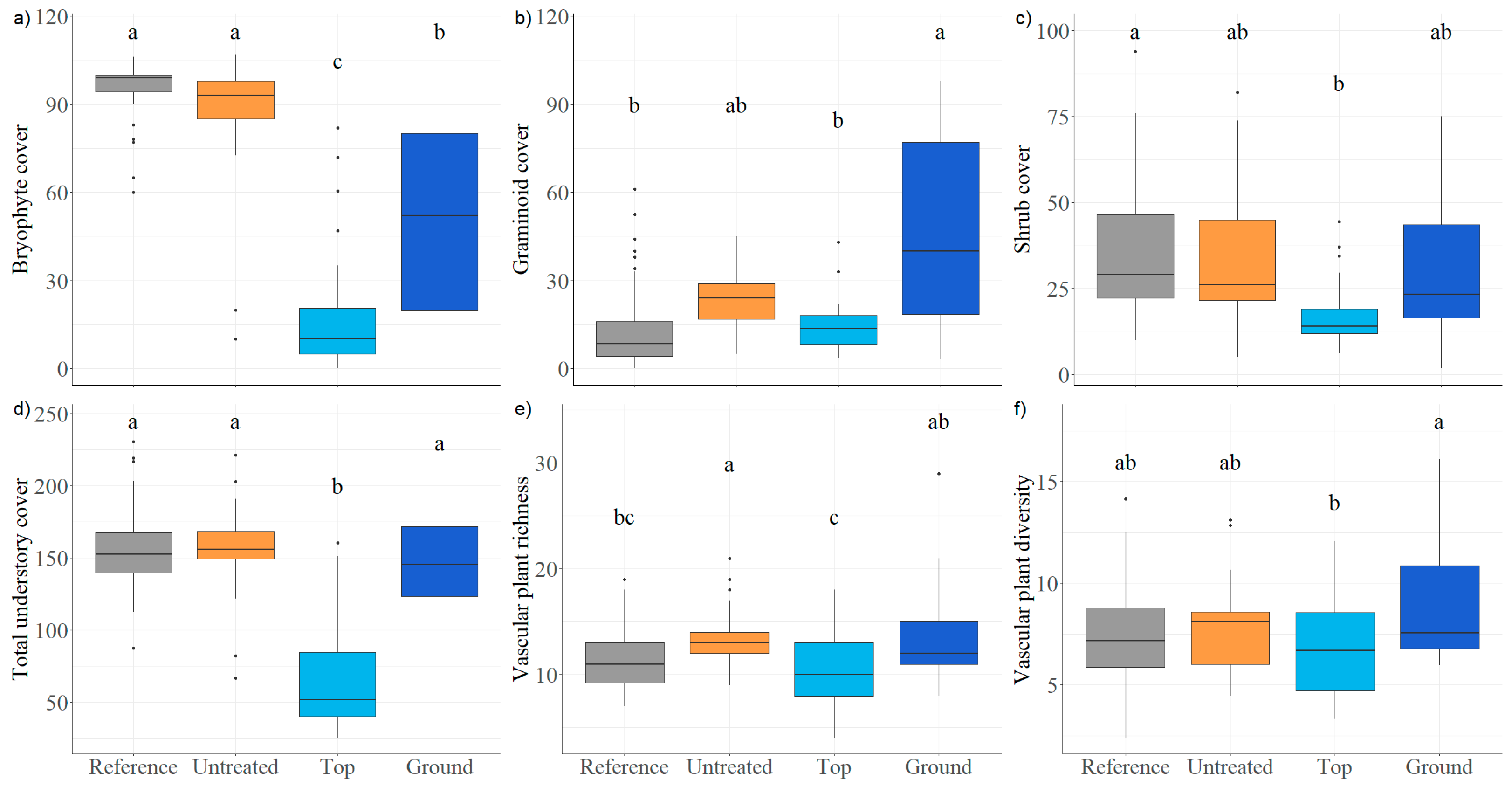
3.1. Understory Vegetation
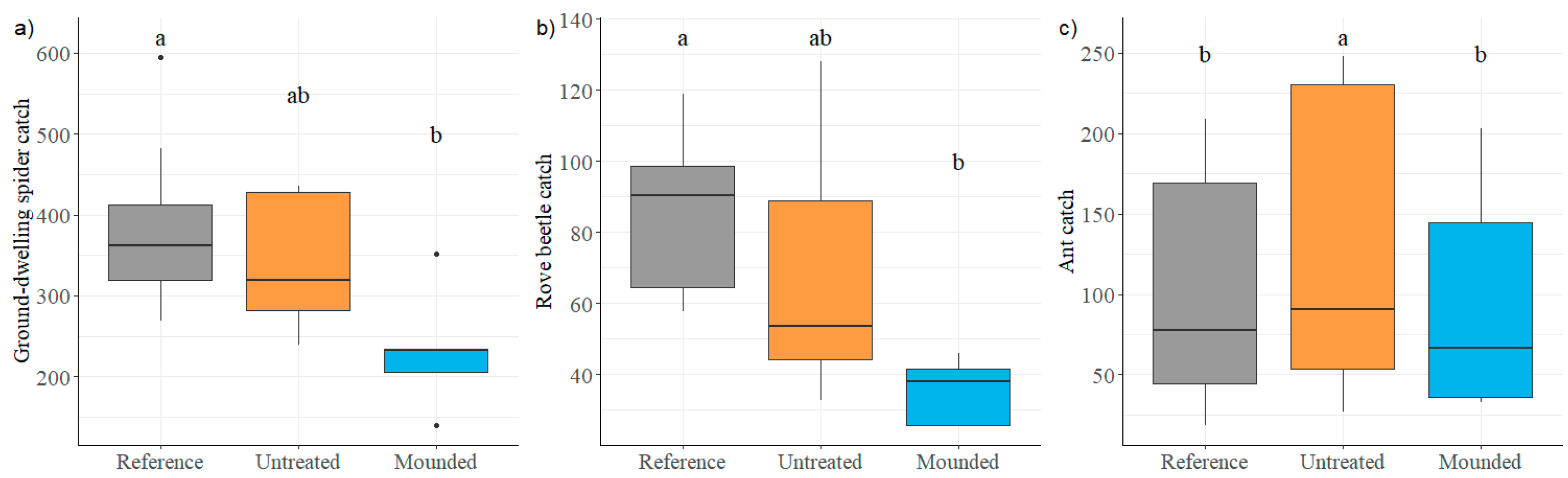
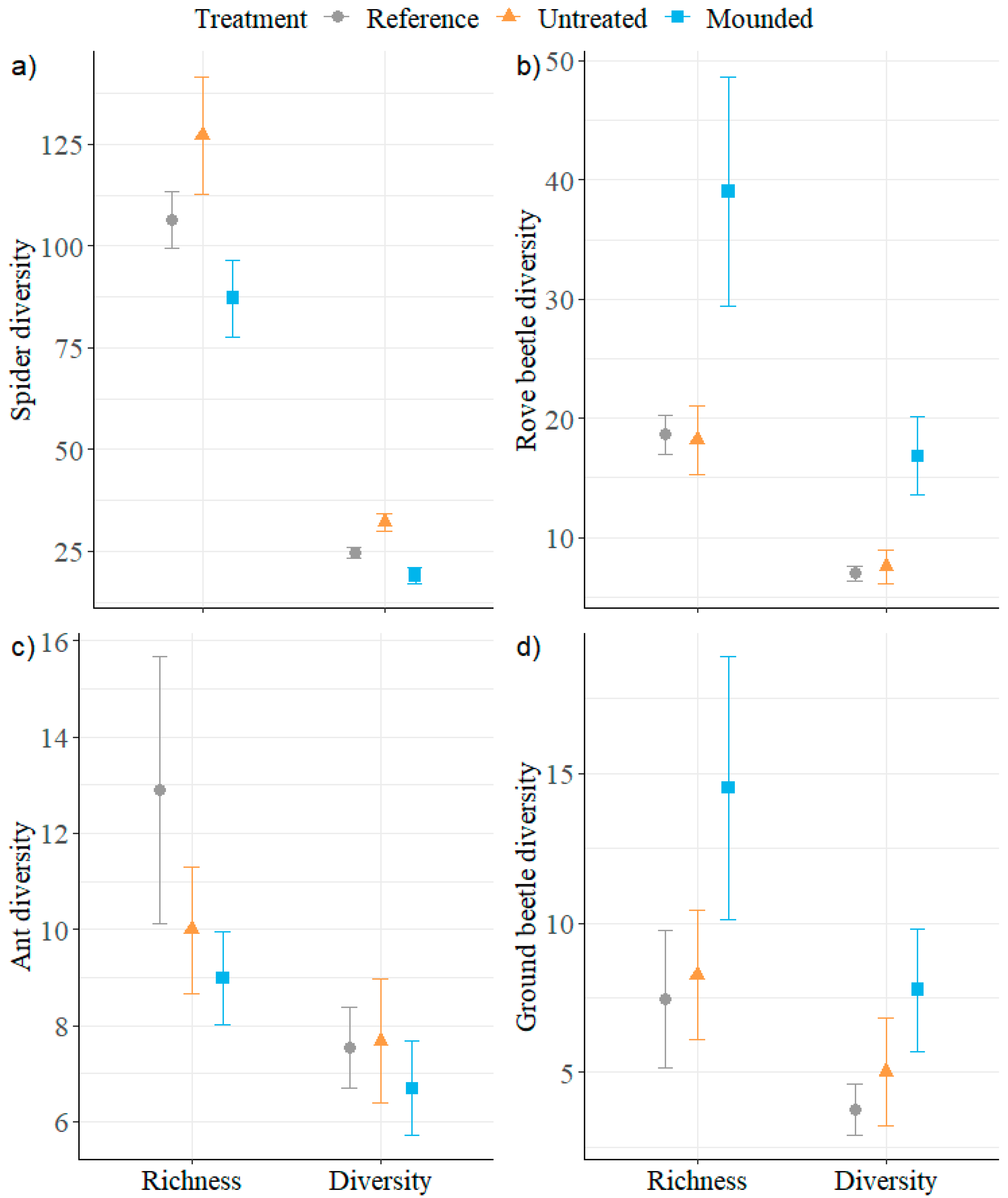
3.2. Ground-Dwelling Arthropods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rydin, H.; Jeglum, J.K. The Biology of Peatlands, 2nd ed.; Oxford University Press: Oxford, UK, 2013; p. 432. [Google Scholar]
- Turetsky, M.R.; St. Louis, V.L. Disturbance in Boreal Peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 359–379. [Google Scholar]
- van Rensen, C.K.; Nielsen, S.E.; White, B.; Vinge, T.; Lieffers, V.J. Natural regeneration of forest vegetation on legacy seismic lines in boreal habitats in Alberta’s oil sands region. Biol. Conserv. 2015, 184, 127–135. [Google Scholar] [CrossRef]
- Lee, P.; Boutin, S. Persistence and developmental transition of wide seismic lines in the western Boreal Plains of Canada. J. Environ. Manag. 2006, 78, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, C.J.; Filicetti, A.T.; Nielsen, S.E. High precision altimeter demonstrates simplification and depression of microtopography on seismic lines in treed peatlands. Forests 2019, 10, 295. [Google Scholar] [CrossRef]
- Filicetti, A.T.; Nielsen, S.E. Tree regeneration on industrial linear disturbances in treed peatlands is hastened by wildfire and delayed by loss of microtopography. Can. J. For. Res. 2020, 50, 936–945. [Google Scholar] [CrossRef]
- Government of Canada. Species at Risk Act: An Act Respecting the Protection of Wildlife Species at Risk in Canada; Canada Gazette Part III: Ottawa, ON, Canada, 2002; Volume 25, pp. 1–104. [Google Scholar]
- Wasser, S.K.; Keim, J.L.; Taper, M.L.; Lele, S.R. The influences of wolf predation, habitat loss, and human activity on caribou and moose in the Alberta oil sands. Front. Ecol. Environ. 2011, 9, 546–551. [Google Scholar] [CrossRef]
- Nagy-Reis, M.; Dickie, M.; Calvert, A.M.; Hebblewhite, M.; Hervieux, D.; Seip, D.R.; Gilbert, S.L.; Venter, O.; DeMars, C.; Boutin, S.; et al. Habitat loss accelerates for the endangered woodland caribou in western Canada. Conserv. Sci. Pract. 2021, 3, e437. [Google Scholar] [CrossRef]
- Latham, A.D.M.; Latham, M.C.; Boyce, M.S.; Boutin, S. Movement responses by wolves to industrial linear features and their effect on woodland caribou in northeastern Alberta. Ecol. Appl. 2011, 21, 2854–2865. [Google Scholar] [CrossRef]
- Pyper, M.P.; Nishi, J.; McNeil, L. Linear Feature Restoration in Caribou Habitat: A Summary of Current Practices and a Roadmap for Future Programs; FUSE Consulting Ltd.: Edmonton, AB, Canada, 2014. [Google Scholar]
- Filicetti, A.T.; Cody, M.; Nielsen, S.E. Caribou conservation: Restoring trees on seismic lines in Alberta, Canada. Forests 2019, 10, 185. [Google Scholar] [CrossRef]
- Londo, A.J.; Mroz, G.D. Bucket Mounding as a Mechanical Site Preparation Technique in Wetlands. North. J. Appl. For. 2001, 18, 7–13. [Google Scholar] [CrossRef]
- Sutton, R.F. Mounding site preparation: A review of European and North American experience. New For. 1993, 7, 151–192. [Google Scholar] [CrossRef]
- Kleinke, K.; Davidson, S.J.; Schmidt, M.; Xu, B.; Strack, M. How mounds are made matters: Seismic line restoration techniques affect peat physical and chemical properties throughout the peat profile. Can. J. For. Res. 2022, 52, 963–976. [Google Scholar] [CrossRef]
- Echiverri, L.F.I.; Macdonald, S.E.; Nielsen, S.E. Disturbing to restore? Effects of mounding on understory communities on seismic lines in treed peatlands. Can. J. For. Res. 2020, 50, 1340–1351. [Google Scholar] [CrossRef]
- Echiverri, L.F.I.; Macdonald, S.E.; Nielsen, S.E. Mounding treatments set back bryophyte recovery on linear disturbances in treed peatlands. Restor. Ecol. 2023, 31, e13813. [Google Scholar] [CrossRef]
- Pinzon, J.; Dabros, A.; Hoffman, P. Soil mounding as a restoration approach of seismic lines in boreal peatlands: Implications on microtopography. Restor. Ecol. 2022, e13835. [Google Scholar] [CrossRef]
- Caners, R.T.; Crisfield, V.; Lieffers, V.J. Habitat heterogeneity stimulates regeneration of bryophytes and vascular plants on disturbed minerotrophic peatlands. Can. J. For. Res. 2019, 49, 281–295. [Google Scholar] [CrossRef]
- Görn, S.; Fischer, K. Measuring the efficiency of fen restoration on carabid beetles and vascular plants: A case study from north-eastern Germany. Restor. Ecol. 2015, 23, 413–420. [Google Scholar] [CrossRef]
- Hoffmann, H.; Kleeberg, A.; Görn, S.; Fischer, K. Riverine fen restoration provides secondary habitat for endangered and stenotopic rove beetles (Coleoptera: Staphylinidae). Insect Conserv. Divers. 2018, 11, 194–203. [Google Scholar] [CrossRef]
- Haase, H.; Balkenhol, B. Spiders (Araneae) as subtle indicators for successional stages in peat bogs. Wetl. Ecol. Manag. 2015, 23, 453–466. [Google Scholar] [CrossRef]
- Lehmitz, R.; Haase, H.; Otte, V.; Russell, D. Bioindication in peatlands by means of multi-taxa indicators (Oribatida, Araneae, Carabidae, Vegetation). Ecol. Indic. 2020, 109, 105837. [Google Scholar] [CrossRef]
- Štokmane, M.; Spuņģis, V. The influence of vegetation structure on spider species richness, diversity and community organization in the Apšuciems calcareous fen, Latvia. Anim. Biodivers. Conserv. 2016, 39, 16. [Google Scholar] [CrossRef]
- Glime, J.M.; Lissner, J. Arthropods: Spiders. In Bryophyte Ecology; Glime, J.M., Ed.; Michigan Technological University and the International Association of Bryologists: E-book; Bryological Interaction: Houghton, MI, USA, 2017; Volume 2, pp. 7-4-2–7-4-31. [Google Scholar]
- Främbs, H. The importance of habitat structure and food supply for carabid beetles (Coleoptera, Carabidae) in peat bogs. Mem. Entomol. Soc. Can. 1994, 126, 145–159. [Google Scholar] [CrossRef]
- Davidson, S.J.; Goud, E.M.; Franklin, C.; Nielsen, S.E.; Strack, M. Seismic line disturbance alters soil physical and chemical properties across boreal forest and peatland soils. Front. Earth Sci. 2020, 8, 281. [Google Scholar] [CrossRef]
- Braverman, M.; Quinton, W.L. Hydrological impacts of seismic lines in the wetland-dominated zone of thawing, discontinuous permafrost, Northwest Territories, Canada. Hydrol. Process. 2016, 30, 2617–2627. [Google Scholar] [CrossRef]
- Siira-Pietkainen, A.S.; Haimi, J.; Siitonen, J. Short-term responses of soil macroarthropod community to clear felling and alternative forest regeneration methods. For. Ecol. Manag. 2003, 172, 339–353. [Google Scholar] [CrossRef]
- Brouillet, L.; Desmet, P.; Coursol, F.; Meades, S.J.; Favreau, M.; Anions, M.; Bélisle, P.; Gendreau, C.; Shorthouse, D. VASCAN: The Database of Vascular Plants of Canada. Available online: http://data.canadensys.net/vascan/ (accessed on 19 July 2021).
- Integrated Taxonomic Information System. Available online: http://www.itis.gov (accessed on 6 September 2022).
- Spence, J.R.; Niemelä, J.K. Sampling carabid assemblages with pitfall traps—The madness and the method. Can. Entomol. 1994, 126, 881–894. [Google Scholar] [CrossRef]
- World Spider Catalog, Version 23.5, Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 6 September 2022).
- Lindroth, C.H. The Ground-Beetles (Carabidae, Excl. Cicindelinae) of Canada and Alaska; Opuscula Entomologica. Supplementum 1961–1969; Entomologiska Sällskapet: Lund, Sweden, 1963. [Google Scholar]
- Newton, A.F.; Thayer, M.K.; Ashe, J.S.; Chandler, D.S. Family 22. Staphylinidae Latreille, 1802. In American Beetles: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia; CRC Press: Boca Raton, FL, USA, 2001; Volume 1, pp. 272–418. [Google Scholar]
- Bousquet, Y.; Bouchard, P.; Davies, A.E.; Sikes, D.S. Checklist of Beetles (Coleoptera) of Canada and Alaska. ZooKeys 2013, 360, 1–44. [Google Scholar] [CrossRef]
- Bolton, B. A New General Catalogue of the Ants of the World; Harvard University Press: Cambridge, UK, 1995. [Google Scholar]
- Ellison, A.M.; Gotelli, N.J.; Farnswoth, E.J.; Alpert, G. A Field Guide to the Ants of New England; Yale University Press: New Haven, CT, USA, 2012; p. 388. [Google Scholar]
- Glasier, J.R.N.; Acorn, J.H.; Nielsen, S.; Proctor, H.C. Ants (Hymenoptera: Formicidae) of Alberta: A key to species based primarily on the worker caste. Can. J. Arthropod Identif. 2013, 22, 1–104. [Google Scholar] [CrossRef]
- Chao, A.; Chiu, C.-H.; Jost, L. Unifying species diversity, phylogenetic diversity, functional Diversity, and related similarity and differentiation measures through Hill numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent Alpha and Beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, 4.1.1; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: Linear and Nonlinear Mixed Effects Models, 3.1-152; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Lenth, R. emmeans: Estimated Marginal Means, Aka Least-Squares Means, 1.7.0; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: iNterpolation and EXTrapolation for Species Diversity, 2.0.20; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, p. 1006. [Google Scholar]
- Rao, C.R. The use and interpretation of principal component analysis in applied research. Sankhyā Indian J. Stat. Ser. A 1964, 26, 329–358. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology package, 2.5-7; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005. [Google Scholar]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Takyi, S.K.; Hillman, G.R. Growth of Coniferous Seedlings on a Drained and Mounded Peatland in Central Alberta. North. J. Appl. For. 2000, 17, 71–79. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Caners, R.T.; Ge, H. Re-establishment of hummock topography promotes tree regeneration on highly disturbed moderate-rich fens. J. Environ. Manag. 2017, 197, 258–264. [Google Scholar] [CrossRef]
- Klimaszewski, J.; Bernier-Cardou, M.; Germain, C. The effects of forestry practices on the abundance of arthropods (Acarina, Araneae, Collembola, Coleoptera and Diptera). Belg. J. Entomol. 2003, 5, 103–116. [Google Scholar]
- Pearce, J.L.; Venier, L.A. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as bioindicators of sustainable forest management: A review. Ecol. Indic. 2006, 6, 780–793. [Google Scholar] [CrossRef]
- Pinzon, J.; Spence, J.R.; Langor, D.W.; Shorthouse, D.P. Ten-year responses of ground-dwelling spiders to retention harvest in the boreal forest. Ecol. Appl. 2016, 26, 2579–2597. [Google Scholar] [CrossRef]
- Venier, L.A.; Work, T.T.; Klimaszewski, J.; Morris, D.M.; Bowden, J.J.; Kwiaton, M.M.; Webster, K.; Hazlett, P. Ground-dwelling arthropod response to fire and clearcutting in Jack Pine: Implications for ecosystem management. Can. J. For. Res. 2017, 47, 1614–1631. [Google Scholar] [CrossRef]
- Wu, L.; Pinzon, J. Response of rove-beetle (Staphylinidae) assemblages to the cumulative effect of wildfire and linear footprint in boreal treed peatlands. Ecol. Evol. 2022, 12, e9564. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-I.; Langor, D.W.; Spence, J.R.; Pinzon, J.; Pohl, G.R.; Hartley, D.J.; Work, T.T.; Wu, L. Rapid recovery of boreal rove beetle (Staphylinidae) assemblages 16 years after variable retention harvest. Ecography 2023, 2023, e06347. [Google Scholar] [CrossRef]
- Pohl, G.R.; Langor, D.W.; Spence, J.R. Rove beetles and ground beetles (Coleoptera: Staphylinidae, Carabidae) as indicators of harvest and regeneration practices in western Canadian foothills forests. Biol. Conserv. 2007, 137, 294–307. [Google Scholar] [CrossRef]
- Andersen, A.N. Responses of ant communities to disturbance: Five principles for understanding the disturbance dynamics of a globally dominant faunal group. J. Anim. Ecol. 2019, 88, 350–362. [Google Scholar] [CrossRef]
- Philpott, S.M.; Perfecto, I.; Ambrecht, I.; Parr, C.L. Ant Diversity and Function in Disturbed and Changing Habitats. In Ant Ecology; Oxford University Press: New York, NY, USA, 2010; pp. 137–156. [Google Scholar]
- Pinzon, J.; Dabros, A.; Riva, F.; Glasier, J.R.N. Short-term effects of wildfire in boreal peatlands: Does fire mitigate the linear footprint of oil and gas exploration? Ecol. Appl. 2021, 31, e02281. [Google Scholar] [CrossRef] [PubMed]
- Pearce, J.L.; Venier, L.A.; Eccles, G.; Pedlar, J.; McKenney, D. Influence of habitat and microhabitat on epigeal spider (Araneae) assemblages in four stand types. Biodivers. Conserv. 2004, 13, 1305–1334. [Google Scholar] [CrossRef]
- Buddle, C.M.; Langor, D.W.; Pohl, G.R.; Spence, J.R. Arthropod responses to harvesting and wildfire: Implications for emulation of natural disturbance in forest management. Biol. Conserv. 2006, 128, 346–357. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Variance Explained | Treatment | |||||
|---|---|---|---|---|---|---|---|
| Adj. R2 | RDA 1 | RDA 2 | F | p Value | Contrast | p Value | |
| Understory plants | 0.17 | 0.13 | 0.05 | 9.5 | 0.001 | ||
| Reference–untreated | 0.001 | ||||||
| Reference–top | 0.001 | ||||||
| Reference–ground | 0.001 | ||||||
| Untreated–top | 0.001 | ||||||
| Untreated–ground | 0.015 | ||||||
| Top–ground | 0.001 | ||||||
| Ground-dwelling spiders | 0.2 | 0.24 | 0.05 | 3.4 | 0.001 | ||
| Reference–untreated | 0.100 | ||||||
| Reference–mounded | 0.001 | ||||||
| Untreated–mounded | 0.023 | ||||||
| Rove beetles | 0.19 | 0.23 | 0.04 | 3.2 | 0.001 | ||
| Reference–untreated | 0.390 | ||||||
| Reference–mounded | 0.001 | ||||||
| Untreated–mounded | 0.029 | ||||||
| Ants | 0.16 | 0.2 | 0.05 | 2.8 | 0.001 | ||
| Reference–untreated | 0.446 | ||||||
| Reference–mounded | 0.001 | ||||||
| Untreated–mounded | 0.013 | ||||||
| Ground beetles | 0.09 | 0.16 | 0.02 | 1.9 | 0.026 | ||
| Reference–untreated | 0.816 | ||||||
| Reference–mounded | 0.014 | ||||||
| Untreated–mounded | 0.100 | ||||||
| Treatment | Species | Specificity | Fidelity | Indicator Value | p Value |
|---|---|---|---|---|---|
| Understory vegetation | |||||
| Reference | Rhododendron groenlandicum | 0.75 | 0.64 | 0.69 | 0.001 |
| Vaccinium oxycoccos | 0.46 | 0.92 | 0.65 | 0.001 | |
| Maianthemum trifolium | 0.35 | 0.96 | 0.58 | 0.004 | |
| Chamaedaphne calyculata | 0.80 | 0.30 | 0.49 | 0.002 | |
| Rubus chamaemorus | 0.82 | 0.28 | 0.48 | 0.007 | |
| Vaccinium vitis-idaea | 0.54 | 0.42 | 0.48 | 0.005 | |
| Untreated | Andromeda polifolia | 0.49 | 0.80 | 0.62 | 0.001 |
| Menyanthes trifoliata | 0.62 | 0.52 | 0.57 | 0.002 | |
| Betula pumila | 0.39 | 0.80 | 0.56 | 0.037 | |
| Carex chordorrhiza | 0.37 | 0.72 | 0.52 | 0.026 | |
| Drosera rotundifolia | 0.45 | 0.52 | 0.48 | 0.016 | |
| Pedicularis parviflora | 0.72 | 0.24 | 0.42 | 0.005 | |
| Salix planifolia | 0.65 | 0.20 | 0.36 | 0.024 | |
| Salix bebbiana | 0.72 | 0.16 | 0.34 | 0.045 | |
| Mounded | Picea mariana (understory) | 0.73 | 1.00 | 0.86 | 0.001 |
| (Top) | Populus tremuloides (understory) | 0.94 | 0.76 | 0.85 | 0.001 |
| Chamaenerion angustifolium | 0.67 | 0.40 | 0.52 | 0.003 | |
| Salix spp. | 1.00 | 0.20 | 0.45 | 0.001 | |
| Agrostis scabra | 0.90 | 0.20 | 0.42 | 0.005 | |
| Equisetum palustre | 0.65 | 0.20 | 0.36 | 0.041 | |
| Vaccinium myrtilloides | 0.65 | 0.20 | 0.36 | 0.021 | |
| Mounded | Carex aquatilis | 0.46 | 0.92 | 0.65 | 0.004 |
| (Ground) | Comarum palustre | 0.47 | 0.88 | 0.64 | 0.002 |
| Salix pedicellaris | 0.43 | 0.88 | 0.61 | 0.006 | |
| Carex magellanica | 0.65 | 0.52 | 0.58 | 0.003 | |
| Carex tenuiflora | 0.41 | 0.56 | 0.48 | 0.049 | |
| Carex diandra | 0.59 | 0.32 | 0.43 | 0.048 | |
| Galium trifidum | 0.65 | 0.24 | 0.40 | 0.045 | |
| Rubus pubescens | 0.78 | 0.20 | 0.40 | 0.001 | |
| Carex utriculata | 0.9 | 0.12 | 0.33 | 0.034 | |
| Ground-dwelling spiders | |||||
| Reference | Diplocentria bidentata | 0.70 | 0.86 | 0.77 | 0.001 |
| Pardosa hyperborea | 0.63 | 0.92 | 0.76 | 0.001 | |
| Piratula insularis | 0.48 | 1.00 | 0.70 | 0.001 | |
| Walckenaeria tricornis | 0.55 | 0.62 | 0.58 | 0.032 | |
| Ozyptila sincera canadensis | 0.66 | 0.50 | 0.57 | 0.004 | |
| Cybaeopsis euopla | 0.56 | 0.56 | 0.56 | 0.016 | |
| Sciastes truncatus | 0.75 | 0.42 | 0.56 | 0.005 | |
| Theonoe stridula | 0.56 | 0.56 | 0.56 | 0.019 | |
| Agyneta olivacea | 0.58 | 0.54 | 0.56 | 0.015 | |
| Diplocentria rectangulata | 0.74 | 0.40 | 0.54 | 0.008 | |
| Vermontia thoracica | 0.82 | 0.28 | 0.48 | 0.012 | |
| Carorita limnaea | 0.64 | 0.32 | 0.45 | 0.045 | |
| Walckenaeria atrotibialis | 0.70 | 0.26 | 0.43 | 0.045 | |
| Untreated | Oedothorax trilobatus | 0.75 | 0.60 | 0.67 | 0.001 |
| Xysticus britcheri | 0.76 | 0.56 | 0.65 | 0.001 | |
| Ceratinella ornatula alaskana | 0.75 | 0.44 | 0.58 | 0.002 | |
| Neon nelli | 0.61 | 0.40 | 0.50 | 0.004 | |
| Cheniseo sphagnicultor | 0.76 | 0.32 | 0.49 | 0.003 | |
| Scotinella pugnata | 0.75 | 0.28 | 0.46 | 0.046 | |
| Mounded | Pardosa fuscula | 0.78 | 0.68 | 0.73 | 0.001 |
| Pardosa xerampelina | 0.95 | 0.44 | 0.65 | 0.001 | |
| Arctosa raptor | 0.57 | 0.64 | 0.60 | 0.011 | |
| Dolomedes striatus | 0.51 | 0.56 | 0.53 | 0.031 | |
| Piratula cantralli | 0.60 | 0.44 | 0.52 | 0.014 | |
| Cicurina arcuata | 0.71 | 0.24 | 0.41 | 0.014 | |
| Aphileta misera | 0.82 | 0.20 | 0.41 | 0.013 | |
| Rove beetles | |||||
| Reference | Reichenbachia spp. | 0.54 | 0.84 | 0.67 | 0.002 |
| Quedius frigidus | 0.54 | 0.72 | 0.62 | 0.012 | |
| Untreated | Dinothenarus pleuralis | 0.77 | 0.28 | 0.47 | 0.006 |
| Mounded | Philonthus flavibasis | 0.68 | 0.50 | 0.58 | 0.003 |
| Olophrum consimile | 0.75 | 0.21 | 0.40 | 0.017 | |
| Ants | |||||
| Untreated | Myrmica lobifrons | 0.63 | 0.76 | 0.69 | 0.005 |
| Formicoxenus quebecensis | 0.89 | 0.20 | 0.42 | 0.012 | |
| Mounded | Formica neorufibarbis | 0.52 | 0.96 | 0.71 | 0.001 |
| Formica dakotensis | 0.98 | 0.20 | 0.44 | 0.004 | |
| Ground beetles | |||||
| Mounded | Agonum gratiosum | 0.74 | 0.44 | 0.57 | 0.001 |
| Pterostichus adstrictus | 0.94 | 0.24 | 0.48 | 0.001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Echiverri, L.; Pinzon, J.; Dabros, A. Inverted Soil Mounding as a Restoration Approach of Seismic Lines in Boreal Peatlands: Implications on Plant and Arthropod Abundance and Diversity. Forests 2023, 14, 2123. https://doi.org/10.3390/f14112123
Echiverri L, Pinzon J, Dabros A. Inverted Soil Mounding as a Restoration Approach of Seismic Lines in Boreal Peatlands: Implications on Plant and Arthropod Abundance and Diversity. Forests. 2023; 14(11):2123. https://doi.org/10.3390/f14112123
Chicago/Turabian StyleEchiverri, Laureen, Jaime Pinzon, and Anna Dabros. 2023. "Inverted Soil Mounding as a Restoration Approach of Seismic Lines in Boreal Peatlands: Implications on Plant and Arthropod Abundance and Diversity" Forests 14, no. 11: 2123. https://doi.org/10.3390/f14112123