
Long Term Seasonal Variability on Litterfall in Tropical Dry Forests, Western Thailand

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Litterfall Collection
2.3. Climate Data
2.4. Data Analysis
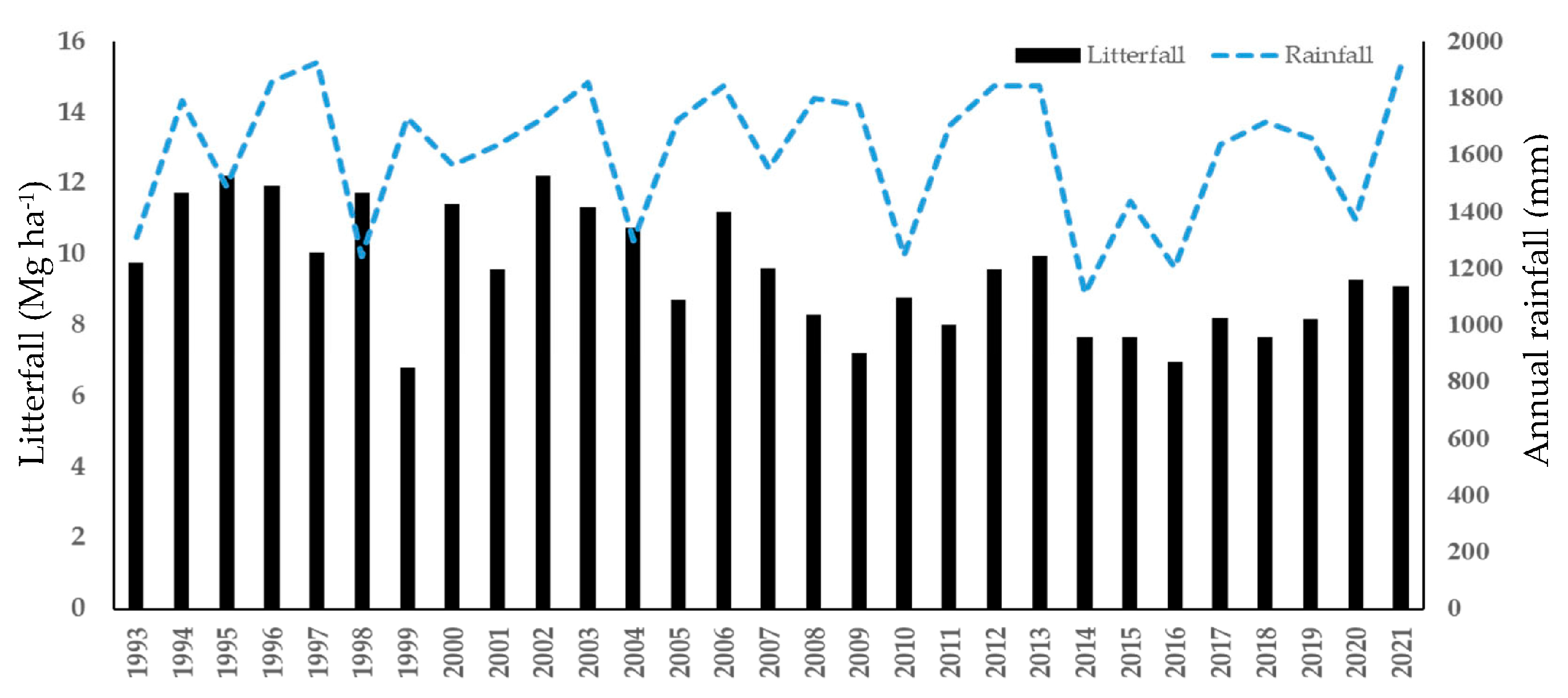
3. Results
3.1. Climate Regime
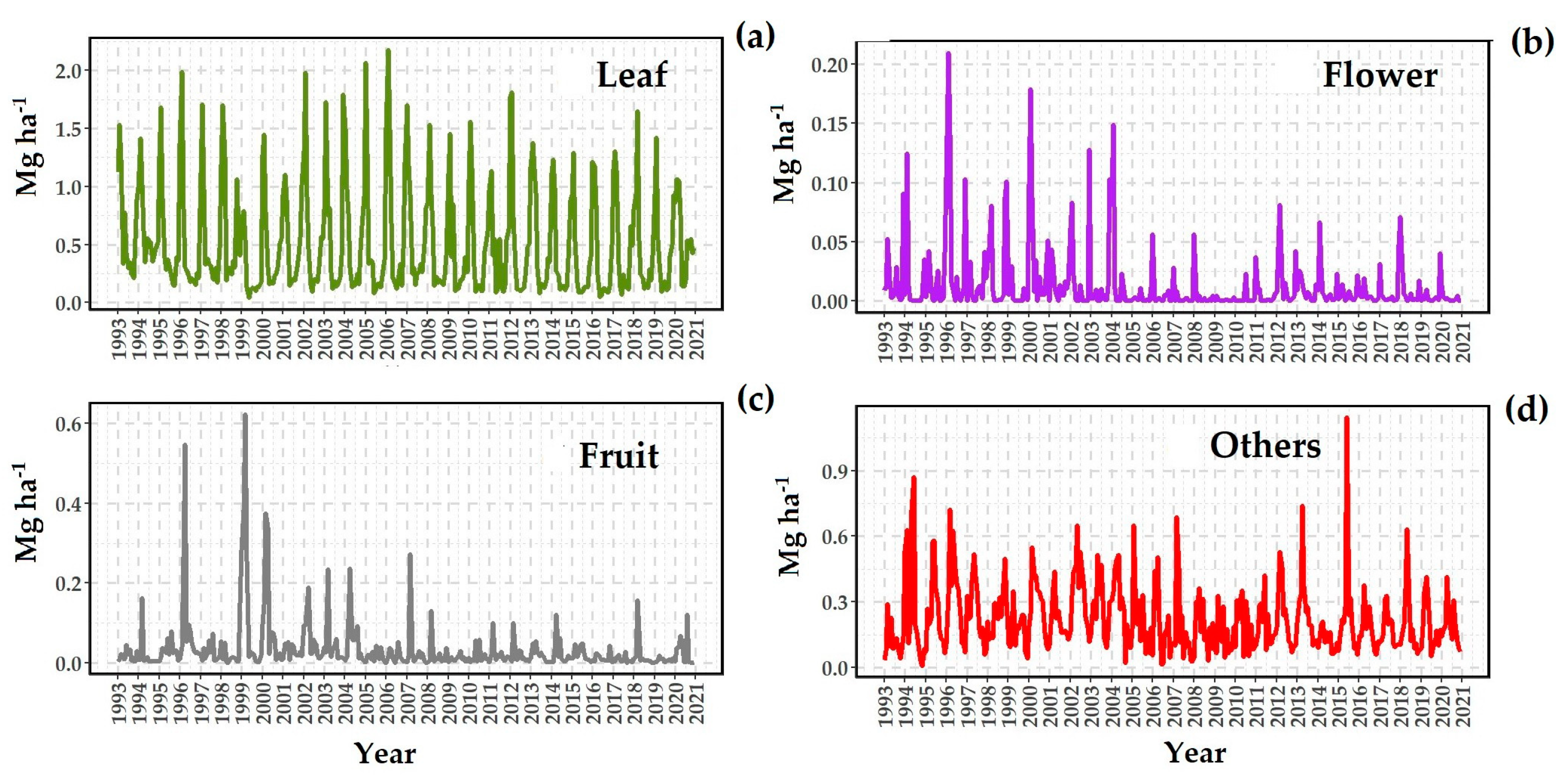
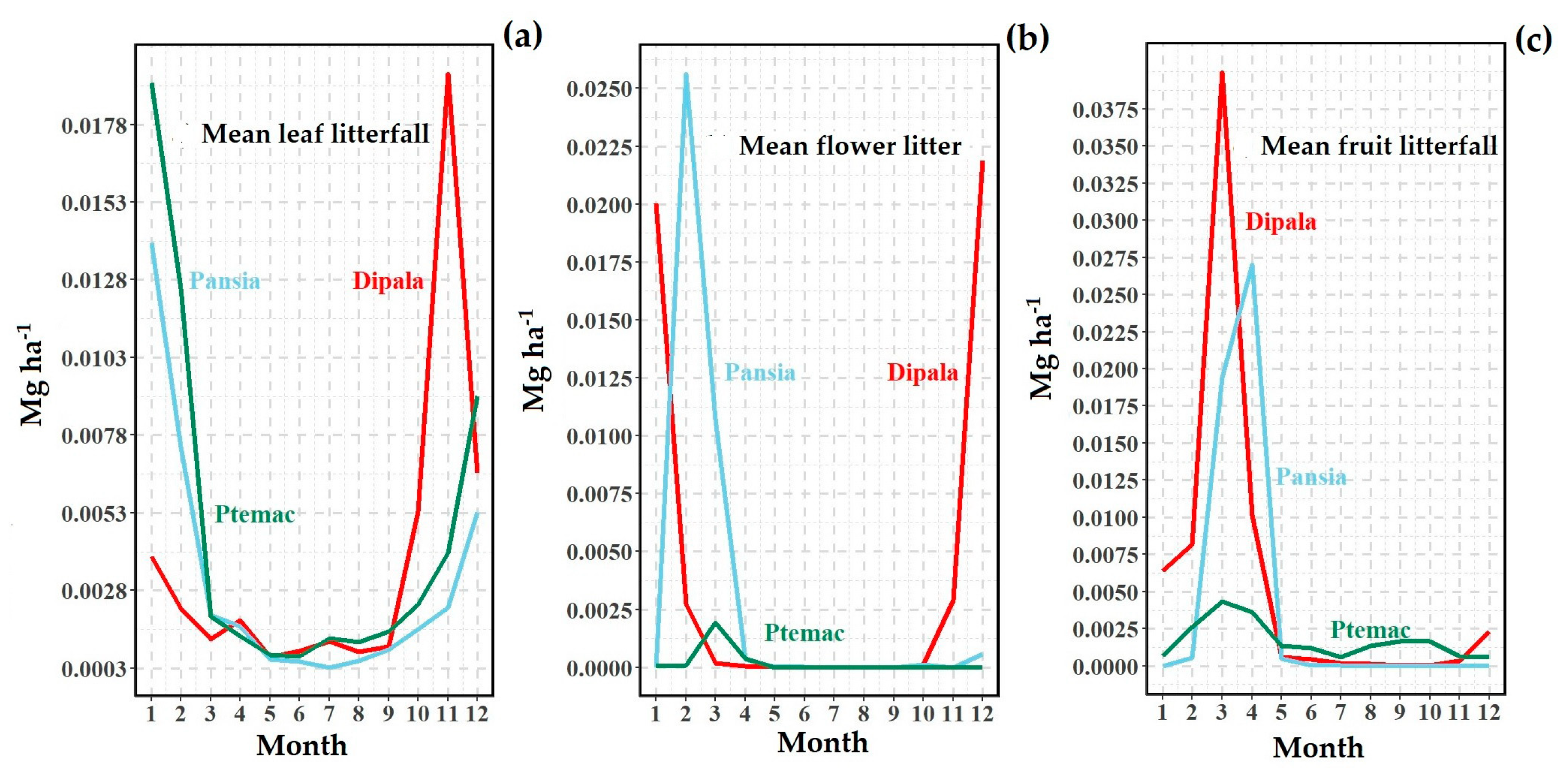
3.2. Litter Production
3.3. Relationship between Litterfall Seasonality and Climate Change
4. Discussion
4.1. Annual Litterfall Production and Seasonality
4.2. Relationship between Litterfall Seasonality and Climate Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Attiwill, P.M.; Adams, M.A. Nutrient cycling in forests. New Phytol. 1993, 124, 561–582. [Google Scholar] [CrossRef]
- Bigelow, S.W.; Canham, C.D. Litterfall as a niche construction process in a northern hardwood forest. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Sayer, E.J.; Joseph Wright, S.; Tanner, E.V.J.; Yavitt, J.B.; Harms, K.E.; Powers, J.S.; Kaspari, M.; Garcia, M.N.; Turner, B.L. Variable responses of lowland tropical forest nutrient status to fertilization and litter manipulation. Ecosystems 2012, 15, 387–400. [Google Scholar] [CrossRef]
- Chave, J.; Navarrete, D.; Almeida, S.; Álvarez, E.; Aragão, L.E.O.C.; Bonal, D.; Châtelet, P.; Silva-Espejo, J.E.; Goret, J.Y.; von Hildebrand, P. Regional and seasonal patterns of litterfall in tropical South America. Biogeosciences 2010, 7, 43–55. [Google Scholar] [CrossRef]
- Kho, L.K.; Malhi, Y.; Tan, S.K.S. Annual budget and seasonal variation of aboveground and belowground net primary productivity in a lowland dipterocarp forest in Borneo. J. Geophys. Res. Biogeosci. 2013, 118, 1282–1296. [Google Scholar] [CrossRef]
- Sayer, E.J.; Powers, J.S.; Tanner, E.V.J. Increased litterfall in tropical forests boosts the transfer of soil CO2 to the atmosphere. PLoS ONE 2007, 2, e1299. [Google Scholar] [CrossRef]
- Marod, D.; Andriyas, T.; Leksungnoen, N.; Kjelgren, R.; Thinkamphaeng, S.; Chansri, P.; Asanok, L.; Hermhuk, S.; Kachina, P.; Thongsawi, J.; et al. Potential variables forcing litterfall in a lower montane evergreen forest using Granger and superposed epoch analyses. Ecosphere 2023, 14, e4572. [Google Scholar] [CrossRef]
- Barlow, J.; Gardner, T.A.; Ferreira, L.V.; Peres, C.A. Litter fall and decomposition in primary, secondary and plantation forests in the Brazilian Amazon. For. Ecol. Manag. 2007, 247, 91–97. [Google Scholar] [CrossRef]
- Lopes, M.C.A.; Araújo, V.F.P.; Vasconcellos, A. The effects of rainfall and vegetation on litterfall production in the semiarid region of northeastern Brazil. Braz. J. Biol. 2015, 75, 703–708. [Google Scholar] [CrossRef]
- Campanella, M.V.; Bertiller, M.B. Leaf litterfall patterns of perennial plant species in the arid Patagonian Monte, Argentina. Plant Ecol. 2010, 210, 43–52. [Google Scholar] [CrossRef]
- Saarsalmi, A.; Starr, M.; Hokkanen, T.; Ukonmaanaho, L.; Kukkola, M.; Nöjd, P.; Sievänen, R. Predicting annual canopy litterfall production for Norway spruce (Picea abies (L.) Karst.) stands. For. Ecol. Manag. 2007, 242, 578–586. [Google Scholar] [CrossRef]
- Zalamea, M.; González, G. Leaffall phenology in a subtropical wet forest in Puerto Rico: From species to community patterns. Biotropica 2008, 40, 295–304. [Google Scholar] [CrossRef]
- Wright, S.J.; van Schaik, C.P. Light and the phenology of tropical trees. Am. Nat. 1994, 143, 192–199. [Google Scholar] [CrossRef]
- Méndez-Alonzo, R.; Pineda-García, F.; Paz, H.; Rosell, J.A.; Olson, M.E. Leaf phenology is associated with soil water availability and xylem traits in a tropical dry forest. Trees 2013, 27, 745–754. [Google Scholar] [CrossRef]
- Smith, V.C.; Ennos, A.R. The effects of air flow and stem flexure on the mechanical and hydraulic properties of the stems of sunflowers Helianthus annuus L. J. Exp. Bot. 2003, 54, 845–849. [Google Scholar] [CrossRef]
- Aryal, D.R.; De Jong, B.H.J.; Ochoa-Gaona, S.; Mendoza-Vega, J.; Esparza-Olguin, L. Successional and seasonal variation in litterfall and associated nutrient transfer in semi-evergreen tropical forests of SE Mexico. Nutr. Cycl. Agroecosystems 2015, 103, 45–60. [Google Scholar] [CrossRef]
- Harrison, R.D. Adaptive significance of phenological variation among monoecious hemi-epiphytic figs in Borneo. Symbiosis 2008, 45, 83–90. [Google Scholar]
- Reich, P.B. Phenology of tropical forests: Patterns, causes, and consequences. Canad. J. Bot. 1995, 73, 164–174. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, W.; Dong, W.; Liu, S. Seasonal patterns of litterfall in forest ecosystem worldwide. Ecol. Complex. 2014, 20, 240–247. [Google Scholar] [CrossRef]
- Sakai, S.; Harrison, R.D.; Momose, K.; Kuraji, K.; Nagamasu, H.; Yasunari, T.; Chong, L.; Nakashizuka, T. Irregular droughts trigger mass flowering in aseasonal tropical forests in Asia. Am. J. Bot. 2006, 93, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Fox, J.E.D.; Xu, Z. Litterfall and nutrient dynamics in a montane moist evergreen broad-leaved forest in Ailao Mountains, SW China. Plant Ecol. 2002, 164, 157–170. [Google Scholar] [CrossRef]
- Miles, L.; Newton, A.C.; DeFries, R.S.; Ravilious, C.; May, I.; Blyth, S.; Kapos, V.; Gordon, J.E. A global overview of the conservation status of tropical dry forests. J. Biogeogr. 2006, 33, 491–505. [Google Scholar] [CrossRef]
- Murphy, P.G.; Lugo, A.E. Dry forests of Central America and the Caribbean. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; Volume 85, pp. 9–34. [Google Scholar]
- Sánchez-Azofeifa, G.A.; Kalacska, M.; Quesada, M.; Calvo-Alvarado, J.C.; Nassar, J.M.; Rodríguez, J.P. Need for Integrated Research for a Sustainable Future in Tropical Dry Forests. Conserv. Biol. 2005, 19, 285–286. [Google Scholar] [CrossRef]
- Morellato, L.P.C.; Talora, D.C.; Takahasi, A.; Bencke, C.C.; Romera, E.C.; Zipparro, V.B. Phenology of Atlantic rain forest trees: A comparative study. Biotropica 2000, 32, 811–823. [Google Scholar] [CrossRef]
- Lawrence, D. Regional-Scale Variation in Litter Production and Seasonality in Tropical Dry Forests of Southern Mexico. Biotropica 2005, 37, 561–570. [Google Scholar] [CrossRef]
- Eamus, D.; Prior, L. Ecophysiology of trees of seasonally dry tropics: Comparisons among phenologies. Adv. Ecol. Res. 2001, 32, 113–197. [Google Scholar]
- Mooney, H.A.; Bullock, S.H.; Medina, E. Introduction. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, UK, 1995; Volume 85, pp. 1–8. [Google Scholar]
- Pezzini, F.F.; Ranieri, B.D.; Brandão, D.O.; Fernandes, G.W.; Quesada, M.; Espírito-Santo, M.M.; Jacobi, C.M. Changes in tree phenology along natural regeneration in a seasonally dry tropical forest. Plant Biosyst. 2014, 148, 965–974. [Google Scholar] [CrossRef]
- Meentemeyer, V.; Box, E.O.; Thompson, R. World patterns and amounts of terrestrial plant litter production. BioScience 1982, 32, 125–128. [Google Scholar] [CrossRef]
- Detto, M.; Wright, S.J.; Calderón, O.; Muller-Landau, H.C. Resource acquisition and reproductive strategies of tropical forest in response to the El Niño–Southern Oscillation. Nat. Commun. 2018, 9, 913. [Google Scholar] [CrossRef]
- Nakagawa, M.; Ushio, M.; Kume, T.; Nakashizuka, T. Seasonal and long-term patterns in litterfall in a Bornean tropical rainforest. Ecol. Res. 2019, 34, 31–39. [Google Scholar] [CrossRef]
- Wagner, F.H.; Hérault, B.; Bonal, D.; Stahl, C.; Anderson, L.O.; Baker, T.R.; Becker, G.S.; Beeckman, H.; Boanerges Souza, D.; Botosso, P.C. Climate seasonality limits leaf carbon assimilation and wood productivity in tropical forests. Biogeosciences 2016, 13, 2537–2562. [Google Scholar] [CrossRef]
- Thuille, A.; Schulze, E.D. Carbon dynamics in successional and afforested spruce stands in Thuringia and the Alps. Glob. Chang. Biol. 2006, 12, 325–342. [Google Scholar] [CrossRef]
- Scheer, M.B.; Gatti, G.; Wisniewski, C. Nutrient fluxes in litterfall of a secondary successional alluvial rain forest in Southern Brazil. Rev. Biol. Trop. 2011, 59, 1869–1882. [Google Scholar] [CrossRef]
- Parsons, S.A.; Congdon, R.A.; Shoo, L.P.; Valdez-Ramirez, V.; Williams, S.E. Spatial variability in litterfall, litter standing crop and litter quality in a tropical rain forest region. Biotropica 2014, 46, 378–386. [Google Scholar] [CrossRef]
- Sundarapandian, S.M.; Swamy, P.S. Litter production and leaf-litter decomposition of selected tree species in tropical forests at Kodayar in the Western Ghats, India. For. Ecol. Manag. 1999, 123, 231–244. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Jassal, R.S. Long term aboveground litterfall production in boreal jack pine (Pinus banksiana) and black spruce (Picea mariana) stands along the Boreal Forest Transect Case Study in western central Canada. Écoscience 2014, 21, 301–314. [Google Scholar] [CrossRef]
- Takahashi, M.; Hirai, K.; Limtong, P.; Leaungvutivirog, C.; Panuthai, S.; Suksawang, S.; Anusontpornperm, S.; Marod, D. Topographic variation in heterotrophic and autotrophic soil respiration in a tropical seasonal forest in Thailand. J. Soil Sci. Plant Nutr. 2011, 57, 452–465. [Google Scholar] [CrossRef]
- Marod, D.; Kutintara, U.; Yarwudhi, C.; Tanaka, H.; Nakashisuka, T. Structural dynamics of a natural mixed deciduous forest in western Thailand. J. Veg. Sci. 1999, 10, 777–786. [Google Scholar] [CrossRef]
- Marod, D.; Kutintara, U.; Tanaka, H.; Nakashizuka, T. The effects of drought and fire on seed and seedling dynamics in a tropical seasonal forest in Thailand. Plant Ecol. 2002, 161, 41–57. [Google Scholar] [CrossRef]
- Proctor, J.; Anderson, J.M.; Chai, P.; Vallack, H.W. Ecological studies in four contrasting lowland rain forests in Gunung Mulu National Park, Sarawak: I. Forest environment, structure and floristics. J. Ecol. 1983, 71, 237–260. [Google Scholar] [CrossRef]
- Metcalfe, D.; Meir, P.; Aragao, L.E.O.C.; da Costa, A.; Almeida, S.; Braga, A.; Gonçalves, P.; Athaydes, J.; Malhi, Y.; Williams, M. Sample sizes for estimating key ecosystem characteristics in a tropical terra firme rainforest. For. Ecol. Manag. 2008, 255, 558–566. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: Berlin/Heidelberg, Germany, 2002; pp. 149–205. [Google Scholar]
- Guisan, A.; Edwards Jr, T.C.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: http://www.R-project.org/ (accessed on 7 March 2022).
- Wood, S.; Wood, M.S. Package ‘mgcv’. In R Package Version 1.29; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Wang, C.G.; Zheng, X.B.; Wang, A.Z.; Dai, G.H.; Zhu, B.K.; Zhao, Y.M.; Dong, S.J.; Zu, W.Z.; Wang, W.; Zheng, Y.G. Temperature and Precipitation Diversely Control Seasonal and Annual Dynamics of Litterfall in a Temperate Mixed Mature Forest, Revealed by Long-Term Data Analysis. J. Geophys. Res. Biogeosci. 2021, 126, e2020JG006204. [Google Scholar] [CrossRef]
- Bray, J.R.; Gorham, E. Litter production in forests of the world. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1964; Volume 2, pp. 101–157. [Google Scholar]
- Ahirwal, J.; Saha, P.; Nath, A.; Nath, A.J.; Deb, S.; Sahoo, U.K. Forests litter dynamics and environmental patterns in the Indian Himalayan region. For. Ecol. Manag. 2021, 499, 119612. [Google Scholar] [CrossRef]
- Hanpattanakit, P.; Chidthaisong, A. Litter production and decomposition in dry dipterocarp forest and their responses to climatic factors. GMSARN Int. J. 2012, 6, 169–174. [Google Scholar]
- Martínez-Yrízar, A.; Sarukhán, J. Litterfall patterns in a tropical deciduous forest in Mexico over a five-year period. J. Trop. Ecol. 1990, 6, 433–444. [Google Scholar] [CrossRef]
- Whigham, D.F.; Zugasty Towle, P.; Cabrera Cano, E.F.; O’Neill, J.P.; Ley, E. Effect of annual variation in precipitation on growth and litter production in a tropical dry forest in the Yucatan of Mexico. Trop. Ecol. 1990, 31, 23–34. [Google Scholar]
- Köhler, L.; Hölscher, D.; Leuschner, C. High litterfall in old-growth and secondary upper montane forest of Costa Rica. Plant Ecol. 2008, 199, 163–173. [Google Scholar] [CrossRef]
- Swamy, S.L.; Dutt, C.B.S.; Murthy, M.S.R.; Mishra, A.; Bargali, S.S. Floristics and dry matter dynamics of tropical wet evergreen forests of Western Ghats, India. Curr. Sci. 2010, 99, 353–364. [Google Scholar]
- Tang, J.-W.; Cao, M.; Zhang, J.-H.; Li, M.-H. Litterfall production, decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: A 10-year study. Plant Soil 2010, 335, 271–288. [Google Scholar] [CrossRef]
- Park, B.B.; Rahman, A.; Han, S.H.; Youn, W.B.; Hyun, H.J.; Hernandez, J.; An, J.Y. Carbon and nutrient inputs by litterfall in evergreen and deciduous forests in Korea. Forests 2020, 11, 143. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox. Silva Fenn. 2002, 36, 703–743. [Google Scholar] [CrossRef]
- Borchert, R.; Meyer, S.A.; Felger, R.S.; Porter-Bolland, L. Environmental control of flowering periodicity in Costa Rican and Mexican tropical dry forests. Glob. Ecol. Biogeogr. 2004, 13, 409–425. [Google Scholar] [CrossRef]
- Nanda, A.; Suresh, H.S.; Krishnamurthy, Y.L. Phenology of a tropical dry deciduous forest of Bhadra wildlife sanctuary, southern India. Ecol. Process. 2014, 3, 1. [Google Scholar] [CrossRef]
- Yang, X.; Wu, J.; Chen, X.; Ciais, P.; Maignan, F.; Yuan, W.; Piao, S.; Yang, S.; Gong, F.; Su, Y. A comprehensive framework for seasonal controls of leaf abscission and productivity in evergreen broadleaved tropical and subtropical forests. Innovation 2021, 2, 1–7. [Google Scholar] [CrossRef]
- Liu, C.; Westman, C.J.; Berg, B.; Kutsch, W.; Wang, G.Z.; Man, R.; Ilvesniemi, H. Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia. Glob. Ecol. Biogeogr. 2004, 13, 105–114. [Google Scholar] [CrossRef]
- Cleverly, J.; Eamus, D.; Coupe, N.R.; Chen, C.; Maes, W.; Li, L.; Faux, R.; Santini, N.S.; Rumman, R.; Yu, Q. Soil moisture controls on phenology and productivity in a semi-arid critical zone. Sci. Total Environ. 2016, 568, 1227–1237. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; Xie, J.S.; Gao, R.; Li, Z.; Jin, Z. Litter production, seasonal pattern and nutrient return in seven natural forests compared with a plantation in southern China. Forestry 2005, 78, 403–415. [Google Scholar] [CrossRef]
- De Queiroz, M.G.; da Silva, T.G.F.; Zolnier, S.; de Souza, C.A.A.; de Souza, L.S.B.; Neto, A.J.S.; de Araújo, G.G.L.; Ferreira, W.P.M. Seasonal patterns of deposition litterfall in a seasonal dry tropical forest. Agric. For. Meteorol. 2019, 279, 107712. [Google Scholar] [CrossRef]
- Lin, K.-C.; Hamburg, S.P.; Tang, S.-l.; Hsia, Y.-J.; Lin, T.-C. Typhoon effects on litterfall in a subtropical forest. Can. J. For. Res. 2003, 33, 2184–2192. [Google Scholar] [CrossRef]
- An, J.Y.; Han, S.H.; Youn, W.B.; Lee, S.I.; Rahman, A.; Dao, H.T.T.; Seo, J.M.; Aung, A.; Choi, H.-S.; Hyun, H.J. Comparison of litterfall production in three forest types in Jeju Island, South Korea. J. For. Res. 2020, 31, 945–952. [Google Scholar] [CrossRef]
- Song, Y.-J.; Tian, W.-B.; Liu, X.-Y.; Yin, F.; Cheng, J.-Y.; Zhu, D.-N.; Ali, A.; Yan, E.-R. Associations between litterfall dynamics and micro-climate in forests of Putuoshan Island, Zhejiang, China. Chin. J. Plant Ecol. 2016, 40, 1154. [Google Scholar]
- Bou, J.; Caritat, A.; Vilar, L. Litterfall and growth dynamics relationship with the meteorological variability in three forests in the Montseny natural park. Folia For. Pol. Ser. A For. 2015, 57, 145–159. [Google Scholar] [CrossRef]
- Brearley, F.Q.; Proctor, J.; Suriantata; Nagy, L.; Dalrymple, G.; Voysey, B.C. Reproductive phenology over a 10-year period in a lowland evergreen rain forest of central Borneo. J. Ecol. 2007, 95, 828–839. [Google Scholar] [CrossRef]
- Curran, L.M.; Caniago, I.; Paoli, G.D.; Astianti, D.; Kusneti, M.; Leighton, M.; Nirarita, C.E.; Haeruman, H. Impact of El Nino and logging on canopy tree recruitment in Borneo. Science 1999, 286, 2184–2188. [Google Scholar] [CrossRef]
- Wright, S.J.; Carrasco, C.; Calderon, O.; Paton, S. The El Niño Southern Oscillation, variable fruit production, and famine in a tropical forest. Ecology 1999, 80, 1632–1647. [Google Scholar] [CrossRef]
- Ashton, P.S.; Givnish, T.J.; Appanah, S. Staggered flowering in the Dipterocarpaceae: New insights into floral induction and the evolution of mast fruiting in the aseasonal tropics. Am. Nat. 1988, 132, 44–66. [Google Scholar] [CrossRef]
- Van Schaik, C.P.; Terborgh, J.W.; Wright, S.J. The phenology of tropical forests: Adaptive significance and the consequences for primary consumers. Annu. Rev. Ecol. Evol. Syst. 1993, 24, 353–377. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Litter Fraction | Model Structure | Deviance Explained [%] | R2 [%] | Variable Significance |
|---|---|---|---|---|
| Leaf | Leaf.litter~f(Year) + f(Month) +f(Tmax) + f(SM) + f(ET) | 83.6 | 82.1 | Year [p < 0.0001 ***] Month [p < 0.0001 ***] Tmax [p = 0.0293 *] SM [p < 0.0001 ***] ET [p = 0.0013 **] |
| Flower | Flower.litter ~ f(Year) + f(Month) +f(VPD) + f(Light) + f(ET) | 67.2 | 65 | Year [p < 0.0001 ***] Month [p = 0.03486 *] VPD [p = 0.00284 **] Light [p = 0.02393 *] ET [p < 0.0001 ***] |
| Fruit | Fruit.litter ~ f(Year) + f(Month) +f(Light) + f(Tmean) + f(SOI) | 67.2 | 64.9 | Year [p < 0.0001 ***] Month [p < 0.0001 ***] Light [p = 0.001668 **] Tmean [p < 0.0001 ***] SOI [p < 0.0001 ***] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marod, D.; Nakashizuka, T.; Saitoh, T.; Hirai, K.; Thinkampheang, S.; Asanok, L.; Phumphuang, W.; Danrad, N.; Pattanakiat, S. Long Term Seasonal Variability on Litterfall in Tropical Dry Forests, Western Thailand. Forests 2023, 14, 2107. https://doi.org/10.3390/f14102107
Marod D, Nakashizuka T, Saitoh T, Hirai K, Thinkampheang S, Asanok L, Phumphuang W, Danrad N, Pattanakiat S. Long Term Seasonal Variability on Litterfall in Tropical Dry Forests, Western Thailand. Forests. 2023; 14(10):2107. https://doi.org/10.3390/f14102107
Chicago/Turabian StyleMarod, Dokrak, Tohru Nakashizuka, Tomoyuki Saitoh, Keizo Hirai, Sathid Thinkampheang, Lamthai Asanok, Wongsatorn Phumphuang, Noppakun Danrad, and Sura Pattanakiat. 2023. "Long Term Seasonal Variability on Litterfall in Tropical Dry Forests, Western Thailand" Forests 14, no. 10: 2107. https://doi.org/10.3390/f14102107