
Pathways and Characteristics of Lead Uptake and Transportation in Rhus chinensis Mill

Abstract
:1. Introduction
2. Materials and methods
2.1. Plant Materials and Growth Conditions
2.2. Experimental Method
2.2.1. Time Course Experiment
2.2.2. Investigation of the Concentration-Dependent Kinetics of Pb
2.2.3. Effect of Different Culture Medium on Pb Uptake and Transportation
2.2.4. Effect of Metabolic Inhibitors on Pb Uptake and Transportation
2.2.5. Effect of Transpiration Inhibitors on Pb Uptake and Transportation
2.2.6. Effect of Low Temperature on Pb Uptake and Transportation
2.2.7. Effects of Ion Channel Inhibitors and Protein Synthesis Inhibitors on Pb Uptake and Transportation
2.3. Histochemical Staining of Pb in Roots
2.4. Statistical Analysis
3. Results
3.1. Time-Dependent Uptake of Pb in Different Organs
3.2. Concentration-Dependent Kinetics of Lead Uptake
3.3. Effect of Different Culture Medium on Pb Uptake and Transportation
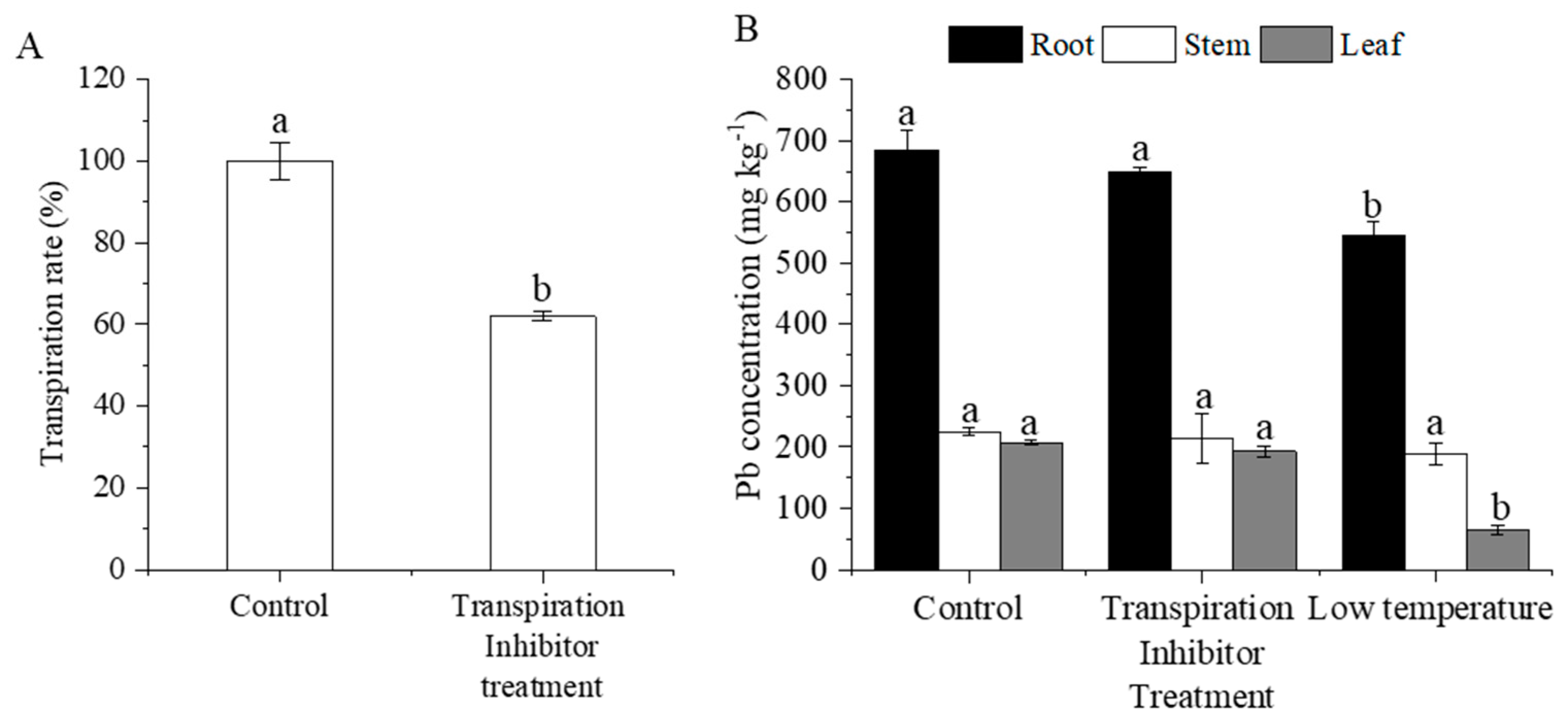
3.4. Effect of Transpiration Inhibitors and Low Temperatures on Pb Uptake and Transportation
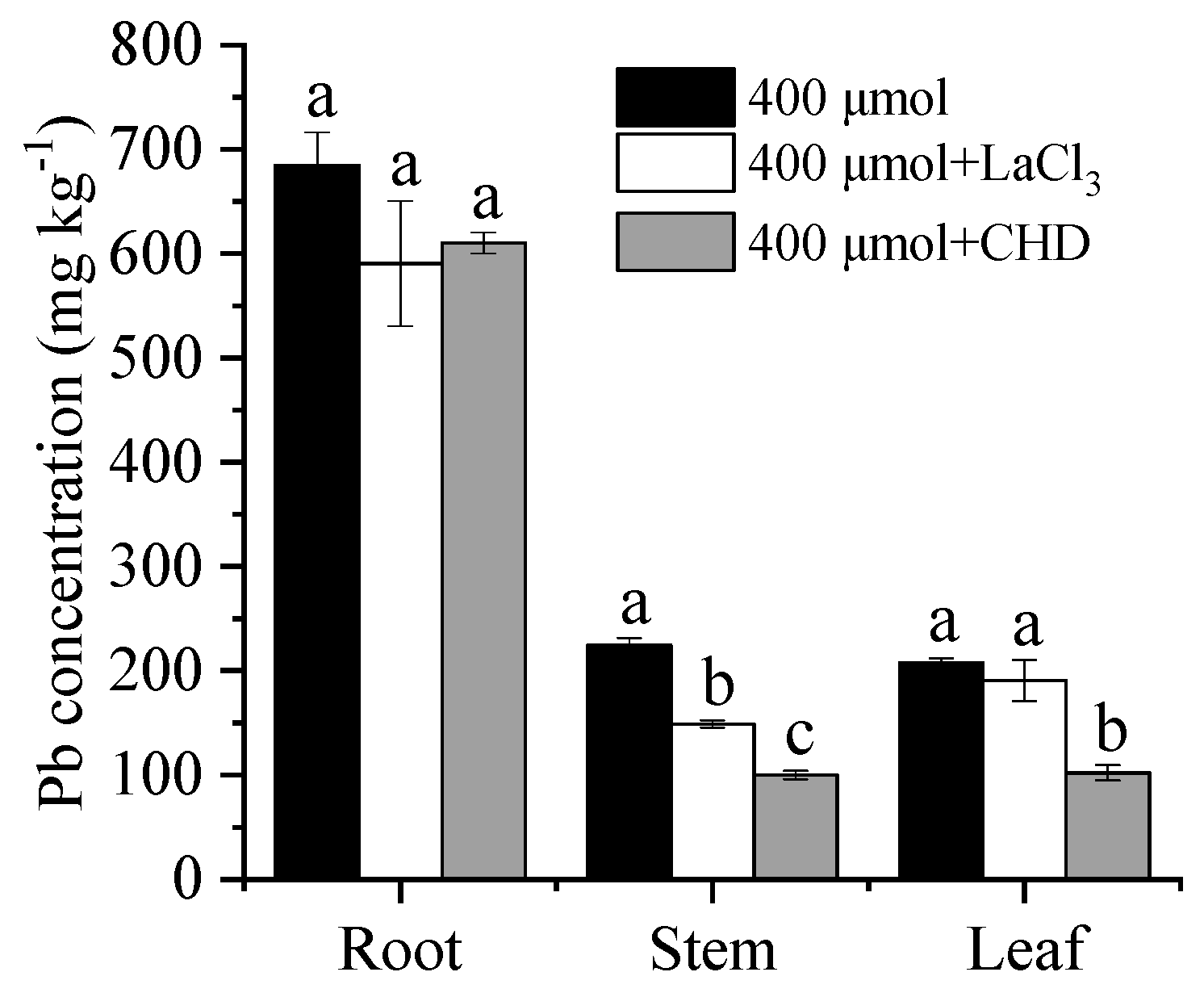
3.5. Effect of Metabolic Inhibitors on Pb Uptake and Transportation
3.6. Effect of Ion Channel Inhibitors and Protein Inhibitors on Lead Uptake
3.7. Pb Localization in Roots
4. Discussion
4.1. Characteristics of Pb Uptake in the Roots of R. chinensis
4.2. Pathways of Pb Uptake and Transportation in R. chinensis
4.3. The Role of Ca Pathway in Pb Uptake and Translocation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef] [PubMed]
- Gul, I.; Manzoor, M.; Kallerhoff, J.; Arshad, M. Enhanced phytoremediation of lead by soil applied organic and inorganic amendments: Pb phytoavailability, accumulation and metal recovery. Chemosphere 2020, 258, 127405. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liao, J.; Chen, Y.; Tian, Y.; Chen, Q.; Gao, S.; Luo, Z.; Yu, X.; Lei, T.; Jiang, M. Efficiency of heterogeneous chelating agents on the phytoremediation potential and growth of Sasa argenteostriata (Regel) EG Camus on Pb-contaminated soil. Ecotoxicol. Environ. Saf. 2022, 238, 113603. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Iqbal, M.; Raza, S.H.; Anwar, S.; Ashraf, M.; Shafiq, F. Tartaric acid soil-amendment increases phytoextraction potential through root to shoot transfer of lead in turnip. Chemosphere 2022, 296, 134055. [Google Scholar] [CrossRef]
- Aghelan, N.; Sobhanardakani, S.; Cheraghi, M.; Lorestani, B.; Merrikhpour, H. Evaluation of some chelating agents on phytoremediation efficiency of Amaranthus caudatus L. and Tagetes patula L. in soils contaminated with lead. J. Environ. Health Sci. Eng. 2021, 19, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Zehra, A.; Sahito, Z.A.; Tong, W.; Tang, L.; Hamid, Y.; Khan, M.B.; Ali, Z.; Naqvi, B.; Yang, X. Assessment of sunflower germplasm for phytoremediation of lead-polluted soil and production of seed oil and seed meal for human and animal consumption. J. Environ. Sci. 2020, 87, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xiao, J.; Salam, M.M.A.; Chen, G. Evaluation of dendroremediation potential of ten Quercus spp. for heavy metals contaminated soil: A three-year field trial. Sci. Total Environ. 2022, 851, 158232. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Shi, X.; Salam, M.M.A.; Chen, G. Integrated study on subcellular localization and chemical speciation of Pb reveals root strategies for Pb sequestration and detoxification in Salix integra. Plant Soil 2021, 467, 197–211. [Google Scholar] [CrossRef]
- Arshad, M.; Naqvi, N.; Gul, I.; Yaqoob, K.; Bilal, M.; Kallerhoff, J. Lead phytoextraction by Pelargonium hortorum: Comparative assessment of EDTA and DIPA for Pb mobility and toxicity. Sci. Total Environ. 2020, 748, 141496. [Google Scholar] [CrossRef]
- Gul, I.; Manzoor, M.; Hashim, N.; Shah, G.M.; Waani, S.P.T.; Shahid, M.; Antoniadis, V.; Rinklebe, J.; Arshad, M. Challenges in microbially and chelate-assisted phytoextraction of cadmium and lead–A review. Environ. Pollut. 2021, 287, 117667. [Google Scholar] [CrossRef]
- Liang, Y.; Xiao, X.; Guo, Z.; Peng, C.; Zeng, P.; Wang, X. Co-application of indole-3-acetic acid/gibberellin and oxalic acid for phytoextraction of cadmium and lead with Sedum alfredii Hance from contaminated soil. Chemosphere 2021, 285, 131420. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Han, H.; Tigabu, M.; Li, Q.; Li, Z.; Zhu, C.; Huang, S.; Cai, L.; Liu, A. Lead contamination alters enzyme activities and microbial composition in the rhizosphere soil of the hyperaccumulator Pogonatherum crinitum. Ecotoxicol. Environ. Saf. 2021, 207, 111308. [Google Scholar] [CrossRef] [PubMed]
- Krzesłowska, M.; Lenartowska, M.; Samardakiewicz, S.; Bilski, H.; Woźny, A. Lead deposited in the cell wall of Funaria hygrometrica protonemata is not stable—A remobilization can occur. Environ. Pollut. 2010, 158, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Gong, X.F.; Tang, Y.P.; Chen, C.L. Lead sequestration in iron plaques developed on Phalaris arundinacea Linn. and Carex cinerascens Kukenth. from Poyang Lake (China). Aquat. Bot. 2015, 122, 54–59. [Google Scholar] [CrossRef]
- Salazar, M.J.; Rodriguez, J.H.; Cid, C.V.; Pignata, M.L. Auxin effects on Pb phytoextraction from polluted soils by Tegetes minuta L. and Bidens pilosa L.: Extractive power of their root exudates. J. Hazard. Mater. 2016, 311, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Tian, S.; Yang, X.; Wang, X.; Brown, P.; Li, T.; He, Z. Enhanced root-to-shoot translocation of cadmium in the hyperaccumulating ecotype of Sedum alfredii. J. Exp. Bot. 2008, 59, 3203–3213. [Google Scholar] [CrossRef] [Green Version]
- Tao, Q.; Jupa, R.; Luo, J.; Lux, A.; Kováč, J.; Wen, Y.; Zhou, Y.; Jan, J.; Liang, Y.; Li, T. The apoplasmic pathway via the root apex and lateral roots contributes to Cd hyperaccumulation in the hyperaccumulator Sedum alfredii. J. Exp. Bot. 2016, 68, 739–751. [Google Scholar]
- Pasricha, S.; Mathur, V.; Garg, A.; Lenka, S.; Verma, K.; Agarwal, S. Molecular mechanisms underlying heavy metal uptake, translocation and tolerance in hyperaccumulators-an analysis: Heavy metal tolerance in hyperaccumulators. Environ. Chall. 2021, 4, 100197. [Google Scholar] [CrossRef]
- Vaculík, M.; Konlechner, C.; Langer, I.; Adlassnig, W.; Puschenreiter, M.; Lux, A.; Hauser, M.-T. Root anatomy and element distribution vary between two Salix caprea isolates with different Cd accumulation capacities. Environ. Pollut. 2012, 163, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Lyubenova, L.; Pongrac, P.; Vogel-Mikuš, K.; Mezek, G.K.; Vavpetič, P.; Grlj, N.; Kump, P.; Nečemer, M.; Regvar, M.; Pelicon, P.; et al. Localization and quantification of Pb and nutrients in Typha latifolia by micro-PIXE. Metallomics 2012, 4, 333–341. [Google Scholar] [CrossRef]
- Sahi, S.V.; Bryant, N.L.; Sharma, N.C.; Singh, S.R. Characterization of a lead hyperaccumulator shrub, Sesbania drummondii. Environ. Sci. Technol. 2002, 36, 4676–4680. [Google Scholar] [CrossRef] [PubMed]
- Samardakiewicz, S.; Woźny, A. The distribution of lead in duckweed (Lemna minor L.) root tip. Plant Soil. 2000, 226, 107–111. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Yang, L.; Liang, J.; Song, Y.; Jiang, S.; Qin, L. Transport pathways of cadmium (Cd) and its regulatory mechanisms in plant. Acta Ecol. Sin. 2015, 35, 7921–7929. [Google Scholar]
- Kumar, B.; Smita, K.; Cumbal Flores, L. Plant mediated detoxification of mercury and lead. Arab. Chem. 2017, 10, S2335–S2342. [Google Scholar]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar]
- Xu, J. Mechanisms of lead uptake/accumulation and tolerance in tea plant (Camellia sinensis L.). Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2011. [Google Scholar]
- Zhou, C.; Huang, M.; Ren, H.; Yu, J.; Wu, J.; Ma, X. Bioaccumulation and detoxification mechanisms for lead uptake identified in Rhus chinensis Mill. seedlings. Ecotoxicol. Environ. Saf. 2017, 142, 59–68. [Google Scholar] [CrossRef]
- Shi, X.; Wang, S.; Wang, D.; Sun, H.; Chen, Y.; Liu, J.; Jiang, Z. Woody species Rhus chinensis Mill. seedlings tolerance to Pb: Physiological and biochemical response. J. Environ. Sci. 2019, 78, 63–73. [Google Scholar] [CrossRef]
- Shi, X.; Chen, Y.; Wang, S.; Li, J. Pb, Zn accumulation and nutrient uptake of 15 plant species grown in abandoned mine tailings. Environ. Sci. (In Chinese) 2012, 33, 2021–2027. [Google Scholar]
- Lu, L.; Tian, S.; Yang, X.; Li, T.; He, Z. Cadmium uptake and xylem loading are active processes in the hyperaccumulator Sedum alfredii. J. Plant Physiol. 2009, 166, 579–587. [Google Scholar] [CrossRef]
- Wang, H.; Shan, X.; Liu, T.; Xie, Y.; Wen, B.; Zhang, S.; Han, F.; van Genuchten, M.T. Organic acids enhance the uptake of lead by wheat roots. Planta 2007, 225, 1483–1494. [Google Scholar] [CrossRef]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Huang, M.; Yin, D.; Wu, P.; Cai, L.; Zhou, C. Uptake kinetic characteristics and subcellular distribution of Pb2+ and Cd2+ in Neyraudia reynaudiana. J. Agro-Environ. Sci (In Chinese) 2016, 35, 1451–1457. [Google Scholar]
- Huang, X.; Duan, S.; Wu, Q.; Yu, M.; Shabala, S. Reducing cadmium accumulation in plants: Structure-function relations and tissue-specific operation of transporters in the spotlight. Plants 2020, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Meyers, D.E.R.; Auchterlonie, G.J.; Webb, R.I.; Wood, B. Uptake and localisation of lead in the root system of Brassica juncea. Environ. Pollut. 2008, 153, 323–332. [Google Scholar] [CrossRef]
- Tung, G.; Temple, P.J. Uptake and localization of lead in corn (Zea mays L.) seedlings, a study by histochemical and electron microscopy. Sci. Total Environ. 1996, 188, 71–85. [Google Scholar] [CrossRef]
- Liu, D. Study on Absorption Characteristics of Lead by Sedum alfredii Hance, a Lead-Rich Plant, and Its Chelation-Induced Remediation Effect. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2007. [Google Scholar]
- Liu, Y. Researches on Adsorption of Cadmium by Willow Root Cell Wall and Kinetics of Cadmium Uptake by Willow Root. M.D. Thesis, Zhejiang University, Hangzhou, China, 2011. [Google Scholar]
- Liu, X.; Peng, K.; Wang, A.; Lian, C.; Shen, Z. Cadmium accumulation and distribution in populations of Phytolacca americana L. and the role of transpiration. Chemosphere 2010, 78, 1136–1141. [Google Scholar] [CrossRef]
- Kim, Y.-Y.; Yang, Y.-Y.; Lee, Y. Pb and Cd uptake in rice roots. Physiol. Plant. 2002, 116, 368–372. [Google Scholar] [CrossRef]
- Tian, S.; Xie, R.; Wang, H.; Hu, Y.; Ge, J.; Liao, X.; Gao, X.; Brown, P.; Lin, X.; Lu, L. Calcium deficiency triggers phloem remobilization of cadmium in a hyperaccumulating species. Plant Physiol. 2016, 172, 2300–2313. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Tian, S.; Zhang, M.; Zhang, J.; Yang, X.; Jiang, H. The role of Ca pathway in Cd uptake and translocation by the hyperaccumulator Sedum alfredii. J. Hazard. Mater. 2010, 183, 22–28. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vmax (nmol g−1 h−1) | Km | Vmax/Km | a | R2 | |
|---|---|---|---|---|---|
| Root | 14.83 | 19.44 | 0.763 | 0.0733 | 0.998 |
| Stem | 7.1 | 82.23 | 0.086 | 0.0206 | 0.882 |
| Leaf | 3.08 | 36.06 | 0.085 | 0.0242 | 0.946 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.; Wang, S.; Wang, Y.; Lu, M.; Shi, X. Pathways and Characteristics of Lead Uptake and Transportation in Rhus chinensis Mill. Forests 2023, 14, 90. https://doi.org/10.3390/f14010090
He W, Wang S, Wang Y, Lu M, Shi X. Pathways and Characteristics of Lead Uptake and Transportation in Rhus chinensis Mill. Forests. 2023; 14(1):90. https://doi.org/10.3390/f14010090
Chicago/Turabian StyleHe, Wenxiang, Shufeng Wang, Yangdong Wang, Mengzhu Lu, and Xiang Shi. 2023. "Pathways and Characteristics of Lead Uptake and Transportation in Rhus chinensis Mill" Forests 14, no. 1: 90. https://doi.org/10.3390/f14010090