
Effects of Nitrogen Form on Root Activity and Nitrogen Uptake Kinetics in Camellia oleifera Seedlings

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Experimental Field
2.2. Experimental Materials
2.3. Experimental Design
2.4. Experimental Methods
2.5. Experimental Materials
3. Results
3.1. Effects of Nitrogen Form on the Root Activity of C. oleifera Seedlings
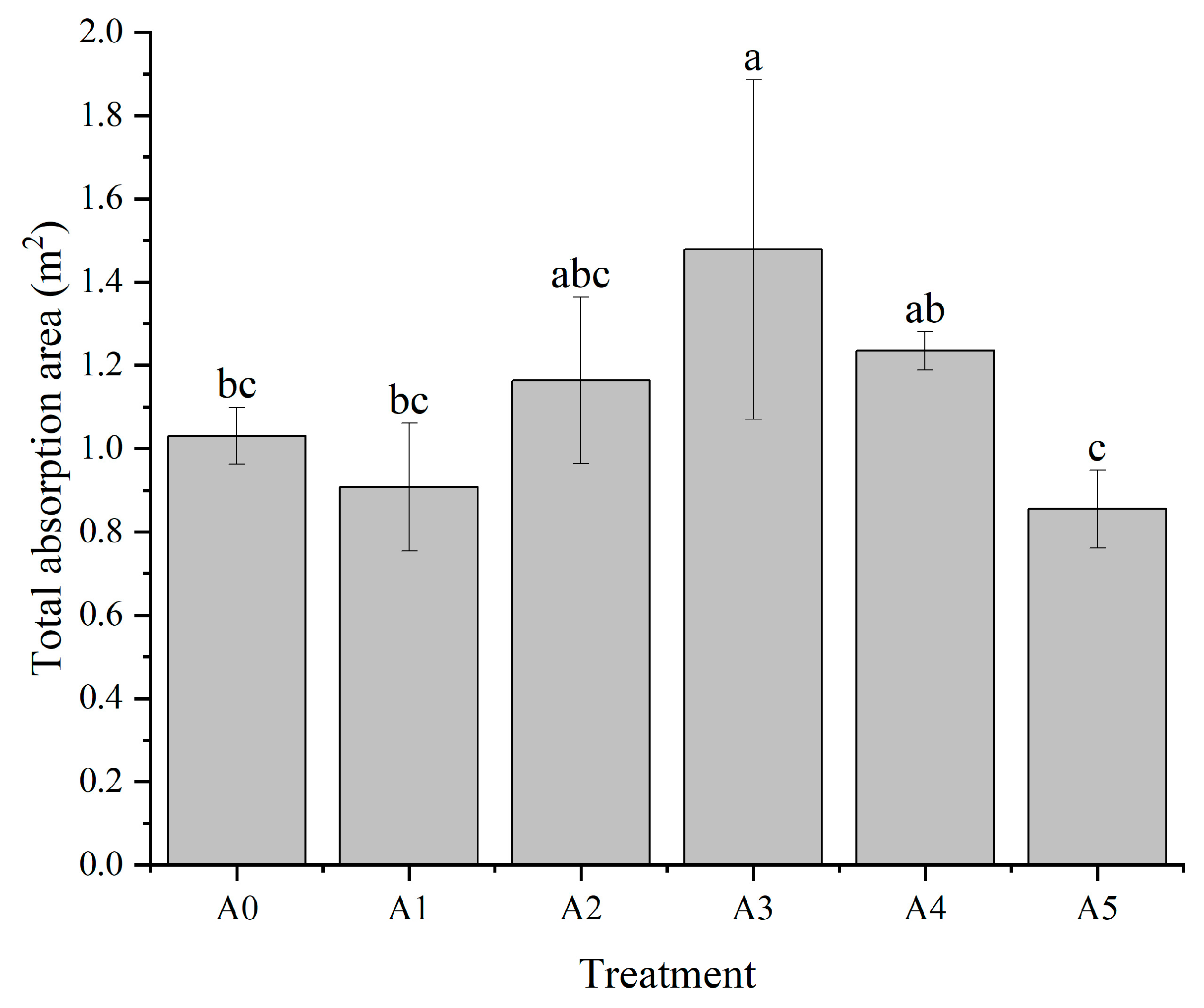
3.1.1. Effect of Nitrogen Form on the Total Absorption Area of C. oleifera Seedling Roots
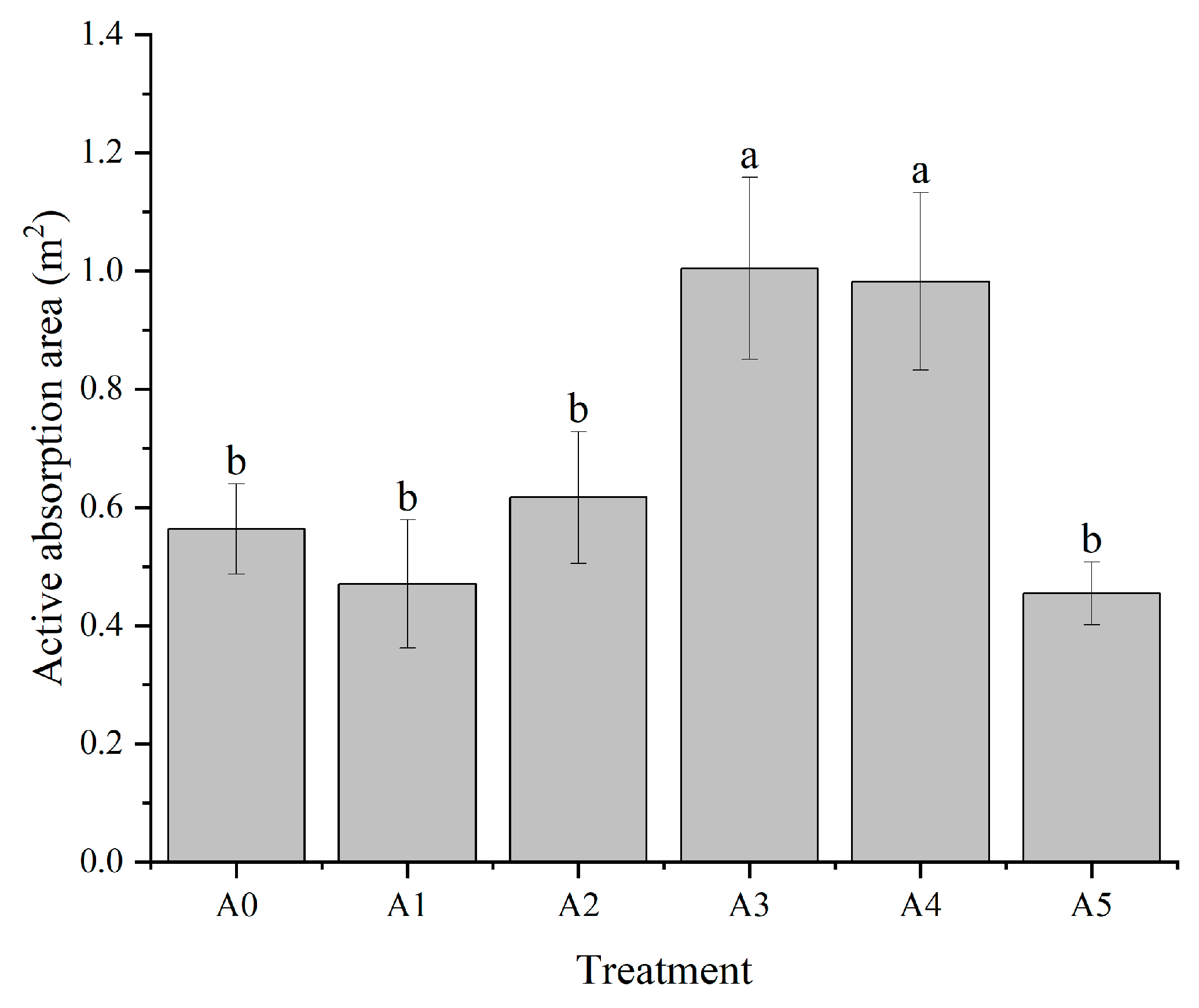
3.1.2. Effect of Nitrogen Form on the Active Absorption Area of C. oleifera Seedling Roots
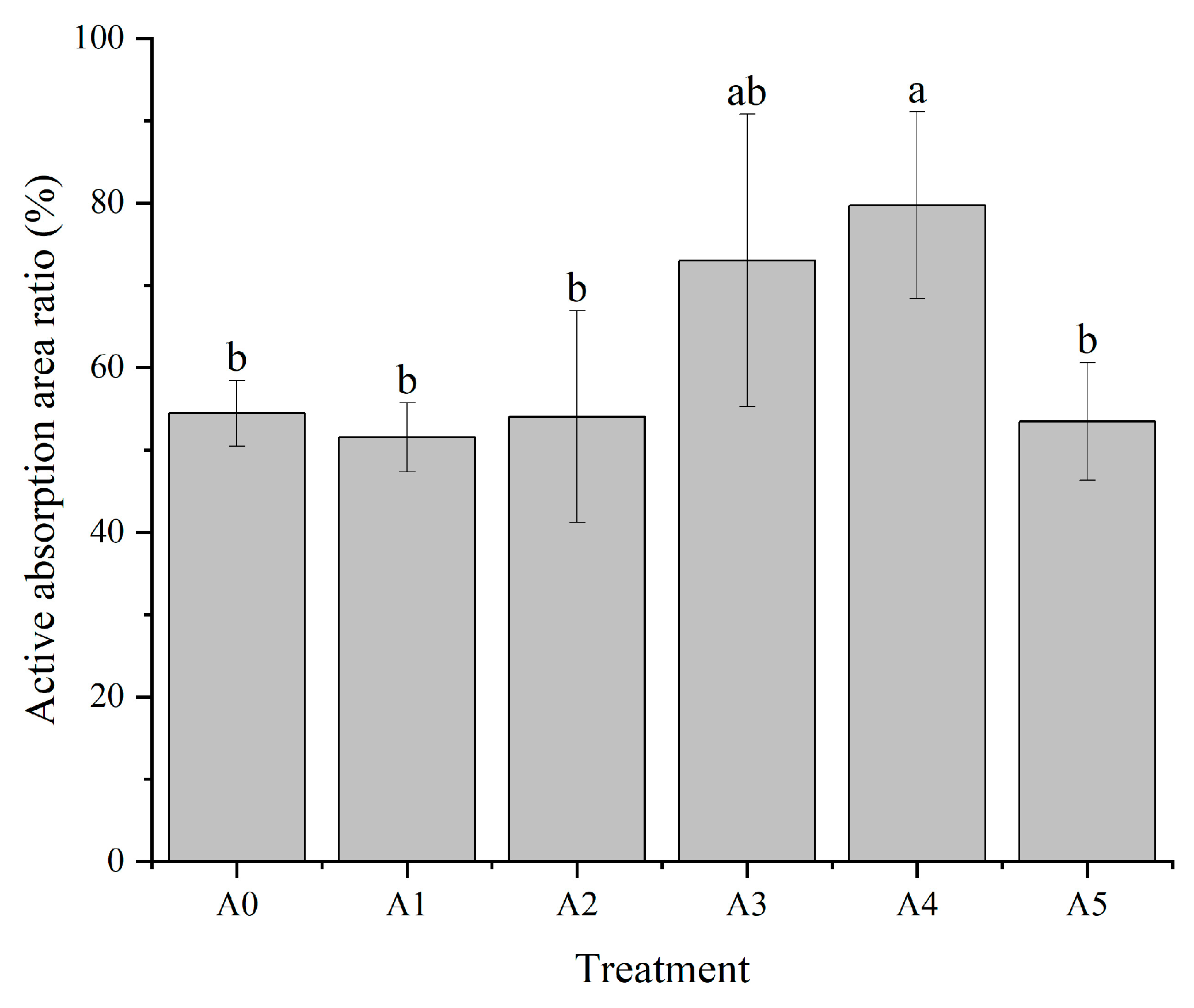
3.1.3. Effect of Nitrogen Form on the Active Absorption Area Ratio of C. oleifera Seedling Roots
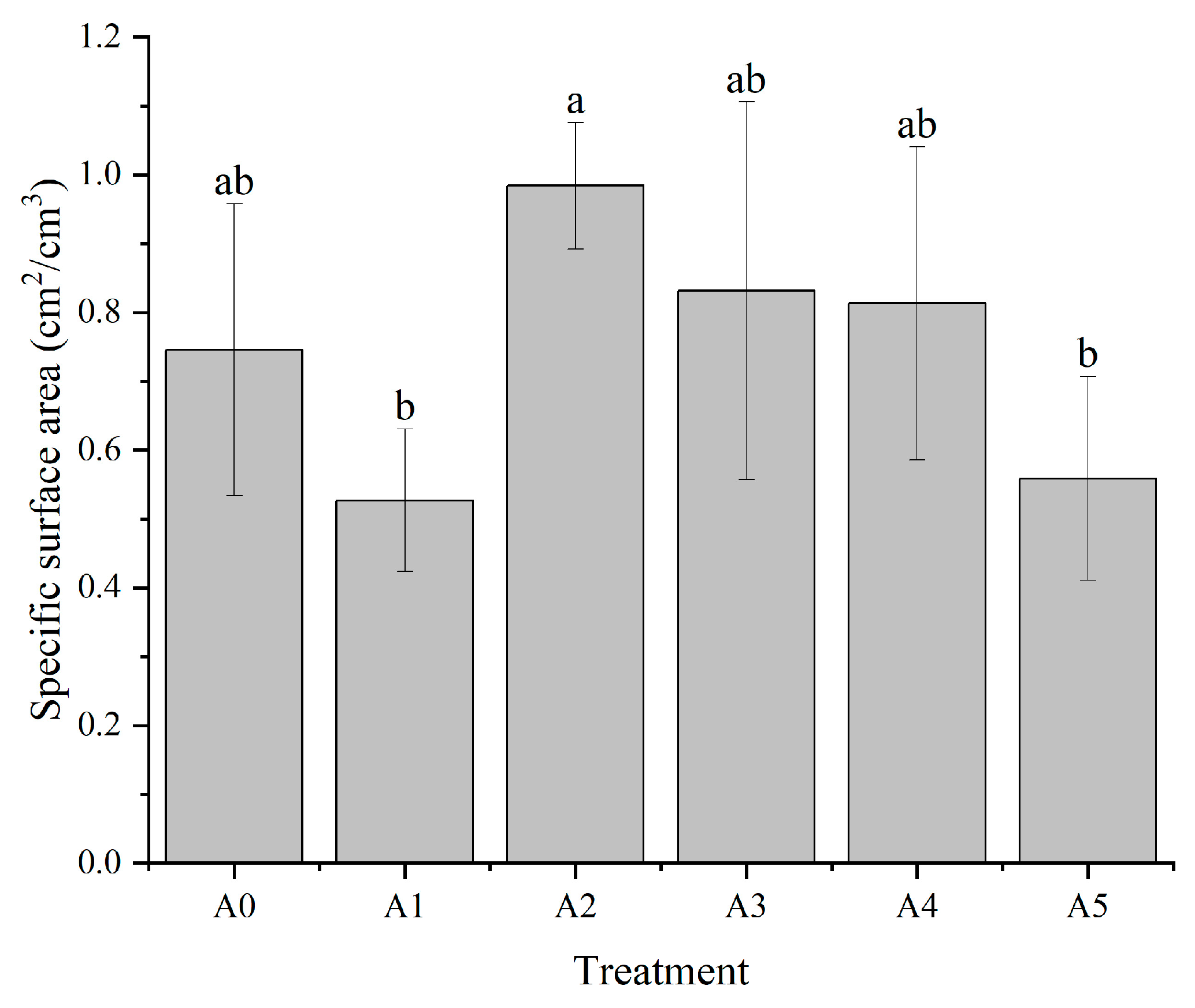
3.1.4. Effect of Nitrogen Form on the Specific Surface Area of C. oleifera Seedling Roots
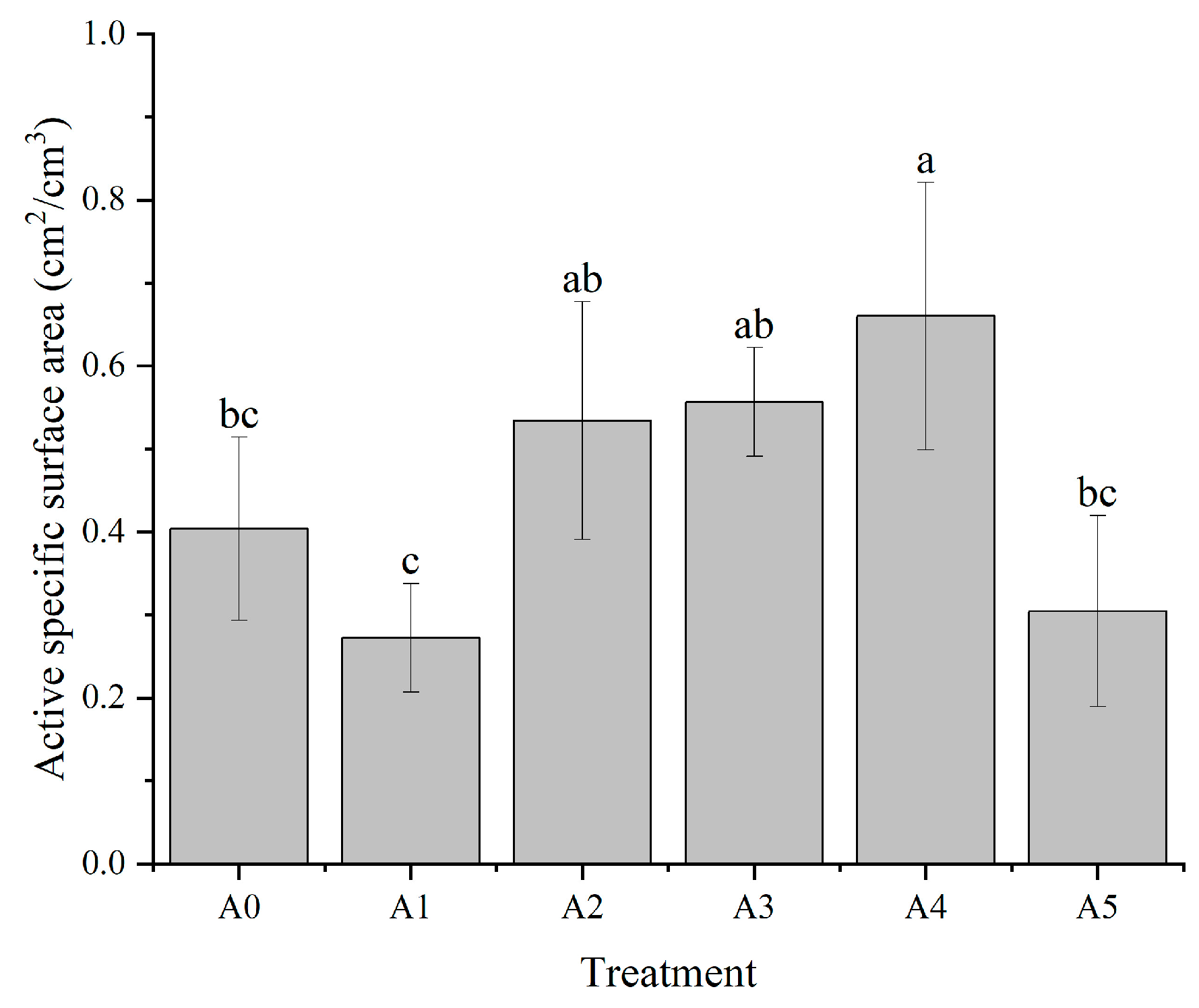
3.1.5. Effect of Nitrogen Form on the Active Specific Surface Area of C. oleifera Seedling Roots
3.2. Effects of Nitrogen Form on the Nitrogen Uptake Kinetics of C. oleifera Seedling Roots
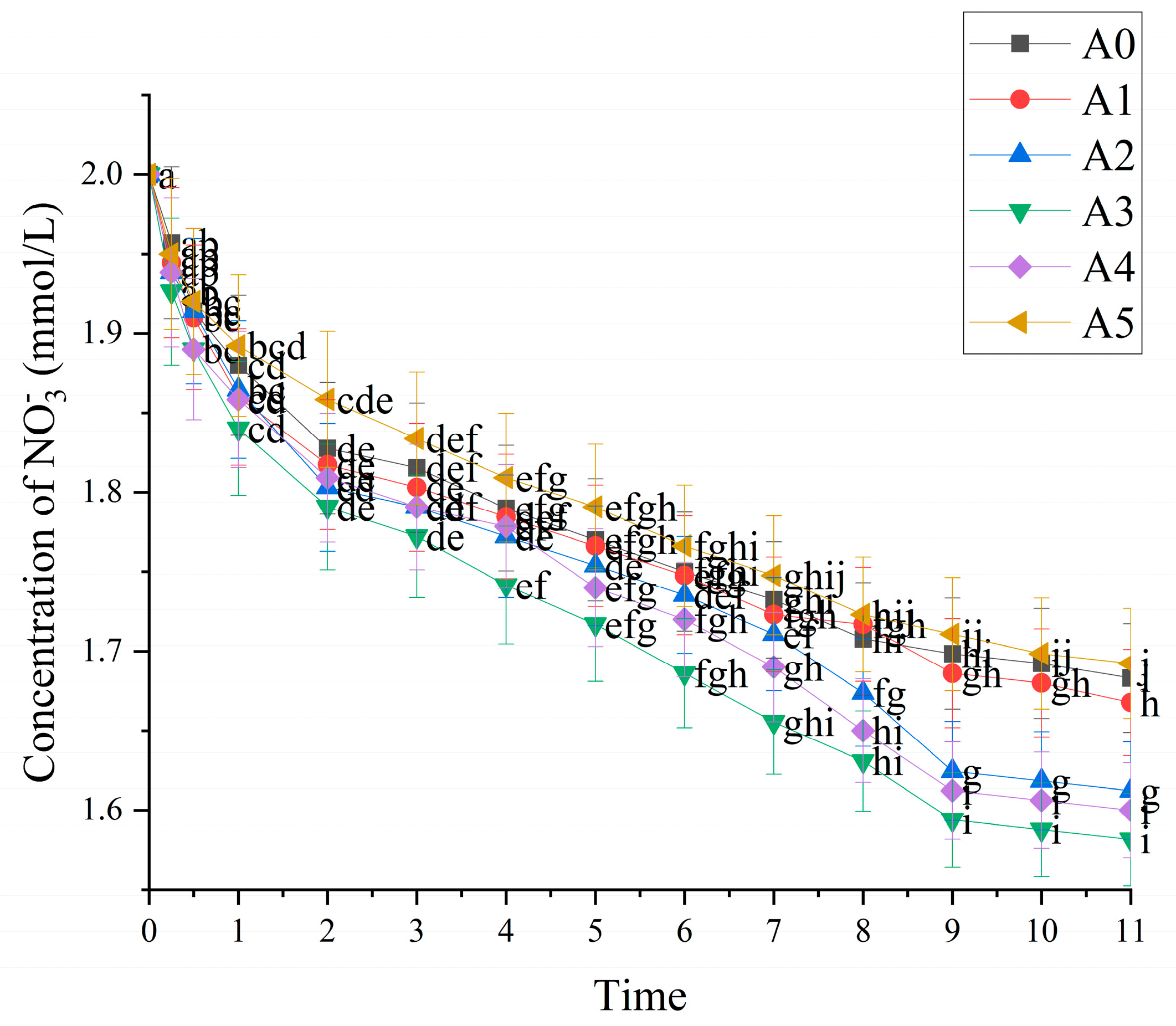
3.2.1. Effect of Nitrogen Form on Uptake Kinetics of C. oleifera Seedling Roots
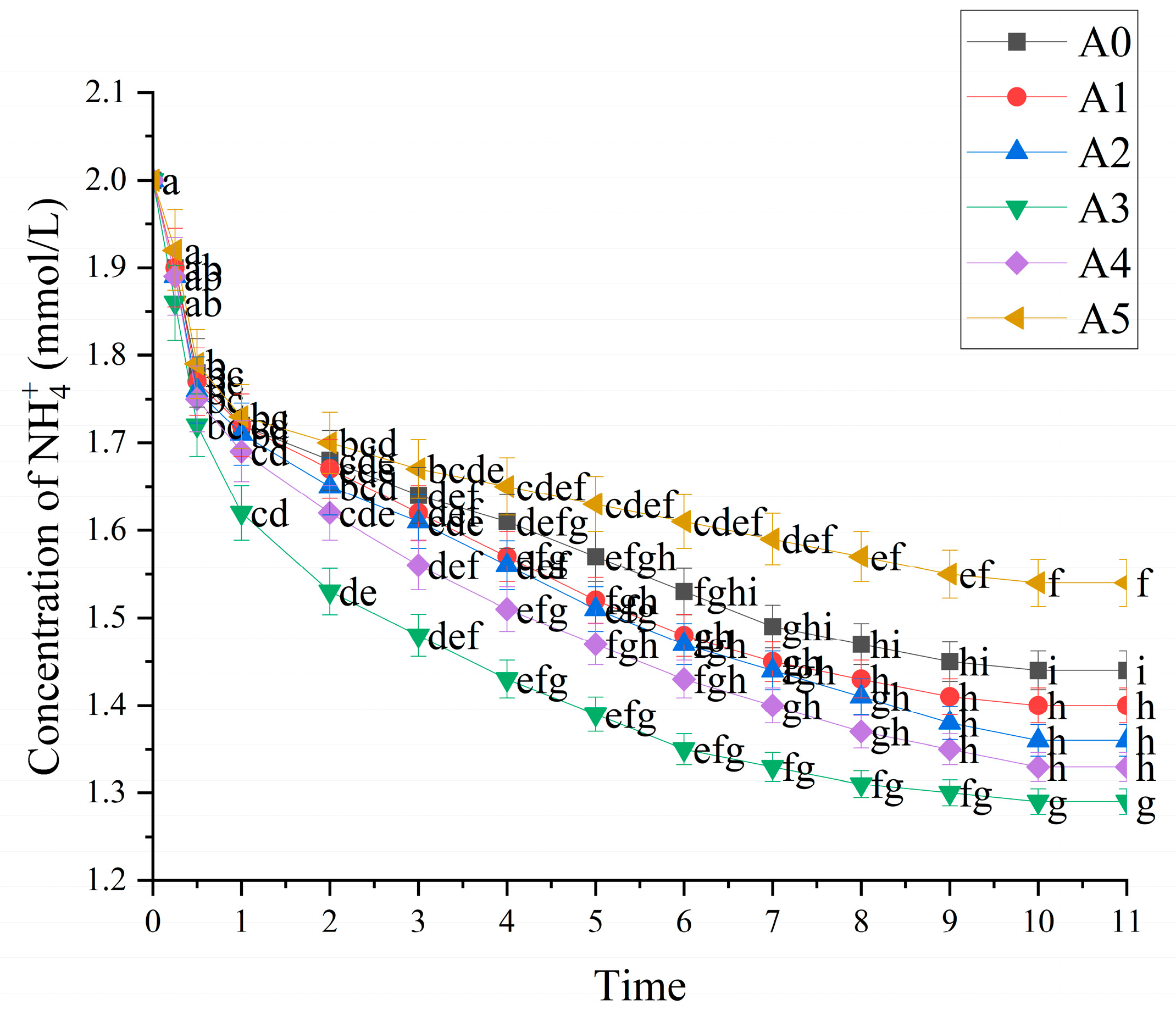
3.2.2. Effect of Nitrogen Form on Uptake Kinetics of C. oleifera Seedling Roots
4. Discussion
4.1. Effects of Nitrogen Form on the Root Activity of C. oleifera Seedlings
4.2. Effects of Nitrogen Form on and Uptake by Roots of C. oleifera Seedlings
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- He, Z.; Liu, C.; Zhang, Z.; Wang, R.; Chen, Y. Intergration of mRNA and miRNA analysis reveals the differentially regulatory network in two different Camellia oleifera cultivars under drought stress. Front. Plant Sci. 2022, 13, 1001357. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Wang, K.; Wang, Y.; Hu, Z.; Yan, C.; Huang, H.; Ma, X.; Cao, Y.; Long, W.; Liu, W.; et al. The genome of oil-Camellia and population genomics analysis provide insights into seed oil domestication. Genome Biol. 2022, 23, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Xiao, S.; Wang, L.; Liao, Z.; Chang, Y.; Mo, W.; Hu, G.; Li, W.; Zhao, G.; Zhu, H.; et al. Chromosome-level genome of Camellia lanceoleosa provides a valuable resource for understanding genome evolution and self-incompatibility. Plant J. 2022, 110, 881–898. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.F.; Huang, B.; Xu, M.; Zhou, P.Y.; Ni, Z.X.; Gong, C.; Wen, Q.; Cao, F.L.; Xu, L.A. The reference genome of camellia chekiangoleosa provides insights into camellia evolution and tea oil biosynthesis. Hortic. Res. 2022, 9, uhab083. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Liu, C.; Wang, X.; Wang, R.; Tian, Y.; Chen, Y. Leaf Transcriptome and Weight Gene Co-expression Network Analysis Uncovers Genes Associated with Photosynthetic Efficiency in Camellia oleifera. Biochem. Genet. 2021, 59, 398–421. [Google Scholar] [CrossRef]
- Wu, B.; Ruan, C.; Shah, A.H.; Li, D.; Li, H.; Ding, J.; Li, J.; Du, W. Identification of miRNA-mRNA Regulatory Modules Involved in Lipid Metabolism and Seed Development in a Woody Oil Tree (Camellia oleifera). Cells 2021, 11, 71. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Ding, X.G.; Zhang, Y.Z.; Liu, Y.J.; Cai, J.; Li, Y.Q. Effects of nutrient deficiencies on root system in Camellia gauchowensis seedlings. Nonwood For. Res. 2014, 32, 170–174. [Google Scholar]
- Inokuchi, R.; Kuma, K.I.; Miyata, T.; Okada, M. Nitrogen-assimilating enzymes in land plants and algae: Phylogenic and physiological perspectives. Physiol. Plant. 2002, 116, 1–11. [Google Scholar] [CrossRef]
- Christian, S.; Wood, C.C.; Roeb, G.W. Characterization of Arabidopsis AtAMT2, a high-affinity ammonium transporter of the plasma membrane. Plant Physiol. 2002, 130, 1788–1796. [Google Scholar]
- Sun, M.; Guo, W.S.; Zhu, X.K.; Feng, C.N.; Guo, K.Q.; Peng, Y.X. Kinetics of Nitrate and Ammonium Uptake by Different Wheat Genotypes at Seedling Stage. J. Triticeae Crops 2006, 26, 84–87. [Google Scholar]
- Nishikawa, T.; Tarutani, K.; Yamamoto, T. Nitrate and phosphate uptake kinetics of the harmful diatom Coscinodiscus wailesii, a causative organism in the bleaching of aquacultured Porphyra thalli. Harmful Algae 2010, 9, 563–567. [Google Scholar] [CrossRef]
- Epstein, E.; Hagen, C.E. A kinetic study of the absorption of alkali cations by barley roots. Plant Physiol. 1952, 27, 457–474. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.P.; Zhang, G.H. Studies on variation among wheat genotypes in Nutilization. J. Plant Nutr. Fertil. Sci. 1996, 2, 331–336. [Google Scholar]
- Huang, J.G.; Yang, B.J.; Yuan, L. Study on kinetics of absorption by various white variety. J. Plant Nutr. Fertil. Sci. 1995, 1, 38–43. [Google Scholar]
- Shi, R.H. The Principles of Plants Nutrients; Jiangsu Science and Technology Publishing House: Nanjing, China, 1989. [Google Scholar]
- Yao, Q.H.; Liu, W.D.; Chen, M.L.; Xie, Z.C. Studies on the verification of mechanistic models of nutrient(P) uptake by plant root system. J. Plant Nutr. Fertil. Sci. 1999, 5, 263–272. [Google Scholar]
- Ni, J.S.; Jiang, X.C.; Feng, X.X.; Huang, M.Q. Study on various variety of maize seedling in absorption, exudates and nitrate reductase activity. Acta Phytophysiol. Sin. 1988, 14, 188–195. [Google Scholar]
- Guo, B.H. Study on rice absorption dynamics of Si, K, Ca, and Mg. J. China West Norm. Univ. Nat. Sci. 2003, 24, 396–401. [Google Scholar]
- Jiang, T.H.; Zheng, S.J.; Shi, J.Q.; Hu, A.T.; Shi, R.H.; Xv, M. Several considerations in kinetics research on nutrients uptake by plants. J. Plant Nutr. Fertil. Sci. 1995, 1, 11–17. [Google Scholar]
- Wang, R.; Chen, L.; Wang, X.; Tang, W.; Peng, Y.; Zhang, Z.; Li, A.; Chen, Y. Effects of different proportion of nitrogen forms on the growth and physiological characteristics of Camellia oleifera seedlings. J. Nanjing For. Univ. Nat. Sci. Ed. 2019, 43, 26–32. (In Chinese) [Google Scholar]
- Lu, W.J.; Li, Y.S. Plant Physiology Experiment Tutorial; China Forestry Publishing House: Beijing, China, 2017. [Google Scholar]
- Lü, W.X.; Ge, Y.; Wu, J.Z. and Chang, J. Study on the method for the determination of nitric nitrogen, ammoniacal nitrogen and total nitrogen in plant. Spectrosc. Spectr. Anal. 2004, 24, 204–206. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis; China Agricultural Publishing House: Beijing, China, 2000; pp. 49–55. [Google Scholar]
- Hua, H.X.; Liang, Y.C.; Lou, Y.S.; Zhang, J. Comparison of research methods for silicon uptake kinetics of rice. Plant Nutr. Fertil. Sci. 2006, 12, 358–362. [Google Scholar]
- Zhai, M.P.; Jiang, S.N. Dynamics of nutrient absorption in root systems of Populus×xiao zhuanica and Robinia pseudoacacia. J. Beijing For. Univ. 2006, 28, 29–33. [Google Scholar]
- Li, D.Y.; Zhou, Y.C. Responses of seedlings growth of Pinus massoniana to calcium concentration. J. Cent. South Univ. For. Technol. 2017, 27, 39–45. [Google Scholar]
- Ma, B. Physiological and Biochemical Effects of Nitrogen Form Supply on Root Respiration of Walnut Seedlings. Master’s Thesis, Xinjiang Agriculture University, Urumqi, China, 2021. [Google Scholar]
- Ma, C. Study on Regulation of Exogenous Nitrogen Forms on Growth, Nitrogen Uptake and Metabolism of Cucumber. Ph.D. Thesis, Gansu Agriculture University, Lanzhou, China, 2020. [Google Scholar]
- Britto, D.T.; Kronzucker, H.J. toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Kronzuker, H.J.; Glass, A.D.M.; Siddiqi, M.Y. Inhibition of nitrate uptake by ammonium in barley. Plant Physiol. 1999, 120, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Lu, X.; Cao, X.; Li, J.; Xiong, J.; Xie, S. Effect of Different Nitrogen Forms on Root Growth and Dynamic Kinetics Characteristics for Citrus sinensis × Poncirus trifoliata. Sci. Silvar Sin. 2015, 51, 114–120. [Google Scholar]
- Wang, L.; Wang, W.; Huang, C.H.; Chang, C.R. Effect of Nitrate Uptake Kinetic with Different Ratios of Ammonium and Nitrate in Banana Seedlings. Chin. J. Trop. Crops 2012, 33, 988–992. [Google Scholar]
- Ren, J.; Xv, C.Y.; Lin, Y.M. Kinetic Characteristics of Different Forms of Absorbing Nitrogen in Root System of Fraxinus mandshurica Rupr. Seedling. Plant Physiol. Commun. 2008, 44, 919–922. [Google Scholar]
- Qin, S.; Li, Z.; Ma, H.; Liu, L.; Liu, G. Effects of Different Nitrogen Forms on Root Respiratory Metabolism and on Biomass in Seedlings of Cerasus sachalinensis. Acta Hortic. Sin. 2011, 38, 1021–1028. [Google Scholar]
- Du, X.H.; Peng, F.R. Effect of Inorganic Nitrogen Forms on Growth and Kinetics of Ammonium and Nitrate Uptake in Camellia sinensis L. Acta Agron. Sin. 2010, 36, 327–334. [Google Scholar] [CrossRef]
- Rennenberg, H.; Wildhagen, H.; Ehlting, B. Nitrogen nutrition of poplar trees. Plant Biol. 2010, 12, 275–291. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | m(-N)/m(-N) | Total Nitrogen/mmol·L−1 | Nitrogen Sources/mmol·L−1 | |
|---|---|---|---|---|
| NaNO3 | (NH4)2SO4 | |||
| A0(CK) | 0:0 | 0 | 0 | 0 |
| A1 | 10:0 | 8 | 8 | 0 |
| A2 | 7:3 | 8 | 5.6 | 1.2 |
| A3 | 5:5 | 8 | 4 | 2 |
| A4 | 3:7 | 8 | 2.4 | 2.8 |
| A5 | 0:10 | 8 | 0 | 4 |
| Treatment | Equation | R2 | Vmax (umol·g−1·h−1) | Km (mmol·L−1) | α |
|---|---|---|---|---|---|
| A0 | Y = 0.0013x2 − 0.044x + 2.0386 | 0.9917 | 16.10 | 1.76 | 0.009 |
| A1 | Y = 0.0014x2 − 0.0462x + 2.0373 | 0.9911 | 17.52 | 1.75 | 0.010 |
| A2 | Y = 0.0009x2 − 0.0432x + 2.0290 | 0.9847 | 16.24 | 1.54 | 0.011 |
| A3 | Y = 0.0014x2 − 0.0531x + 2.0414 | 0.9947 | 17.57 | 1.64 | 0.011 |
| A4 | Y = 0.0009x2 − 0.0441x + 2.0424 | 0.9858 | 16.29 | 1.56 | 0.011 |
| A5 | Y = 0.0013x2 − 0.0436x + 2.0525 | 0.9950 | 15.62 | 1.79 | 0.009 |
| Treatment | Equation | R2 | Vmax (umol·g−1·h−1) | Km (mmol·L−1) | α |
|---|---|---|---|---|---|
| A0 | Y = 0.0032 x2 − 0.0888x + 2.0549 | 0.9889 | 32.49 | 1.59 | 0.020 |
| A1 | Y = 0.0037x2 − 0.0993x + 2.0744 | 0.9946 | 35.01 | 1.57 | 0.022 |
| A2 | Y = 0.0034x2 − 0.0971x + 2.0632 | 0.9927 | 33.95 | 1.54 | 0.022 |
| A3 | Y = 0.0057x2 − 0.1361x + 2.0988 | 0.9924 | 45.07 | 1.49 | 0.030 |
| A4 | Y = 0.0041x2 − 0.1111x + 2.0825 | 0.9947 | 39.35 | 1.52 | 0.026 |
| A5 | Y = 0.0032x2 − 0.0793x + 2.0401 | 0.9676 | 29.62 | 1.67 | 0.018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; He, Z.; Zhang, Z.; Xv, T.; Wang, X.; Liu, C.; Chen, Y. Effects of Nitrogen Form on Root Activity and Nitrogen Uptake Kinetics in Camellia oleifera Seedlings. Forests 2023, 14, 161. https://doi.org/10.3390/f14010161
Wang R, He Z, Zhang Z, Xv T, Wang X, Liu C, Chen Y. Effects of Nitrogen Form on Root Activity and Nitrogen Uptake Kinetics in Camellia oleifera Seedlings. Forests. 2023; 14(1):161. https://doi.org/10.3390/f14010161
Chicago/Turabian StyleWang, Rui, Zhilong He, Zhen Zhang, Ting Xv, Xiangnan Wang, Caixia Liu, and Yongzhong Chen. 2023. "Effects of Nitrogen Form on Root Activity and Nitrogen Uptake Kinetics in Camellia oleifera Seedlings" Forests 14, no. 1: 161. https://doi.org/10.3390/f14010161