
Distinct Rainfall Interception Profiles among Four Common Pacific Northwest Tree Species


Abstract
:1. Introduction
2. Materials and Methods
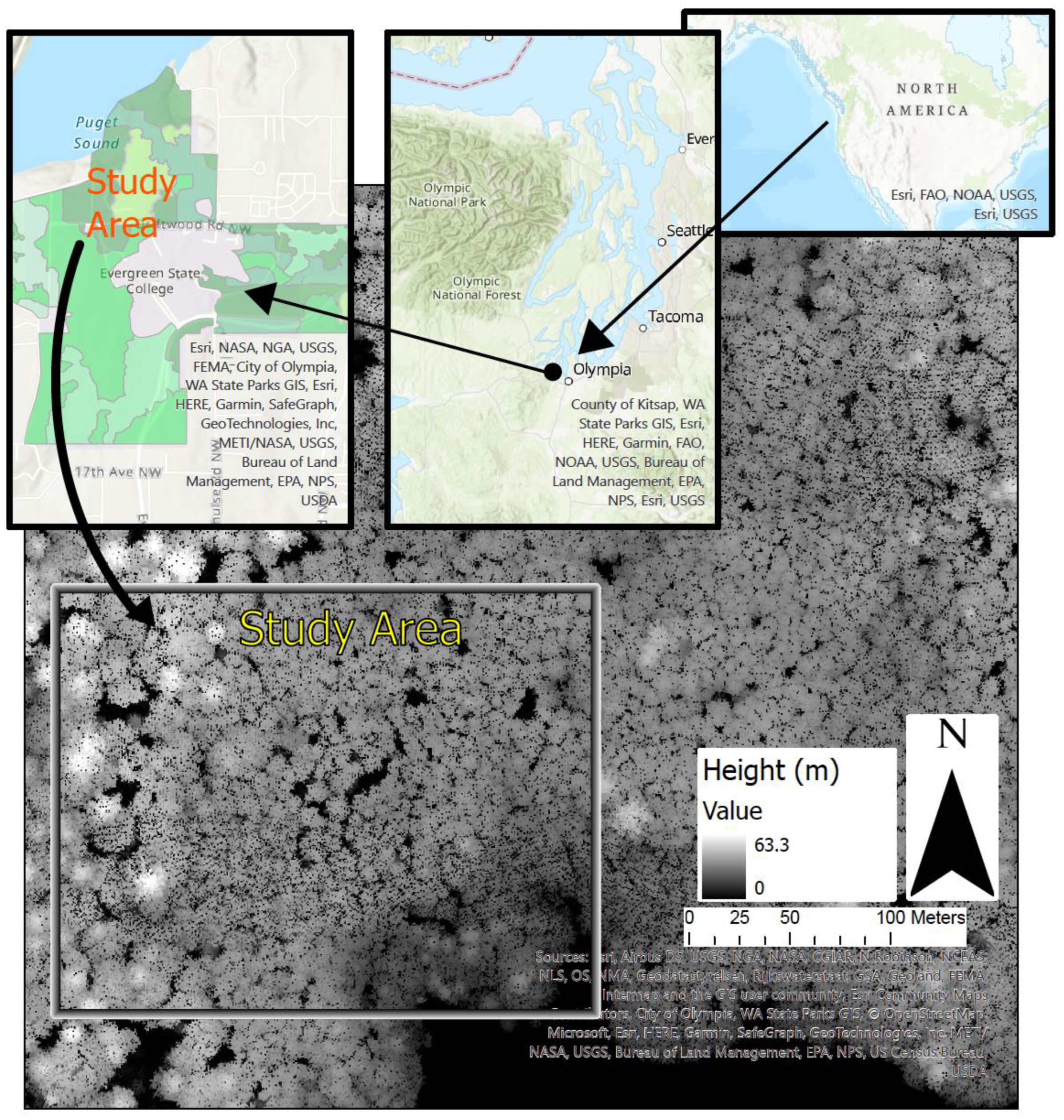
2.1. Site Characteristics
2.2. Weather Measurement
2.3. Throughfall, Soil Moisture
2.4. Statistical Analysis
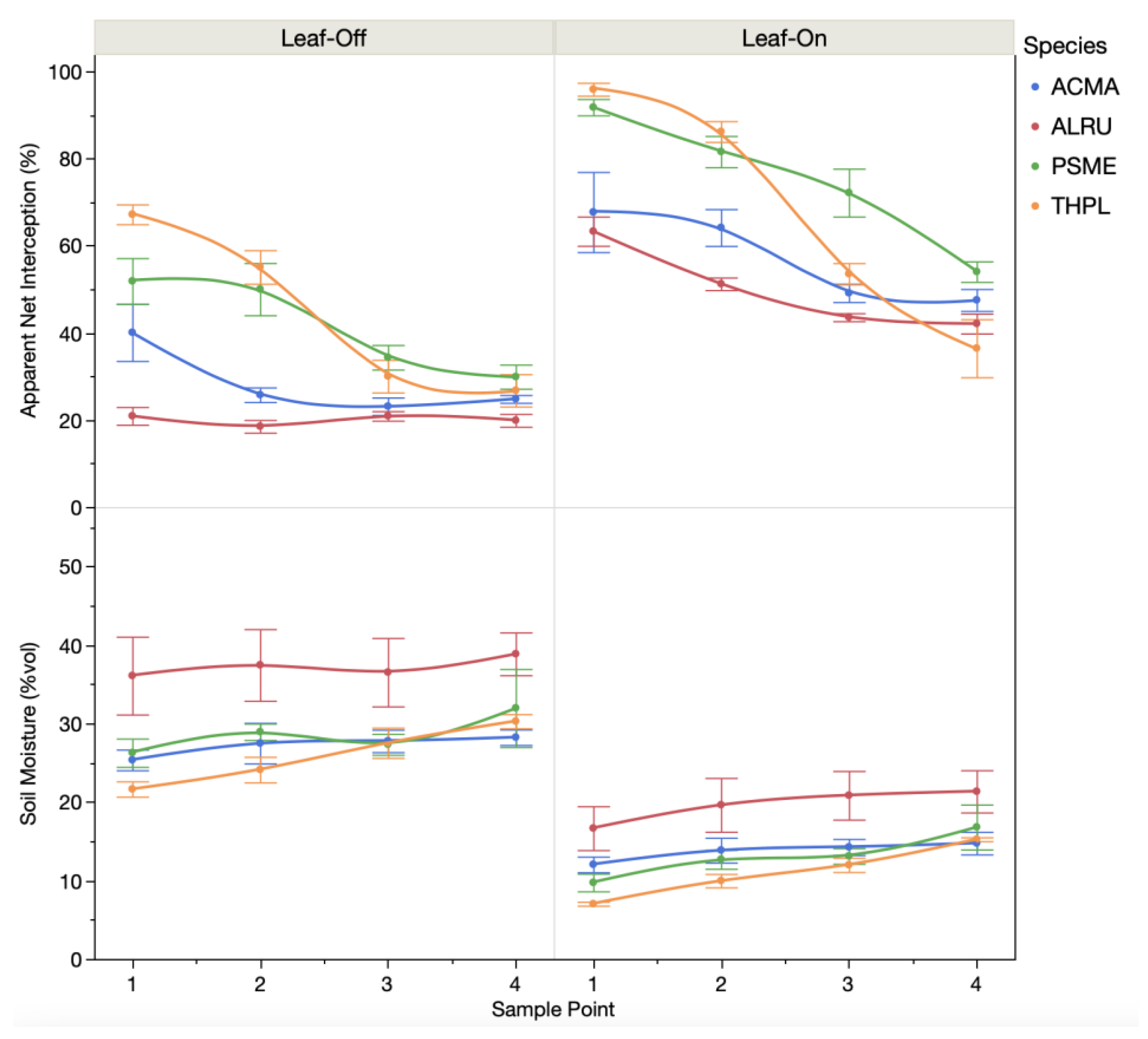
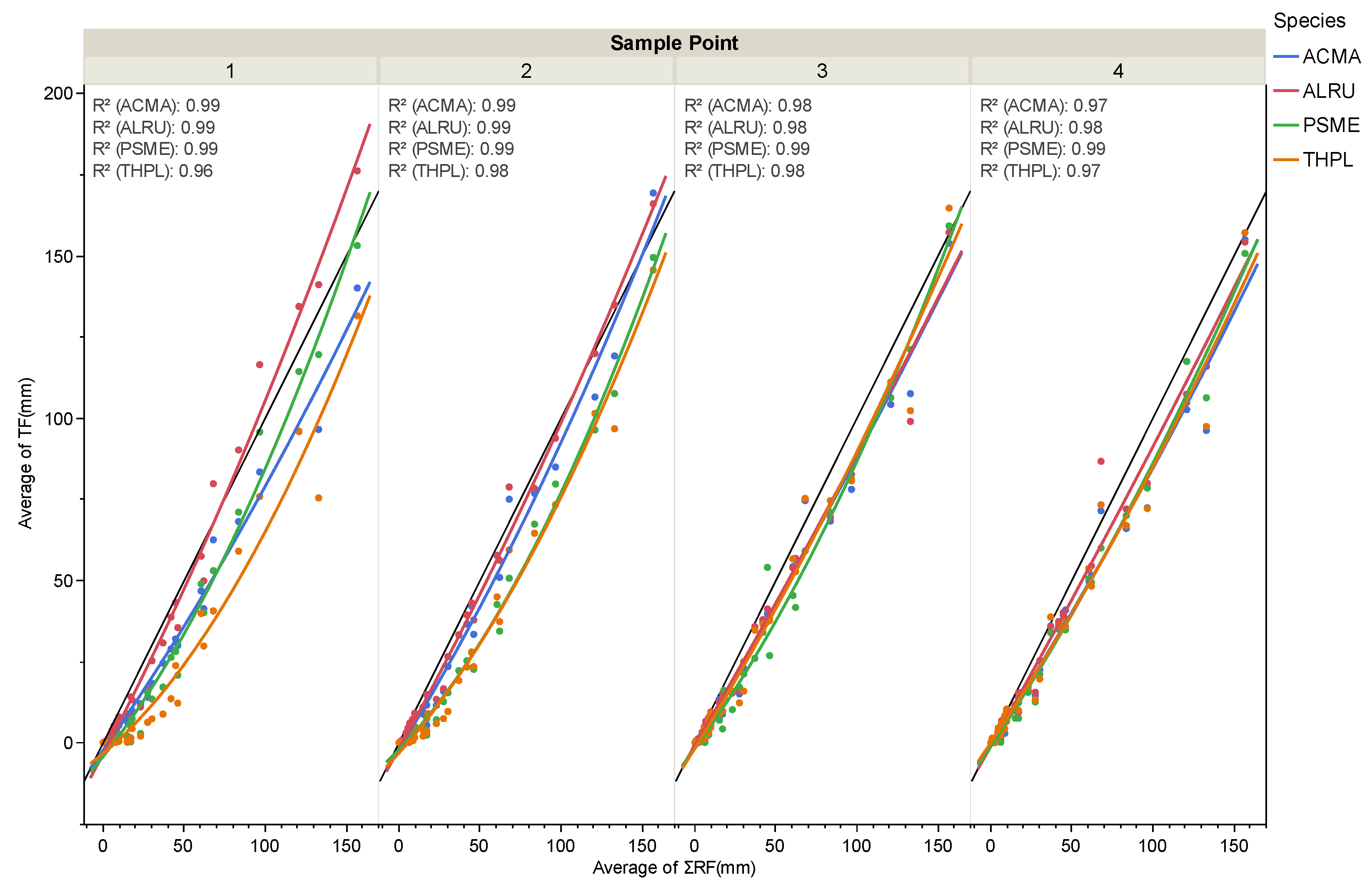
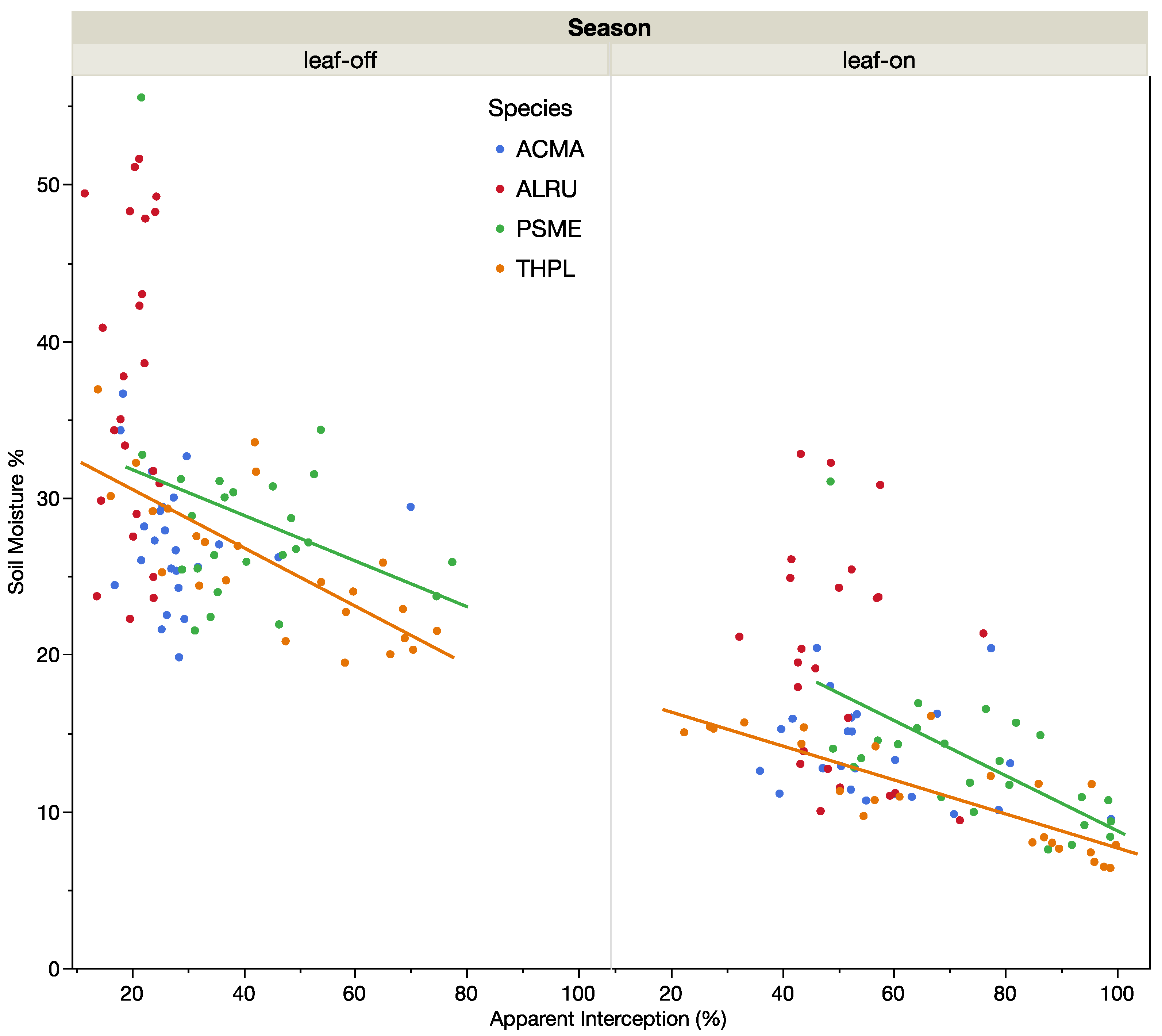
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horton, R.E. Rainfall Interception. Mon. Weather Rev. 1919, 47, 603–623. [Google Scholar] [CrossRef]
- Carlyle-Moses, D.E.; Gash, J.H.C. Rainfall Interception Loss by Forest Canopies. In Forest Hydrology and Biogeochemistry: Synthesis of Past Research and Future Directions; Ecological Studies; Levia, D.F., Carlyle-Moses, D., Tanaka, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 407–423. ISBN 978-94-007-1363-5. [Google Scholar]
- David, J.; Valente, F.; Gash, J. Evaporation of Intercepted Rainfall. In Encyclopedia of Hydrological Sciences; Anderson, M.G., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2005; pp. 627–634. ISBN 978-04-708-4894-4. [Google Scholar]
- Muzylo, A.; Llorens, P.; Valente, F.; Keizer, J.J.; Domingo, F.; Gash, J.H.C. A Review of Rainfall Interception Modelling. J. Hydrol. 2009, 370, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, S.M.M.; Gordon, D.A.; Van Stan, J.T., II. A Global Synthesis of Throughfall and Stemflow Hydrometeorology. In Precipitation Partitioning by Vegetation: A Global Synthesis; Van Stan, I., John, T., Gutmann, E., Friesen, J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 49–70. ISBN 978-3-030-29702-2. [Google Scholar]
- Asadian, Y.; Weiler, M. A New Approach in Measuring Rainfall Interception by Urban Trees in Coastal British Columbia. Water Qual. Res. J. 2009, 44, 16–25. [Google Scholar] [CrossRef]
- Crockford, R.H.; Richardson, D.P. Partitioning of Rainfall in a Eucalypt Forest and Pine Plantation in Southeastern Australia: I Throughfall Measurement in a Eucalypt Forest: Effect of Method and Species Composition. Hydrol. Process. 1990, 4, 131–144. [Google Scholar] [CrossRef]
- Llorens, P.; Poch, R.; Latron, J.; Gallart, F. Rainfall Interception by a Pinus sylvestris Forest Patch Overgrown in a Mediterranean Mountainous Abandoned Area I. Monitoring Design and Results down to the Event Scale. J. Hydrol. 1997, 199, 331–345. [Google Scholar] [CrossRef]
- Link, T.E.; Unsworth, M.; Marks, D. The Dynamics of Rainfall Interception by a Seasonal Temperate Rainforest. Agric. For. Meteorol. 2004, 124, 171–191. [Google Scholar] [CrossRef] [Green Version]
- Llorens, P.; Domingo, F. Rainfall Partitioning by Vegetation under Mediterranean Conditions. A Review of Studies in Europe. J. Hydrol. 2007, 335, 37–54. [Google Scholar] [CrossRef]
- Miralles, D.G.; Gash, J.H.; Holmes, T.R.H.; de Jeu, R.A.M.; Dolman, A.J. Global Canopy Interception from Satellite Observations. J. Geophys. Res. Atmos. 2010, 115, D16122. [Google Scholar] [CrossRef]
- Sadeghi, S.M.M.; Attarod, P.; Van Stan, J.T.; Pypker, T.G. The Importance of Considering Rainfall Partitioning in Afforestation Initiatives in Semiarid Climates: A Comparison of Common Planted Tree Species in Tehran, Iran. Sci. Total Environ. 2016, 568, 845–855. [Google Scholar] [CrossRef]
- Sadeghi, S.M.M.; Van Stan, J.T.; Pypker, T.G.; Friesen, J. Canopy Hydrometeorological Dynamics across a Chronosequence of a Globally Invasive Species, Ailanthus altissima (Mill., Tree of Heaven). Agric. For. Meteorol. 2017, 240–241, 10–17. [Google Scholar] [CrossRef]
- Sadeghi, S.M.M.; Van Stan, J.T.; Pypker, T.G.; Tamjidi, J.; Friesen, J.; Farahnaklangroudi, M. Importance of Transitional Leaf States in Canopy Rainfall Partitioning Dynamics. Eur. J. For. Res. 2018, 137, 121–130. [Google Scholar] [CrossRef]
- Fathizadeh, O.; Sadeghi, S.M.M.; Holder, C.D.; Su, L. Leaf Phenology Drives Spatio-Temporal Patterns of Throughfall under a Single Quercus castaneifolia C.A.Mey. Forests 2020, 11, 688. [Google Scholar] [CrossRef]
- Bouten, W.; Heimovaara, T.J.; Tiktak, A. Spatial Patterns of Throughfall and Soil Water Dynamics in a Douglas Fir Stand. Water Resour. Res. 1992, 28, 3227–3233. [Google Scholar] [CrossRef]
- Beier, C. Water and Element Fluxes Calculated in a Sandy Forest Soil Taking Spatial Variability into Account. For. Ecol. Manag. 1998, 101, 269–280. [Google Scholar] [CrossRef]
- Baba, M.; Okazaki, M. Spatial Variability of Soil Solution Chemistry under Hinoki Cypress (Chamaecyparis obtusa) in Tama Hills. Soil Sci. Plant Nutr. 1999, 45, 321–336. [Google Scholar] [CrossRef]
- Raat, K.J.; Draaijers, G.P.J.; Schaap, M.G.; Tietema, A.; Verstraten, J.M. Spatial Variability of Throughfall Water and Chemistry and Forest Floor Water Content in a Douglas Fir Forest Stand. Hydrol. Earth Syst. Sci. 2002, 6, 363–374. [Google Scholar] [CrossRef]
- Voigt, G.K. Distribution of Rainfall under Forest Stands. For. Sci. 1960, 6, 2–10. [Google Scholar]
- Yue, K.; De Frenne, P.; Fornara, D.A.; Van Meerbeek, K.; Li, W.; Peng, X.; Ni, X.; Peng, Y.; Wu, F.; Yang, Y.; et al. Global Patterns and Drivers of Rainfall Partitioning by Trees and Shrubs. Glob. Chang. Biol. 2021, 27, 3350–3357. [Google Scholar] [CrossRef]
- Levia, D.F.; Hudson, S.A.; Llorens, P.; Nanko, K. Throughfall Drop Size Distributions: A Review and Prospectus for Future Research. WIREs Water 2017, 4, e1225. [Google Scholar] [CrossRef] [Green Version]
- Keim, R.F.; Skaugset, A.E.; Weiler, M. Temporal Persistence of Spatial Patterns in Throughfall. J. Hydrol. 2005, 314, 263–274. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; Sumera, M.M. Old-Growth Forest Canopy Structure and Its Relationship to Throughfall Interception. For. Sci. 2004, 50, 290–298. [Google Scholar]
- Lian, X.; Zhao, W.; Gentine, P. Recent Global Decline in Rainfall Interception Loss Due to Altered Rainfall Regimes. Nat. Commun. 2022, 13, 7642. [Google Scholar] [CrossRef]
- Moore, G.W.; Bond, B.J.; Jones, J.A.; Phillips, N.; Meinzer, F.C. Structural and Compositional Controls on Transpiration in 40- and 450-Year-Old Riparian Forests in Western Oregon, USA. Tree Physiol. 2004, 24, 481–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, G.W.; Bond, B.J.; Jones, J.A. A Comparison of Annual Transpiration and Productivity in Monoculture and Mixed-Species Douglas-Fir and Red Alder Stands. For. Ecol. Manag. 2011, 262, 2263–2270. [Google Scholar] [CrossRef]
- Ryan, M.G.; Yoder, B.J. Hydraulic Limits to Tree Height and Tree Growth. BioScience 1997, 47, 235–242. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Phillips, N.; Lunch, C.; Bond, B.J.; Ryan, M.G. An Investigation of Hydraulic Limitation and Compensation in Large, Old Douglas-Fir Trees. Tree Physiol. 2002, 22, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Massman, W.J. The Derivation and Validation of a New Model for the Interception of Rainfall by Forests. Agric. Meteorol. 1983, 28, 261–286. [Google Scholar] [CrossRef]
- Bond, B.J.; Jones, J.A.; Moore, G.; Phillips, N.; Post, D.; McDonnell, J.J. The Zone of Vegetation Influence on Baseflow Revealed by Diel Patterns of Streamflow and Vegetation Water Use in a Headwater Basin. Hydrol. Process. 2002, 16, 1671–1677. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Goward, T.; Asplund, J. Canopy Throughfall Links Canopy Epiphytes to Terrestrial Vegetation in Pristine Conifer Forests. Fungal Ecol. 2021, 52, 101075. [Google Scholar] [CrossRef]
- Zinke, P.J. The Pattern of Influence of Individual Forest Trees on Soil Properties. Ecology 1962, 43, 130–133. [Google Scholar] [CrossRef]
- Fischer, D.G.; Hart, S.C.; Whitham, T.G.; Martinsen, G.D.; Keim, P. Ecosystem Implications of Genetic Variation in Water-Use of a Dominant Riparian Tree. Oecologia 2004, 139, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.G.; Dickson, B.G.; Whitham, T.G.; Hart, S.C. Self-Similarity, Leaf Litter Traits, and Neighborhood Predicting Fine Root Dynamics in a Common-Garden Forest. Front. Environ. Sci. 2019, 7, 142. [Google Scholar] [CrossRef]
- Sensoy, H.; Tanyel, M. Effect of Heavy Rain Conditions on Throughfall in Evergreens and Conifers in Urban Settings. Pol. J. Environ. Stud. 2021, 31, 271–279. [Google Scholar] [CrossRef]
- The Evergreen State College. The Evergreen State College Weather Station. Available online: https://weather.evergreen.edu/ (accessed on 12 December 2022).
- Kirsch, J.L.; Fischer, D.G.; Kazakova, A.N.; Biswas, A.; Kelm, R.E.; Carlson, D.W.; LeRoy, C.J. Diversity-Carbon Flux Relationships in a Northwest Forest. Diversity 2012, 4, 33–58. [Google Scholar] [CrossRef] [Green Version]
- Rex, I.; Fischer, D.G.; Bartlett, R. A Decade of Understory Community Dynamics and Stability in a Mature Second-Growth Forest in Western Washington. Northwest Sci. 2023; in press. [Google Scholar]
- Waring, B.; Alvarez-Cansino, L.; Barry, K.; Becklund, K.; Dale, S.; Gei, M.; Keller, A.; Lopez, O.; Markesteijn, L.; Mangan, S.; et al. Pervasive and Strong Effects of Plants on Soil Chemistry: A Meta-Analysis of Individual Plant ‘Zinke’ Effects. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151001. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.E.; Boyer, E.W.; Dent, C.L.; Gergel, S.E.; Grimm, N.B.; Groffman, P.M.; Hart, S.C.; Harvey, J.W.; Johnston, C.A.; Mayorga, E.; et al. Biogeochemical Hot Spots and Hot Moments at the Interface of Terrestrial and Aquatic Ecosystems. Ecosystems 2003, 6, 301–312. [Google Scholar] [CrossRef]
- Levia, D.; Frost, E. Variability of Throughfall Volume and Solute Inputs in Wooded Ecosytems. Prog. Phys. Geogr. 2006, 30, 605–632. [Google Scholar] [CrossRef]
- Groffman, P.M.; Butterbach-Bahl, K.; Fulweiler, R.W.; Gold, A.J.; Morse, J.L.; Stander, E.K.; Tague, C.; Tonitto, C.; Vidon, P. Challenges to Incorporating Spatially and Temporally Explicit Phenomena (Hotspots and Hot Moments) in Denitrification Models. Biogeochemistry 2009, 93, 49–77. [Google Scholar] [CrossRef]
- Pypker, T.; Unsworth, M.; Bond, B. The Role of Epiphytes in Rainfall Interception by Forests in the Pacific Northwest. I. Laboratory Measurements of Water Storage. Can. J. For. Res. Rev. Can. Rech. For. 2006, 36, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Nadkarni, N.M. Biomass and Mineral Capital of Epiphytes in an Acer macrophyllum Community of a Temperate Moist Coniferous Forest, Olympic Peninsula, Washington State. Can. J. Bot. 1984, 62, 2223–2228. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Pypker, T.G. A Review and Evaluation of Forest Canopy Epiphyte Roles in the Partitioning and Chemical Alteration of Precipitation. Sci. Total Environ. 2015, 536, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Peck, J.E. The Association of Commercially Harvestable Bryophytes and Their Host Species in Northwestern Oregon. Bryologist 1997, 100, 383–393. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Max Cover (%), Distance from Tree (m) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Catchment Point | |||||||||
| Species | DBH 1 (cm) | 1 | 2 | 3 | 4 | ||||
| Acer macrophyllum | 41.3 | 86.3 | 0.2 | 86.0 | 2.2 | 87.0 | 4.1 | 87.3 | 6.0 |
| 33.3 | 86.4 | 0.4 | 86.4 | 2.3 | 85.5 | 4.2 | 85.8 | 6.2 | |
| 30.4 | 87.3 | 0.5 | 84.1 | 2.3 | 82.6 | 3.9 | 83.2 | 5.7 | |
| 39.3 | 80.9 | 0.4 | 86.5 | 2.6 | 82.2 | 4.4 | 82.5 | 6.3 | |
| 29.4 | 79.9 | 0.3 | 80.6 | 1.5 | 83.4 | 2.6 | 87.4 | 3.8 | |
| 51.1 | 83.3 | 0.5 | 81.6 | 2.1 | 84.7 | 3.6 | 84.1 | 5.1 | |
| Alnus rubra | 39.7 | 81.6 | 0.3 | 81.5 | 1.6 | 80.3 | 3.0 | 80.8 | 4.1 |
| 32.2 | 87.1 | 0.3 | 84.3 | 1.0 | 82.4 | 1.8 | 81.4 | 2.3 | |
| 33.4 | 77.7 | 0.2 | 77.4 | 1.1 | 77.1 | 1.9 | 77.7 | 4.2 | |
| 36.9 | 86.0 | 0.4 | 84.3 | 1.5 | 83.4 | 2.6 | 82.9 | 4.0 | |
| 38.2 | 83.2 | 0.4 | 82.1 | 1.5 | 80.8 | 2.5 | 80.7 | 3.6 | |
| 33.4 | 83.7 | 0.4 | 82.8 | 1.4 | 82.4 | 2.7 | 82.8 | 4.0 | |
| Pseudotsuga menziesii | 42.2 | 91.6 | 0.3 | 87.6 | 1.5 | 88.8 | 2.8 | 54.3 | 4.2 |
| 42.1 | 84.8 | 0.3 | 86.8 | 1.8 | 85.7 | 3.0 | 84.4 | 4.4 | |
| 36.2 | 83.8 | 0.2 | 83.6 | 1.8 | 86.7 | 3.3 | 86.7 | 4.7 | |
| 47.8 | 84.1 | 0.3 | 83.9 | 1.7 | 85.1 | 3.0 | 84.2 | 4.4 | |
| 52 | 85.2 | 0.3 | 84.8 | 2.2 | 86.5 | 4.0 | 87.0 | 5.9 | |
| 69.8 | 85.3 | 0.5 | 84.7 | 1.5 | 83.2 | 2.6 | 81.4 | 3.7 | |
| Thuja plicata | 73.2 | 87.2 | 0.6 | 81.7 | 2.6 | 68.8 | 4.3 | 58.8 | 6.5 |
| 58.8 | 93.5 | 0.3 | 91.2 | 2.5 | 78.6 | 4.2 | 77.0 | 6.0 | |
| 107.4 | 87.3 | 0.4 | 82.6 | 2.1 | 80.3 | 3.9 | 73.2 | 5.4 | |
| 62.3 | 86.9 | 0.5 | 90.2 | 2.6 | 89.7 | 4.9 | 87.8 | 7.3 | |
| 113.7 | 89.9 | 0.5 | 89.7 | 2.0 | 90.4 | 3.7 | 87.8 | 5.2 | |
| 85.2 | 89.3 | 0.4 | 87.8 | 2.0 | 85.1 | 3.6 | 84.7 | 5.1 | |
| Catchment Sample Point | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 |  |  |  |  |  |  |  | |||||||||
| 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |
| A. macrophyllum | A. rubra | P. menziesii | T. plicata | |||||||||||||
| Leaf-Off | A | A | A | A | ||||||||||||
| B | B | B | B | |||||||||||||
| C | C | C | C | C | C | C | ||||||||||
| D | D | D | D | D | D | D | D | D | D | D | ||||||
| Leaf-On | A | A | A | A | ||||||||||||
| B | B | B | B | |||||||||||||
| C | C | C | C | C | ||||||||||||
| D | D | D | D | D | ||||||||||||
| E | E | E | E | E | E | |||||||||||
| F | F | F | F | F | F | F | ||||||||||
| G | G | G | G | G | G | G | G | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fischer, D.G.; Vieira, S.T.; Jayakaran, A.D. Distinct Rainfall Interception Profiles among Four Common Pacific Northwest Tree Species. Forests 2023, 14, 144. https://doi.org/10.3390/f14010144
Fischer DG, Vieira ST, Jayakaran AD. Distinct Rainfall Interception Profiles among Four Common Pacific Northwest Tree Species. Forests. 2023; 14(1):144. https://doi.org/10.3390/f14010144
Chicago/Turabian StyleFischer, Dylan G., Spencer T. Vieira, and Anand D. Jayakaran. 2023. "Distinct Rainfall Interception Profiles among Four Common Pacific Northwest Tree Species" Forests 14, no. 1: 144. https://doi.org/10.3390/f14010144