
Correlation between the Concentration of Secondary Metabolites and Soil Microorganisms in Sophora Koreensis Nakai from Different Habitat


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Collection and Soil Sampling
2.2. Analysis of Compounds in Leaves, Stems and Roots
2.3. Soil Microorganism Community Analysis
2.4. Statistical Analysis
3. Results and Discussion
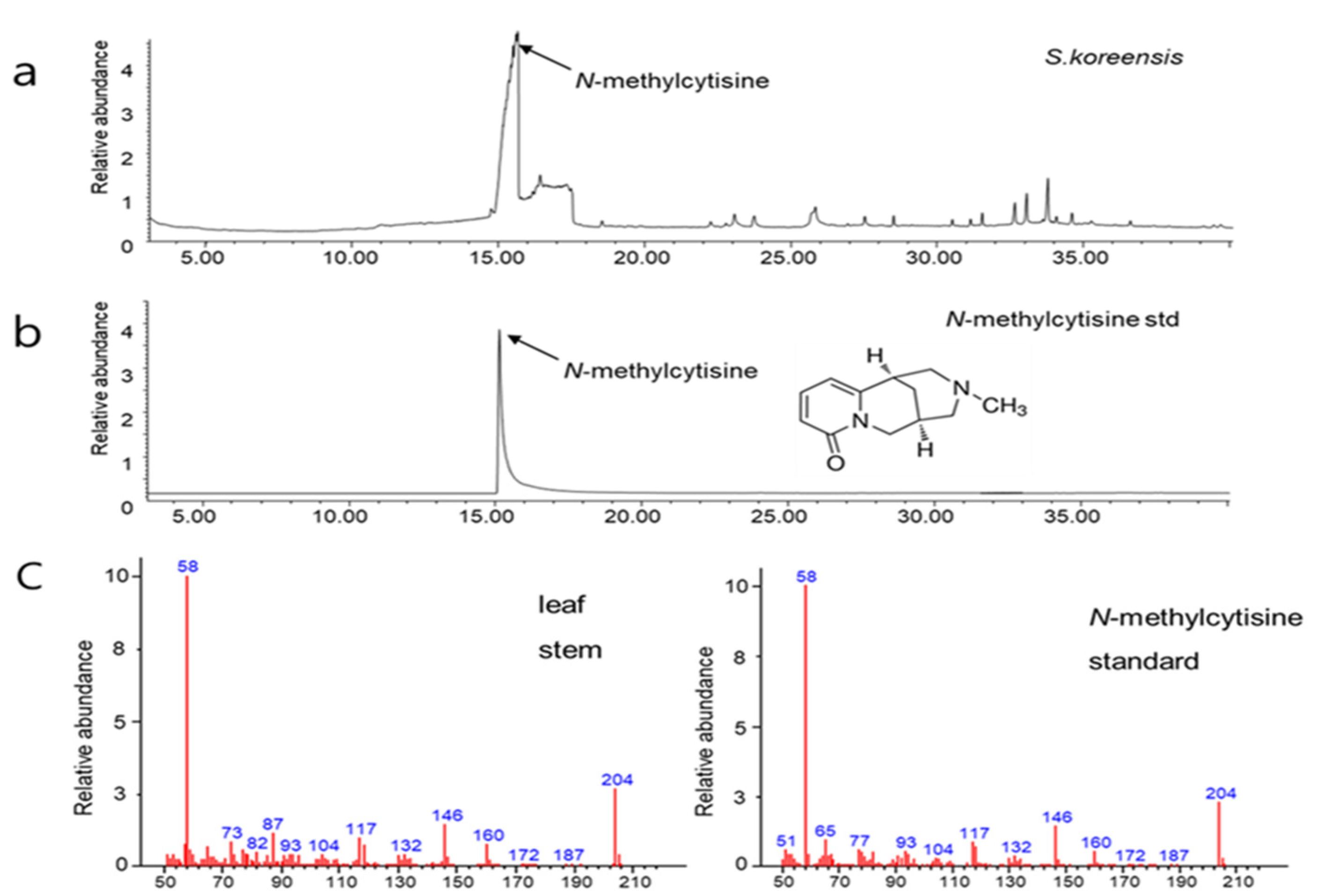
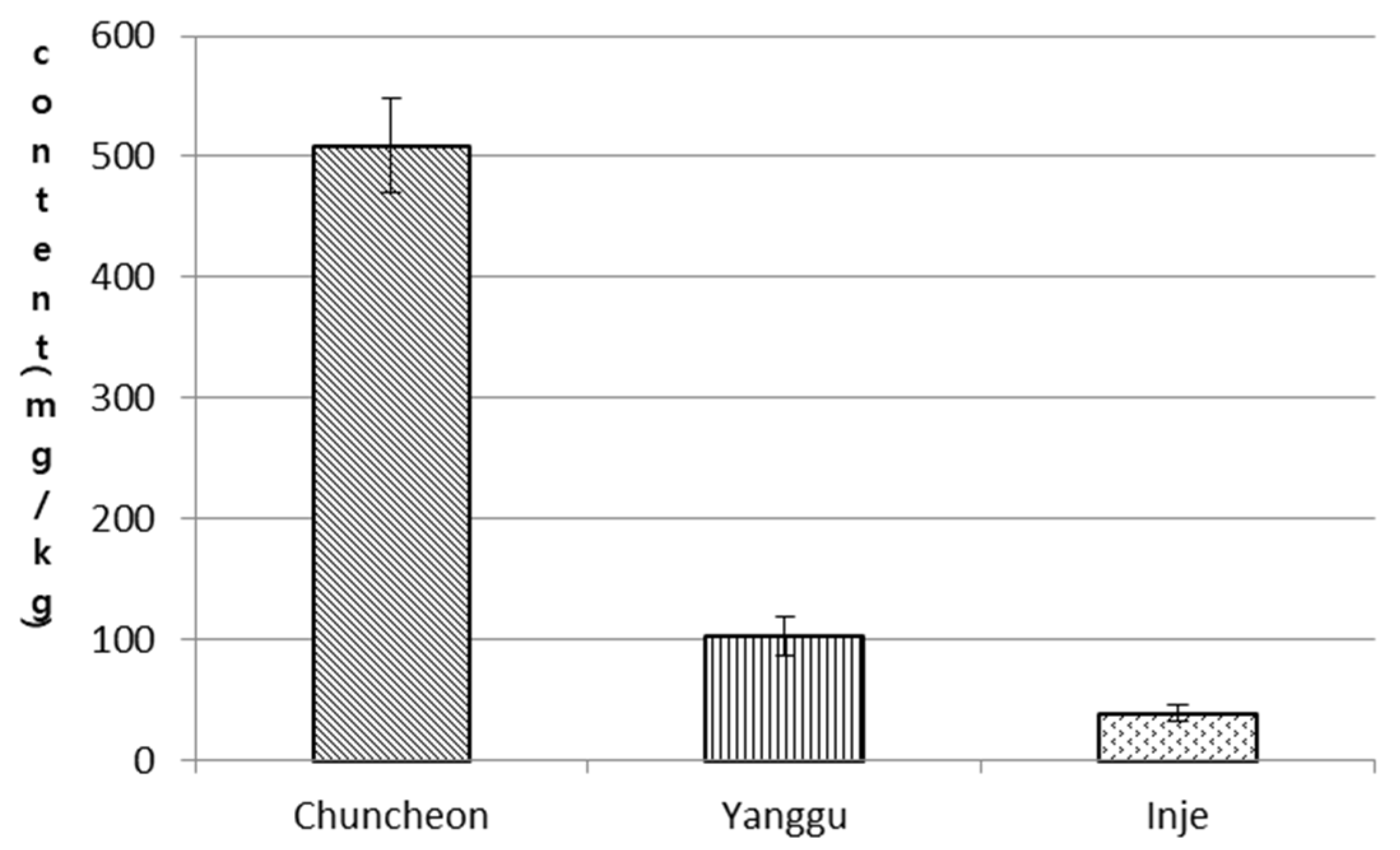
3.1. Comparison of Secondary Compounds in Parts of S. koreensis Plants from Different Habitats
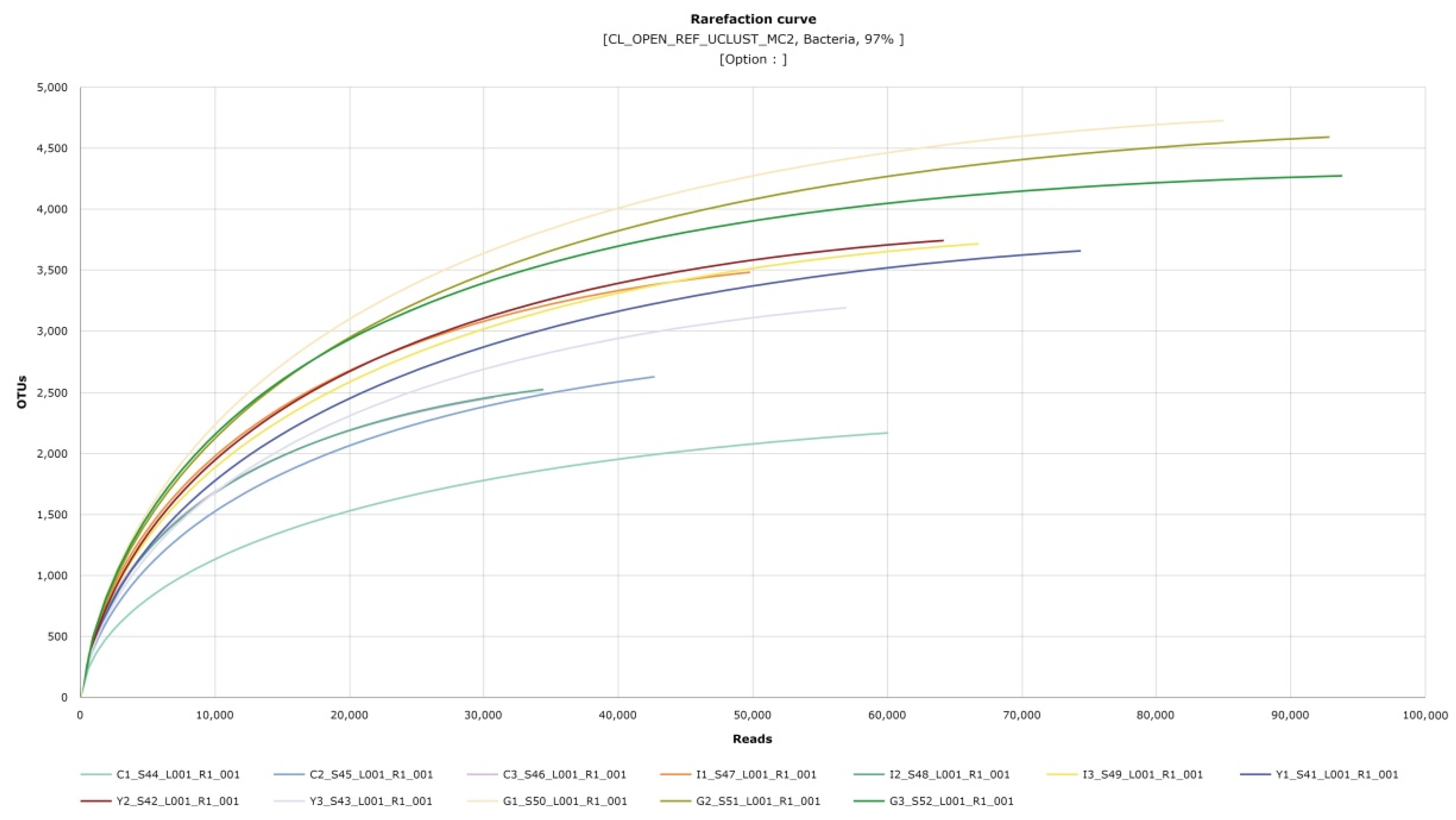
3.2. Soil Microbial Analysis by Habitat
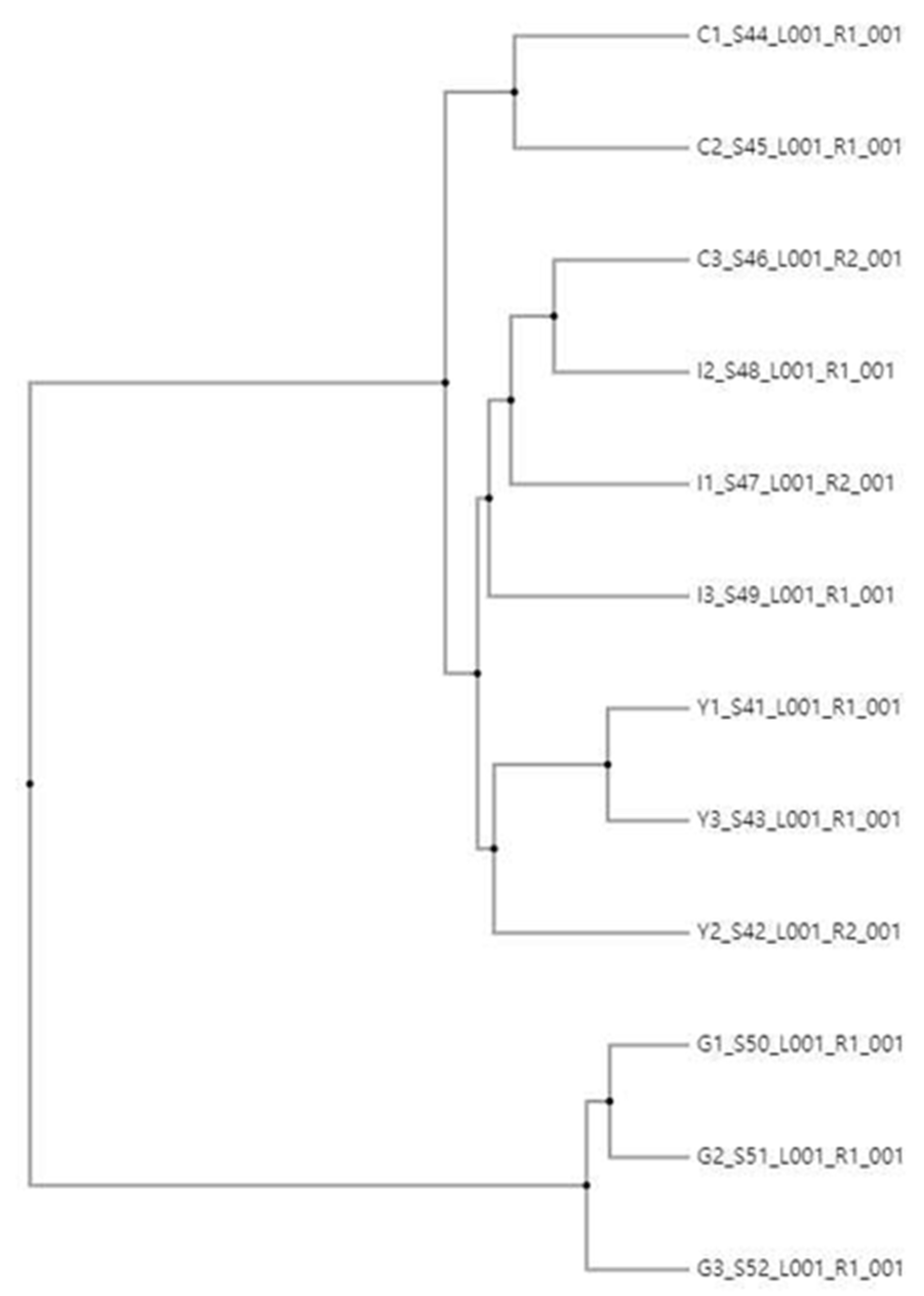
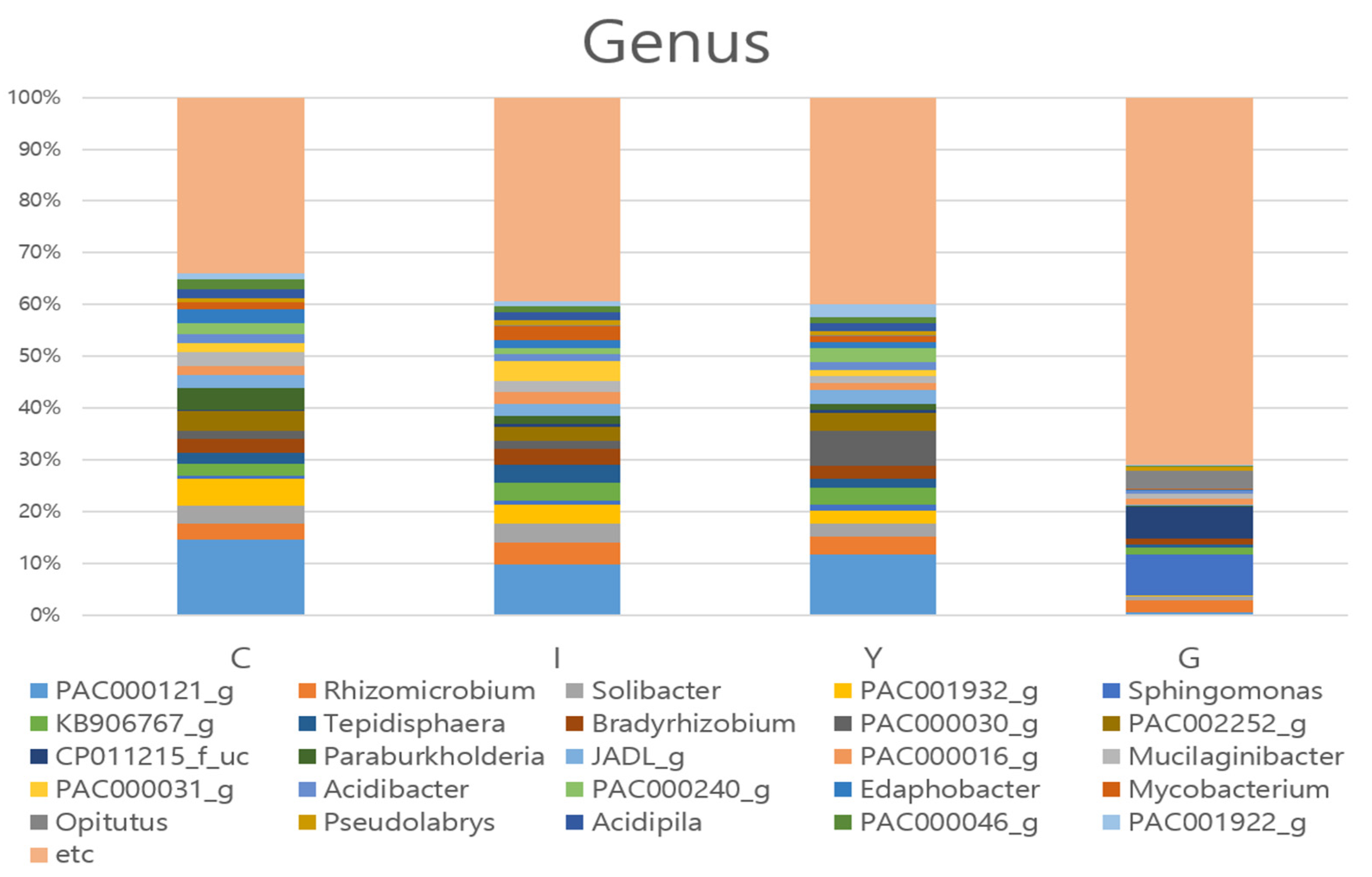
3.3. Comparison of Bacterial Communities at the Genus Level
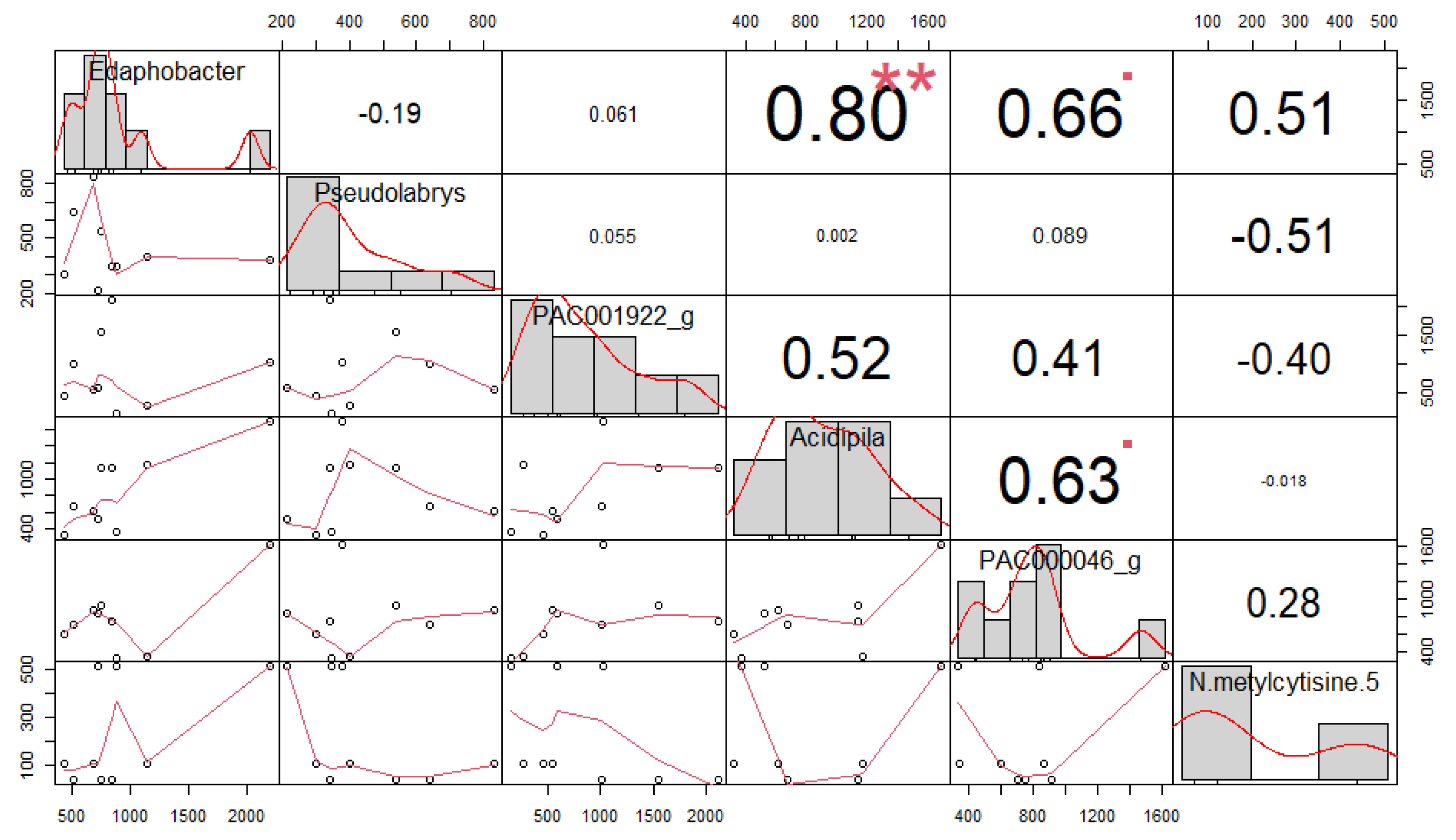
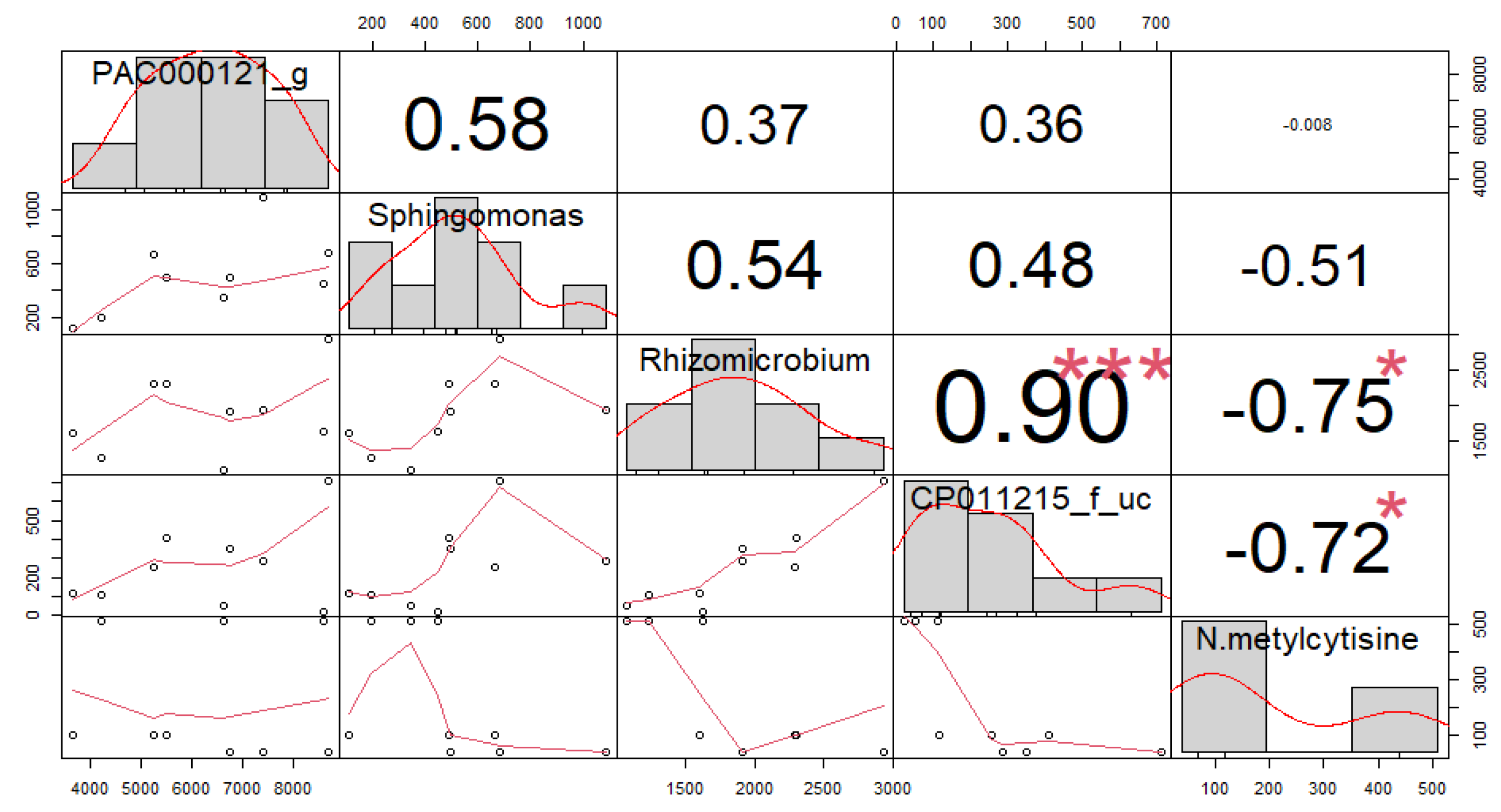
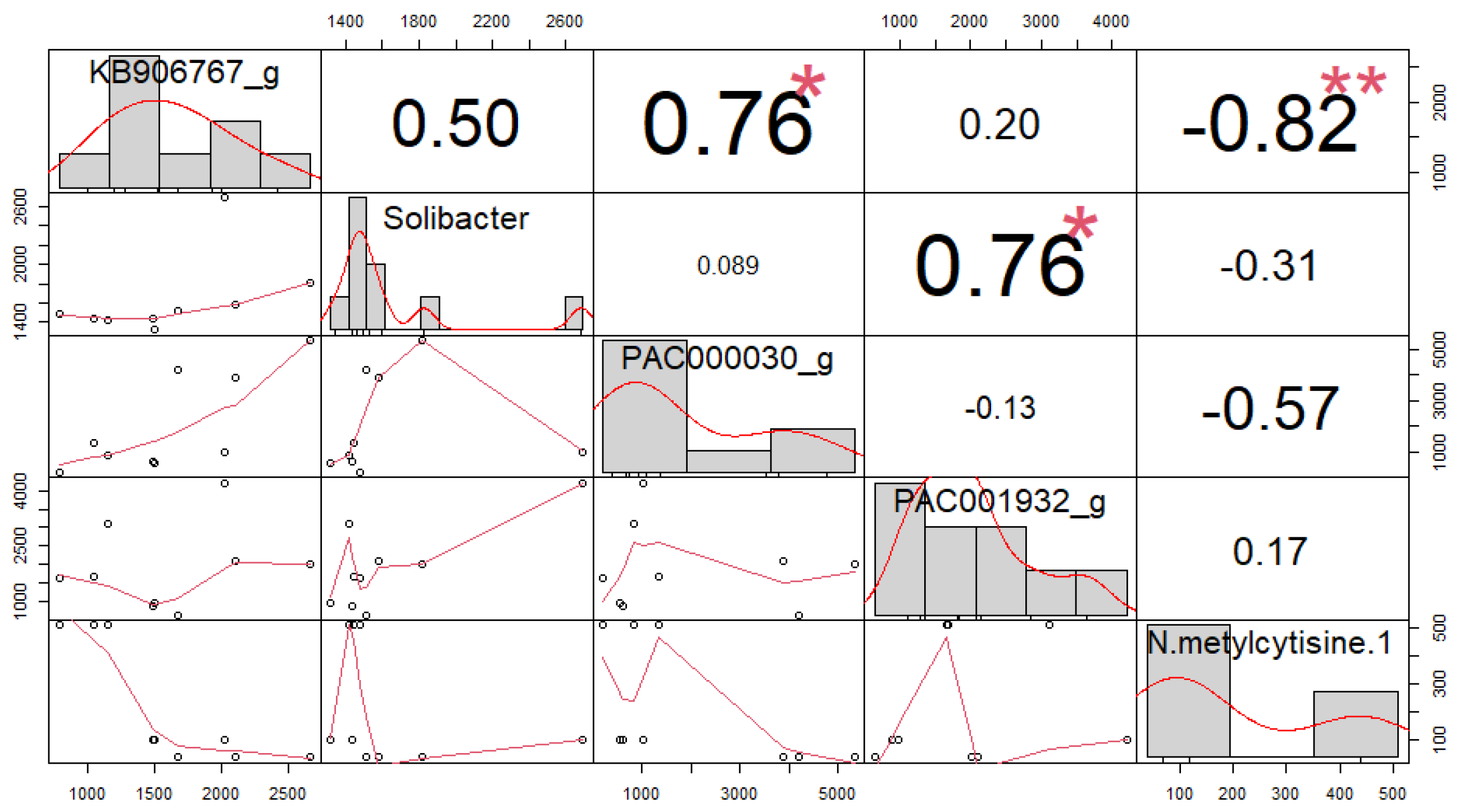
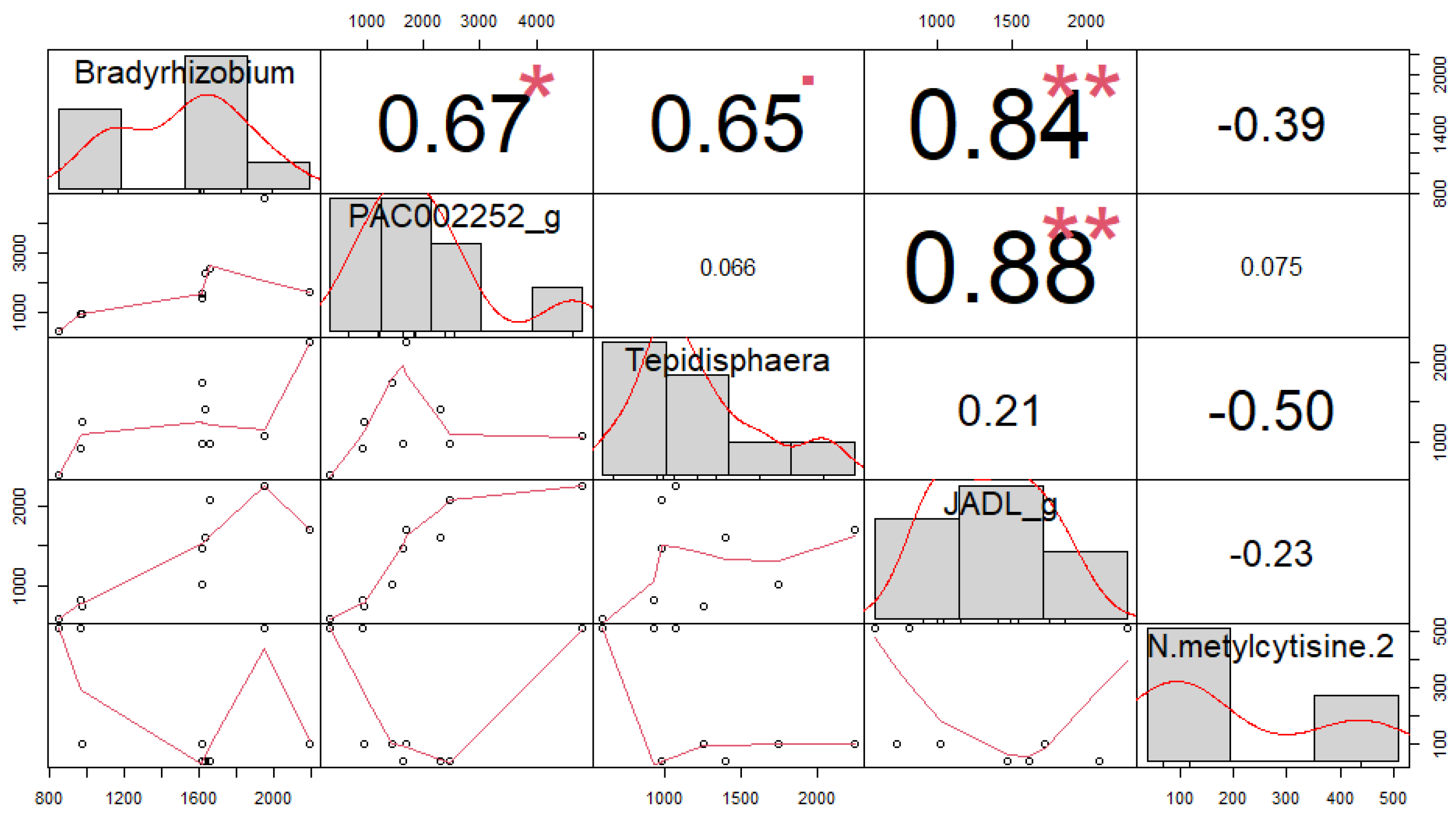
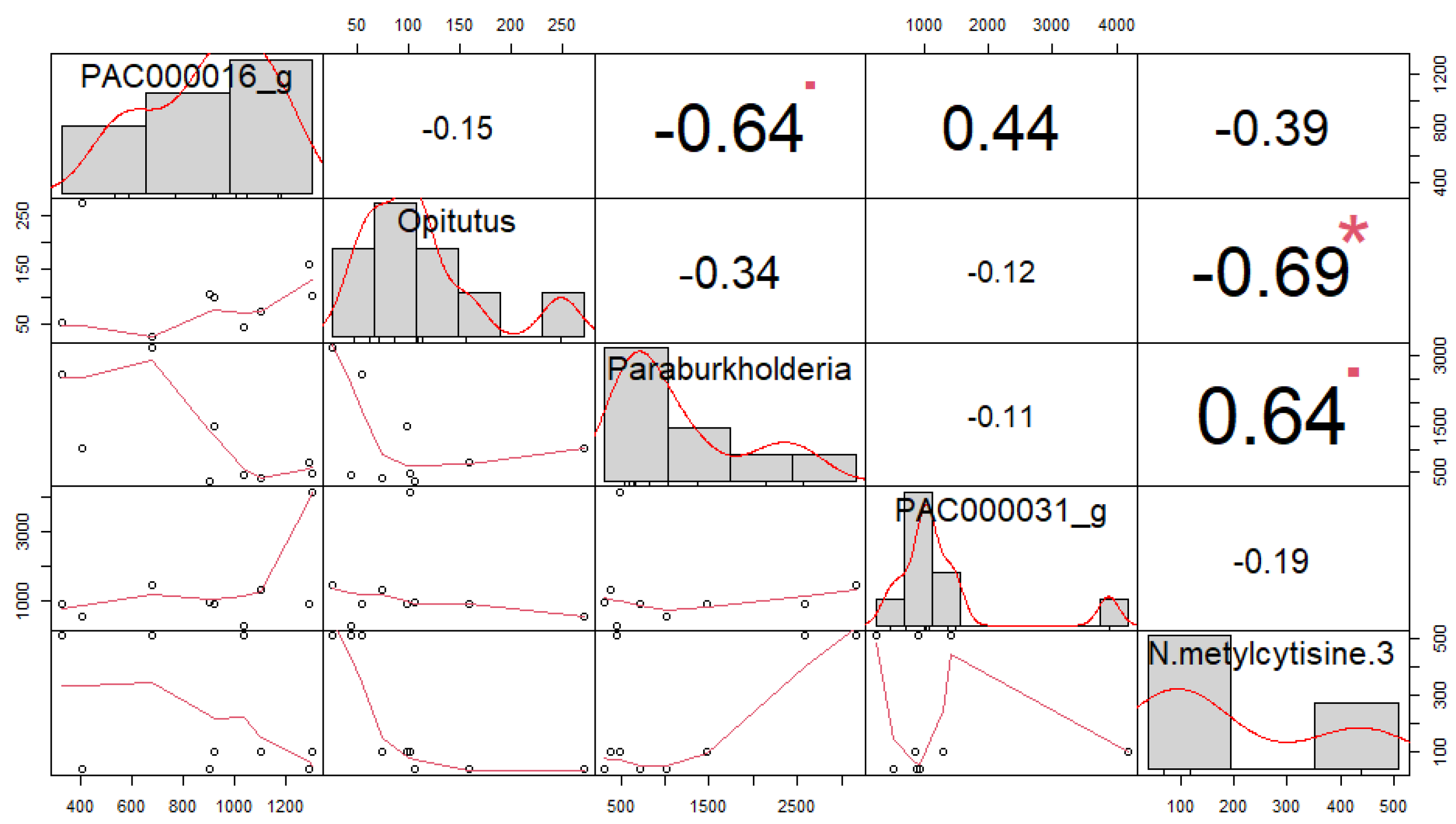
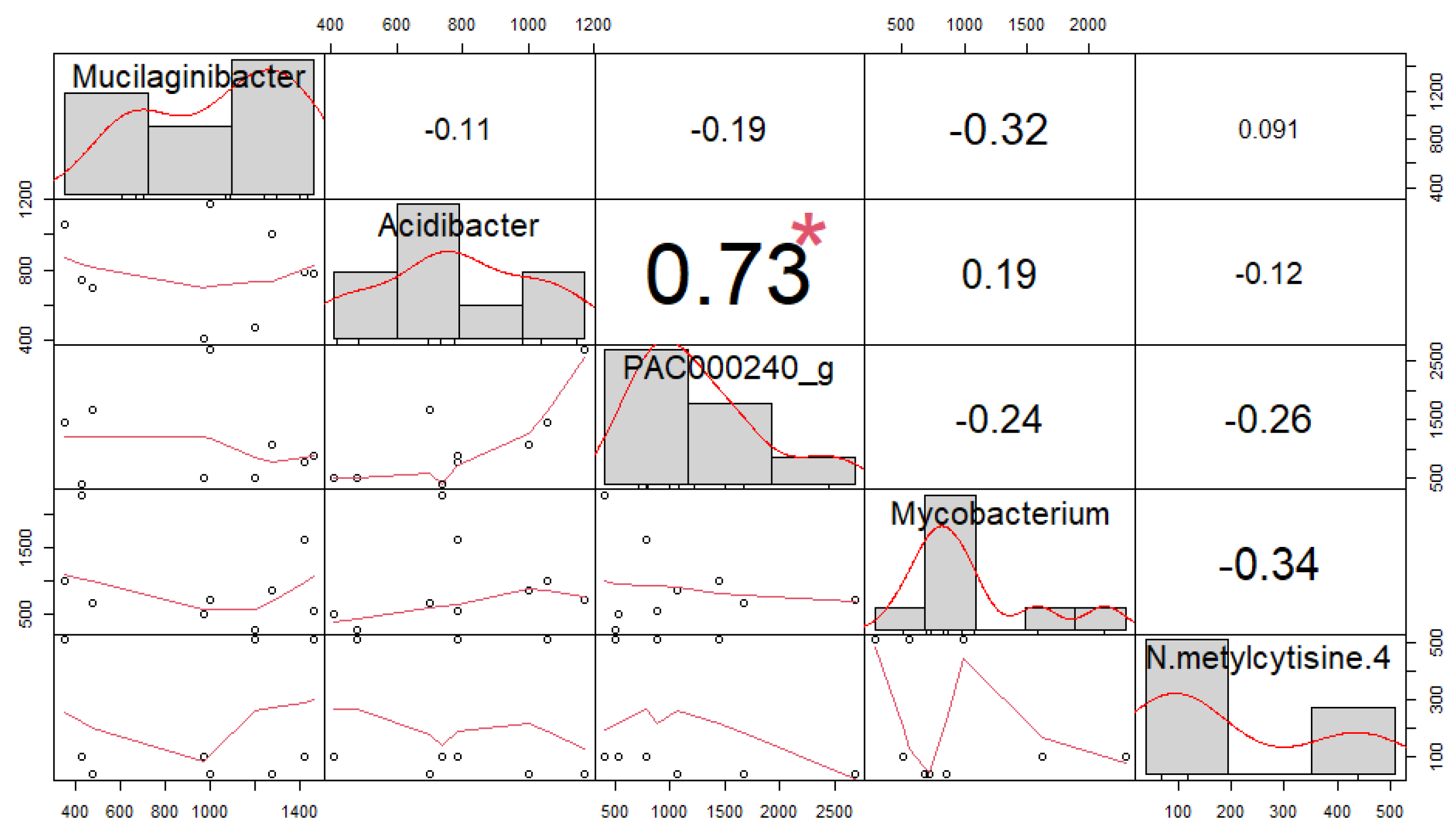
3.4. Correlation among Soil Microorganisms and Secondary Compounds
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, W.T. A discussion on Korean endemic genera plants. Korean J. Plant Taxon. 1969, 1, 15–21. [Google Scholar]
- Nakai, T. Notulae ad plantas Japoniae et Koreae XXI. Shokubutsugaku Zasshi 1919, 33, 193–216. [Google Scholar] [CrossRef]
- Nakai, T. Genera nova Rhamnacearum et Leguminosarum ex Asia orientali. Shokubutsugaku Zasshi 1923, 37, en29–en34. [Google Scholar] [CrossRef]
- Yoo, B.H.; Ku, Y.B.; Oh, H.K.; Suh, M.H.; Shim, K.Y.; Yeon, M.H.; Lee, M.H. The Conservation Strategy for the Endangered and Reserved Plants Based on the Ecological and Genetic characteristics (V); National Institute of Environmental Research: Incheon, Korea, 2005.
- Jang, J.S.; Kim, H.; Jeon, J.I. The New Dendrology; Hyangmunsa: Seoul, Korea, 2019; pp. 236–237. [Google Scholar]
- Cheon, K.; Jang, S.; Lee, W.; Yoo, K. The natural habitat and distribution of Echinosophora koreensis (Nakai) Nakai in Korea. Korean J. Plant Taxon. 2009, 39, 254–263. [Google Scholar] [CrossRef]
- Kang, K.S. Find Echinosophora koreensis NAKAI: Yellow flowers adorn the barren land. Amat. Bot. Club Korea 1992, 24, 274–275. [Google Scholar]
- Murakoshi, I.; Fukuchi, K.; Haginiwa, J.; Ohmiya, S.; Otomasu, H. N-(3-Oxobutyl) cytisine: A new lupin alkaloid from Echinosophora koreensis. Phytochemistry 1977, 16, 1460–1461. [Google Scholar] [CrossRef]
- Murakoshi, I.; Kidoguchi, E.; Kubota, M.; Haginiwa, J.; Ohmiya, S.; Otomasu, H. Lupin alkaloids from Echinosophora koreensis. Phytochemistry 1982, 21, 2385–2388. [Google Scholar] [CrossRef]
- Kim, C.M.; Kang, S.S. Studies on the constituents of the stems of Echinosophora koreensis. Yakhak Hoeji 1986, 30, 139–142. [Google Scholar]
- Murakoshi, I.; Watanabe, M.; Okuda, T.; Kidoguchi, E.; Haginiwa, J.; Ohmiya, S.; Otomasu, H. Lupin alkaloids from flowers of Echinosophora koreensis. Phytochemistry 1985, 24, 2707–2708. [Google Scholar] [CrossRef]
- Kim, C.; Lee, K. A study on chemical constituents and biological activity of Echinosophora koreensis Nakai. Korean J. Pharmacogn. 1990, 21, 137–141. [Google Scholar]
- Kim, J.; Byun, J.; Son, K.; Kim, H.; Chang, H.; Kang, S. Isolation of flavonoids and a saponin from Echinosophora koreensis. Korean J. Pharmacogn. 2002, 33, 110–115. [Google Scholar]
- Choi, E.J.; Kwon, H.C.; Sohn, Y.C.; Nam, C.W.; Park, H.B.; Kim, C.Y.; Yang, H.O. Four flavonoids from Echinosophora koreensis and their effects on alcohol metabolizing enzymes. Arch. Pharm. Res. Jpn. 2009, 32, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.H.; Kang, H.K.; Hosseindoust, A.; Mun, J.Y.; Moturi, J.; Tajudeen, H.; Lee, H.; Cheong, E.J.; Kim, J.S. Effects of Scopoletin Supplementation and Stocking Density on Growth Performance, Antioxidant Activity, and Meat Quality of Korean Native Broiler Chickens. Foods 2021, 10, 1505. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Nakashima, N.; Kimura, I.; Kimura, M. Hypoglycemic activity and mechanisms of extracts from mulberry leaves (folium mori) and cortex mori radicis in streptozotocin-induced diabetic mice. Yakugaku Zasshi J. Pharm. Soc. Jpn. 1995, 115, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.; Chen, F.; Nakashima, N.; Kimura, I.; Asano, N.; Koya, S. Antihyperglycemic effects of N-containing sugars derived from mulberry leaves in streptozocin-induced diabetic mice. J. Trad. Med. 1995, 12, 214–219. [Google Scholar]
- Cha, J.Y.; Kim, H.J.; Chung, C.H.; Cho, Y.S. Antioxidative activities and contents of polyphenolic compound of Cudrania tricuspidata. J. Korean Soc. Food Sci. Nutr. 1999, 28, 1310–1315. [Google Scholar]
- Zlatić, N.M.; Stanković, M.S. Variability of secondary metabolites of the species Cichorium intybus L. from different habitats. Plants 2017, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Eo, H.J.; Kang, Y.; Kim, D.S.; Park, Y.; Kim, H.J.; Park, G.H. Analysis of the correlation between marker compounds contents and cultivation environment of Cornus officinalis. J. Appl. Biol. Chem. 2020, 63, 175–180. [Google Scholar] [CrossRef]
- Jeong, D.H.; Kim, K.Y.; Park, H.W.; Jung, C.R.; Kim, H.J.; Jeon, K.S. Growth characteristics of Ligusticum chuanxing Hort. according to soil and meteorological environment by each cultivation. Korean J. Plant Resour. 2021, 34, 64–72. [Google Scholar]
- Kim, K.; Huh, J.; Um, Y.; Jeon, K.S.; Kim, H. The comparative of growth characteristics and ginsenoside contents in wild-simulated ginseng (Panax ginseng CA Meyer) on different years by soil properties of cultivation regions. Korean J. Plant Resour. 2020, 33, 651–658. [Google Scholar]
- Jeong, S.J.; Oh, J.S.; Seok, W.Y.; Cho, M.Y.; Seo, J.B. The Effect of Chitosan and Wood Vinegar Treatment on the Growth of Eggplant and Leaf Lettuce. Korean J. Org. Agric. 2007, 15, 437–452. [Google Scholar]
- Koo, H.B.; Hwang, H.; Han, J.Y.; Cheong, E.J.; Kwon, Y.; Choi, Y.E. Enhanced production of pinosylvin stilbene with aging of Pinus strobus callus and nematicidal activity of callus extracts against pinewood nematodes. Sci. Rep. 2022, 12, 770. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic. Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heck Jr, K.L.; van Belle, G.; Simberloff, D. Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 1975, 56, 1459–1461. [Google Scholar] [CrossRef]
- Rohlf, F.J. Shape statistics: Procrustes superimpositions and tangent spaces. J. Classif. 1999, 16, 197–223. [Google Scholar] [CrossRef]
- Schmeller, T.; Sauerwein, M.; Sporer, F.; Wink, M.; Müller, W.E. Binding of quinolizidine alkaloids to nicotinic and muscarinic acetylcholine receptors. J. Nat. Prod. 1994, 57, 1316–1319. [Google Scholar] [CrossRef] [PubMed]
- Petruczynik, A.; Wróblewski, K.; Misiurek, J.; Plech, T.; Szalast, K.; Wojtanowski, K.; Mroczek, T.; Szymczak, G.; Waksmundzka-Hajnos, M.; Tutka, P. Determination of Cytisine and N-Methylcytisine from Selected Plant Extracts by High-Performance Liquid Chromatography and Comparison of Their Cytotoxic Activity. Toxins 2020, 12, 557. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.C.; Chen, K.K. The pharmacological action of N-methylcytisine. J. Pharmacol. Exp. Ther. 1943, 79, 334–339. [Google Scholar]
- Eo, H.J.; Park, Y.; Park, G.H.; Kim, J.; Kim, D.S.; Kang, Y.; Kim, K.; Jang, J.H.; Kim, H. Study on the Correlation between the Soil Properties and Albiflorin, Paeoniflorin Contents of Paeonia lactiflora Pall. Korean J. Plant Resour. 2021, 34, 384–394. [Google Scholar]
- Cheon, K.S. Distribution, Natural Habitat Characteristics and Genetic Diversity of Echinosophora koreensis (Nakai) Nakai. Master’s Thesis, Univ. of Kangwon, Chuncheon, Korea, 2010. [Google Scholar]
- Ryffel, F.; Helfrich, E.J.; Kiefer, P.; Peyriga, L.; Portais, J.; Piel, J.; Vorholt, J.A. Metabolic footprint of epiphytic bacteria on Arabidopsis thaliana leaves. ISME J. 2016, 10, 632–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef] [PubMed]
- Pilatti, D.M.; Fortes, A.; Jorge, T.; Boiago, N.P. Comparison of the phytochemical profiles of five native plant species in two different forest formations. Braz. J. Biol. 2018, 79, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanInsberghe, D.; Maas, K.R.; Cardenas, E.; Strachan, C.R.; Hallam, S.J.; Mohn, W.W. Non-symbiotic Bradyrhizobium ecotypes dominate North American forest soils. ISME J. 2015, 9, 2435–2441. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.; Subramaniam, S. Bradyrhizobium japonicum USDA110: A representative model organism for studying the impact of pollutants on soil microbiota. Sci. Total Environ. 2018, 624, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Reichman, S.M. Probing the plant growth-promoting and heavy metal tolerance characteristics of Bradyrhizobium japonicum CB1809. Eur. J. Soil Biol. 2014, 63, 7–13. [Google Scholar] [CrossRef]
- Zawoznik, M.S.; Tomaro, M.L. Effect of chlorimuron-ethyl on Bradyrhizobium japonicum and its symbiosis with soybean. Pest. Manag. Sci. 2005, 61, 1003–1008. [Google Scholar] [CrossRef]
- Soria, M.A.; Pagliero, F.E.; Correa, O.S.; Kerber, N.L.; Garcia, A.F. Tolerance of Bradyrhizobium japonicum E109 to osmotic stress and the stability of liquid inoculants depend on growth phase. World J. Microbiol. Biotechnol. 2006, 22, 1235–1241. [Google Scholar] [CrossRef]
- Embleton, L. Silver Nanoparticles and the Plant-Associating Abilities of Rhizobiaceae Bacteria. Master’s Thesis, Queen’s University, Kingston, ON, Canada, 2016. [Google Scholar]
- Van Passel, M.W.; Kant, R.; Palva, A.; Copeland, A.; Lucas, S.; Lapidus, A.; Glavina del Rio, T.; Pitluck, S.; Goltsman, E.; Clum, A. Genome sequence of the verrucomicrobium Opitutus terrae PB90-1, an abundant inhabitant of rice paddy soil ecosystems. J. Bacteriol. 2011, 193, 2367–2368. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Ladau, J.; Clemente, J.C.; Leff, J.W.; Owens, S.M.; Pollard, K.S.; Knight, R.; Gilbert, J.A.; McCulley, R.L. Reconstructing the microbial diversity and function of pre-agricultural tallgrass prairie soils in the United States. Science 2013, 342, 621–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.B. Conservation Strategies and Vegetation Characteristics of Echinosophora koreensis of Korean Endemic Plants. Master’s Thesis, Univ. of Kangwon, Chuncheon, Korea, 2019. [Google Scholar]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A.S. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Männistö, M.K.; Tiirola, M.; Häggblom, M.M. Bacterial communities in Arctic fjelds of Finnish Lapland are stable but highly pH-dependent. FEMS Microbiol. Ecol. 2007, 59, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Jeon, J.; Lee, S.; Kim, H.; Ahn, T.; Song, H. Plant growth promotion in soil by some inoculated microorganisms. J. Microbiol. 2003, 41, 271–276. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5 > 3) |
|---|---|
| 16S v34_F | TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG |
| TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGN*CCTACGGGNGGCWGCAG | |
| TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGNNCCTACGGGNGGCWGCAG | |
| TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGNNNCCTACGGGNGGCWGCAG | |
| 16S v34_R | GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC |
| GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGNGACTACHVGGGTATCTAATCC | |
| GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGNNGACTACHVGGGTATCTAATCC | |
| GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGNNNGACTACHVGGGTATCTAATCC |
| Area | Target Reads | OTUs | ACE | CHAO | Shannon | Phylogenetic Diversity |
|---|---|---|---|---|---|---|
| C1 | 60,046 | 2167 | 2496.92 | 2375.37 | 5.56 | 2554 |
| C2 | 42,686 | 2626 | 3095.61 | 2936.56 | 6.05 | 3099 |
| C3 | 30,755 | 2454 | 2862.87 | 2755.94 | 6.35 | 2834 |
| 44,495.67 ± 8503.86 a* | 2415.67 ± 133.88 a | 2818.47 ± 174.25 a | 2689.29 ± 165.39 a | 5.99 ± 0.23 a | 2829.00 ± 157.35 a | |
| G1 | 85,014 | 4727 | 4980.30 | 4851.04 | 6.76 | 4818 |
| G2 | 92,879 | 4590 | 4802.33 | 4695.96 | 6.64 | 4635 |
| G3 | 93,790 | 4254 | 4353.61 | 4303.22 | 6.59 | 4337 |
| 90,561.00 ± 2785.94 b | 4523.67 ± 140.51 c | 4712.08 ± 186.45 c | 4616.74 ± 163.03 c | 6.66 ± 0.05 b | 4596.67 ± 140.17 b | |
| I1 | 49,775 | 3473 | 3819.37 | 3682.19 | 6.53 | 3559 |
| I2 | 34,407 | 2522 | 2831.94 | 2707.12 | 6.35 | 2769 |
| I3 | 66,734 | 3716 | 4011.67 | 3857.06 | 6.45 | 3643 |
| 50,305.33 ± 9335.77 a | 3237.00 ± 364.32 ab | 3554.33 ± 365.43 ab | 3415.46 ± 357.75 ab | 6.44 ± 0.05 ab | 3323.67 ± 279.39 a | |
| Y1 | 74,273 | 3658 | 3961.13 | 3821.48 | 6.19 | 3445 |
| Y2 | 64,172 | 3742 | 4009.72 | 3877.06 | 6.30 | 3786 |
| Y3 | 56,853 | 3193 | 3552.73 | 3404.81 | 6.08 | 3250 |
| 65,099.33 ± 5050.05 ab | 3531.00 ± 170.73 b | 3841.19 ± 144.91 bc | 3701.12 ± 149.02 b | 6.19 ± 0.06 ab | 3693.67 ± 156.63 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Cheong, E.J. Correlation between the Concentration of Secondary Metabolites and Soil Microorganisms in Sophora Koreensis Nakai from Different Habitat. Forests 2022, 13, 1079. https://doi.org/10.3390/f13071079
Lee H, Cheong EJ. Correlation between the Concentration of Secondary Metabolites and Soil Microorganisms in Sophora Koreensis Nakai from Different Habitat. Forests. 2022; 13(7):1079. https://doi.org/10.3390/f13071079
Chicago/Turabian StyleLee, Hwa, and Eun Ju Cheong. 2022. "Correlation between the Concentration of Secondary Metabolites and Soil Microorganisms in Sophora Koreensis Nakai from Different Habitat" Forests 13, no. 7: 1079. https://doi.org/10.3390/f13071079