
Ecological Dynamics and Regeneration Expansion of Treeline Ecotones in Response to Climate Change in Northern Bhutan Himalayas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Field Methods, Study Sites and Materials
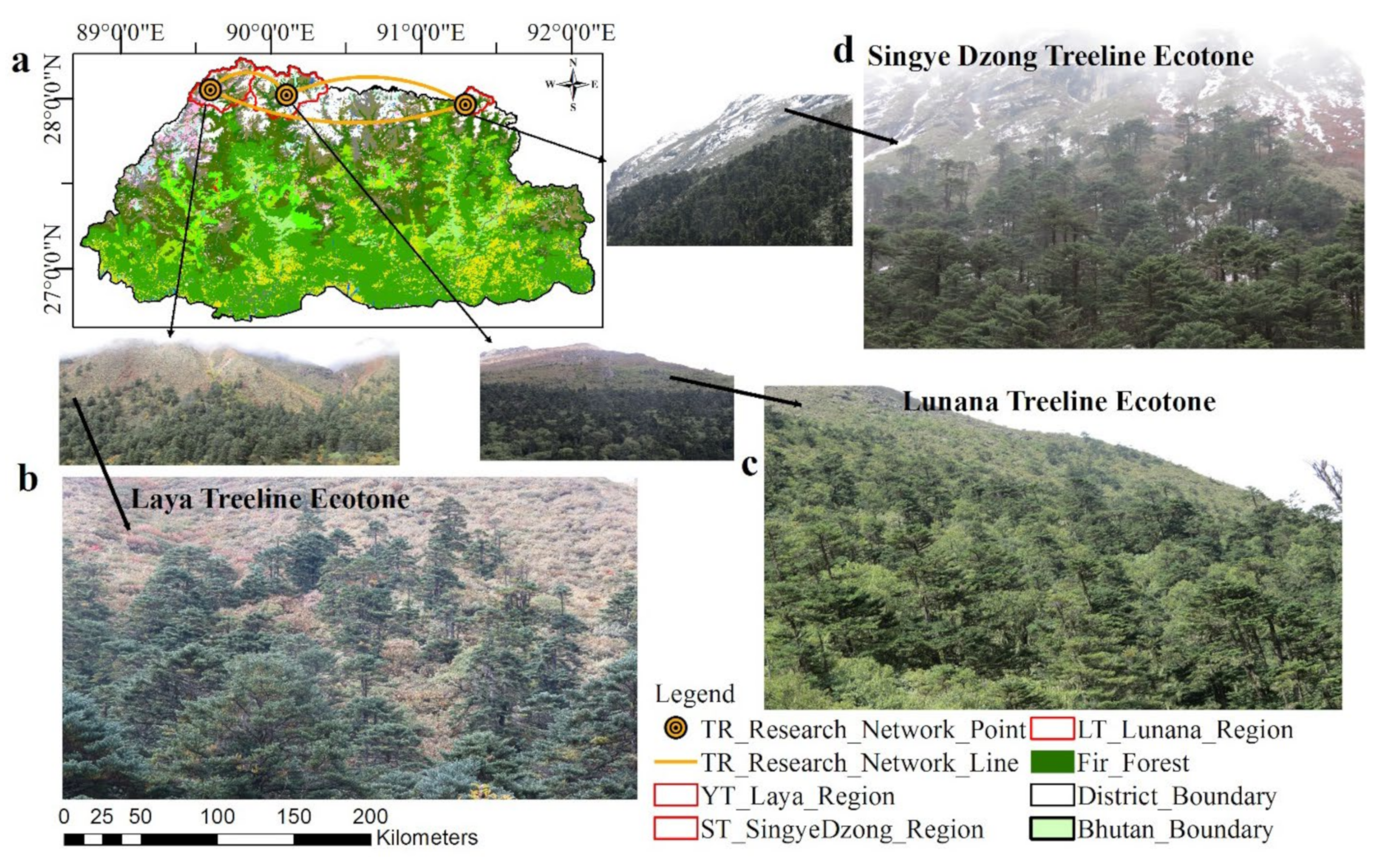
2.1.1. Site Selection
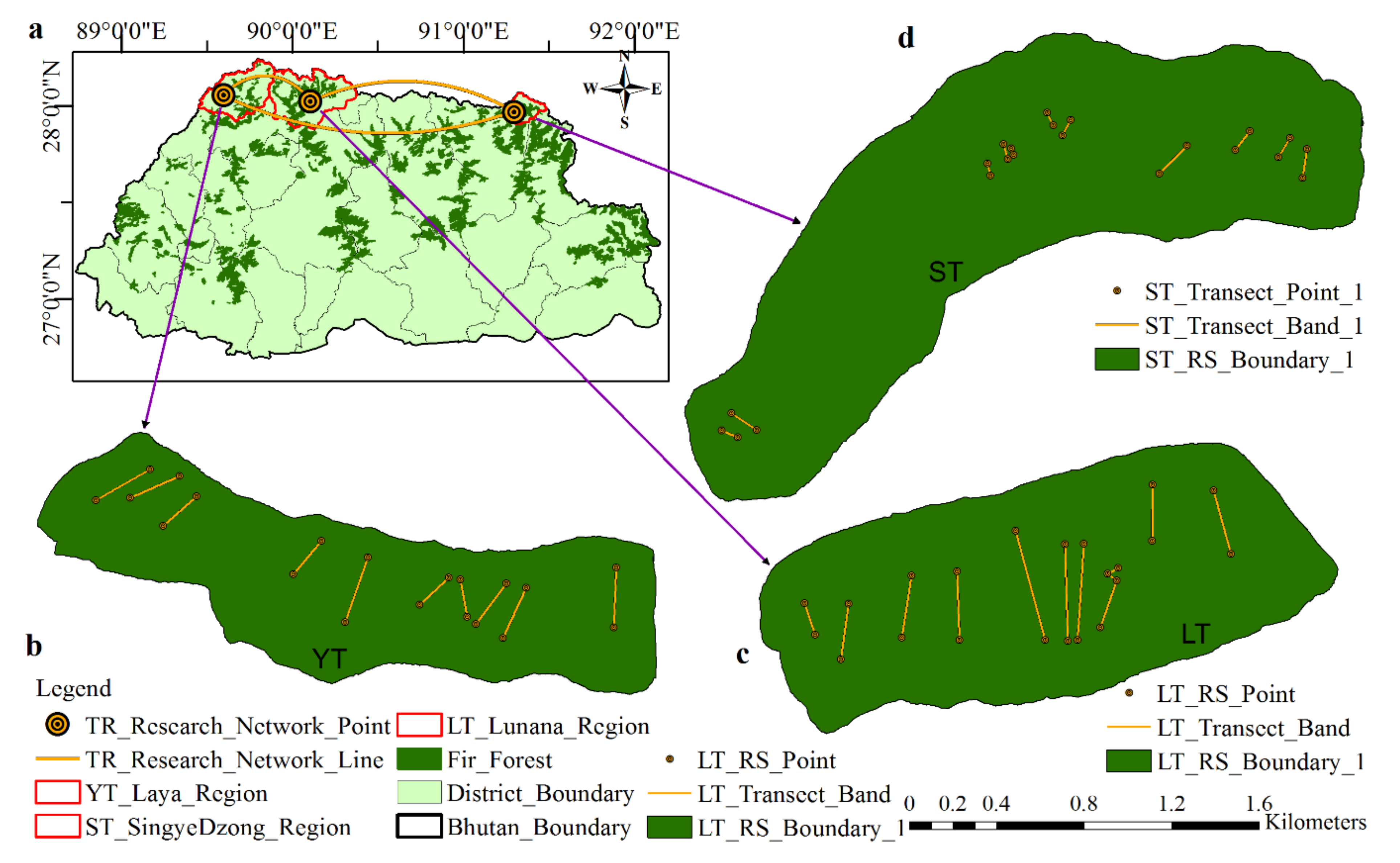
2.1.2. Sampling Transects-Enumeration, Measurement and Coring of Trees
2.2. Laboratory Sanding Method, Cross-Dating Analysis, Tree Age Estimation Techniques
2.3. Climate Data—Grid CRU TS 4.01 and Tree-Ring Based Reconstructed Temperature
2.4. Tree Demographic Size, Age and Recruitment Frequency Tempo-Spatial Distribution in Relation to Elevation and Climate
2.5. Dendroecological Analysis—Tree Regeneration, Delimiting Distribution and Upward Expansion of Treeline Ecotones Driven under Changes in Climate and Elevations
2.6. GIS Analysis—Spatial Interpolation and Modelling of Climate and Elevation Delimiting the Distribution of Fir Forests
3. Results
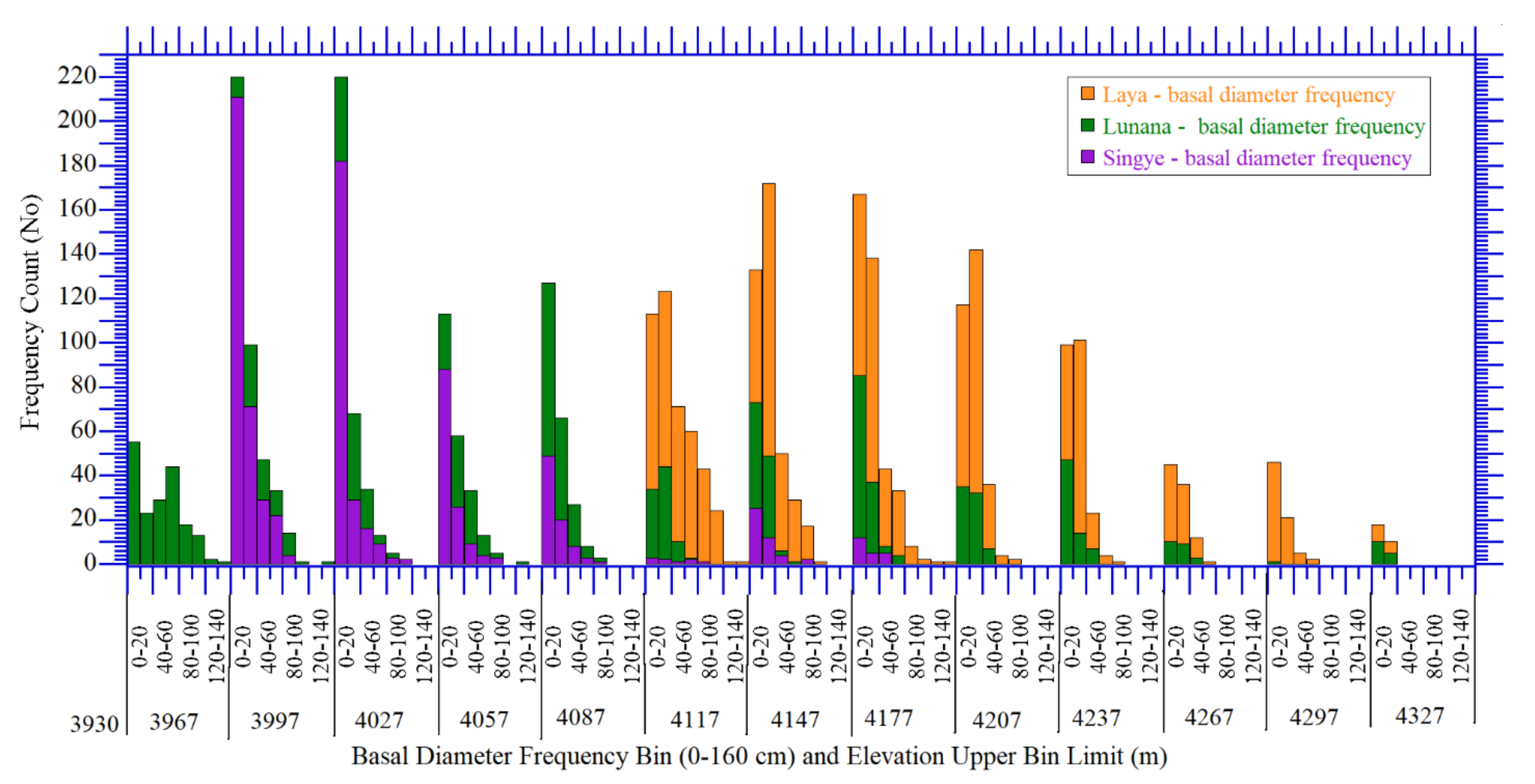
3.1. Dendrochronologically Estimated Tree Ages Based on Basal Diameters and Tree-Rings
3.2. Tree Demographic Population Size and Age Class Frequency in Relation to Tempo-Spatial Distribution
3.3. Tree Demographic Size and Recruitment Frequency in Relation to Tempo-Spatial Distribution
3.3.1. Tree Basal Diameter and Recruitment Frequency Distribution in Response to Elevation Gradients
3.3.2. Tree Size and Age Distribution as Site Specific Dynamic Response of Treeline Ecotones
3.4. Tree Regeneration and Growth Dynamics in Response to Climate Change
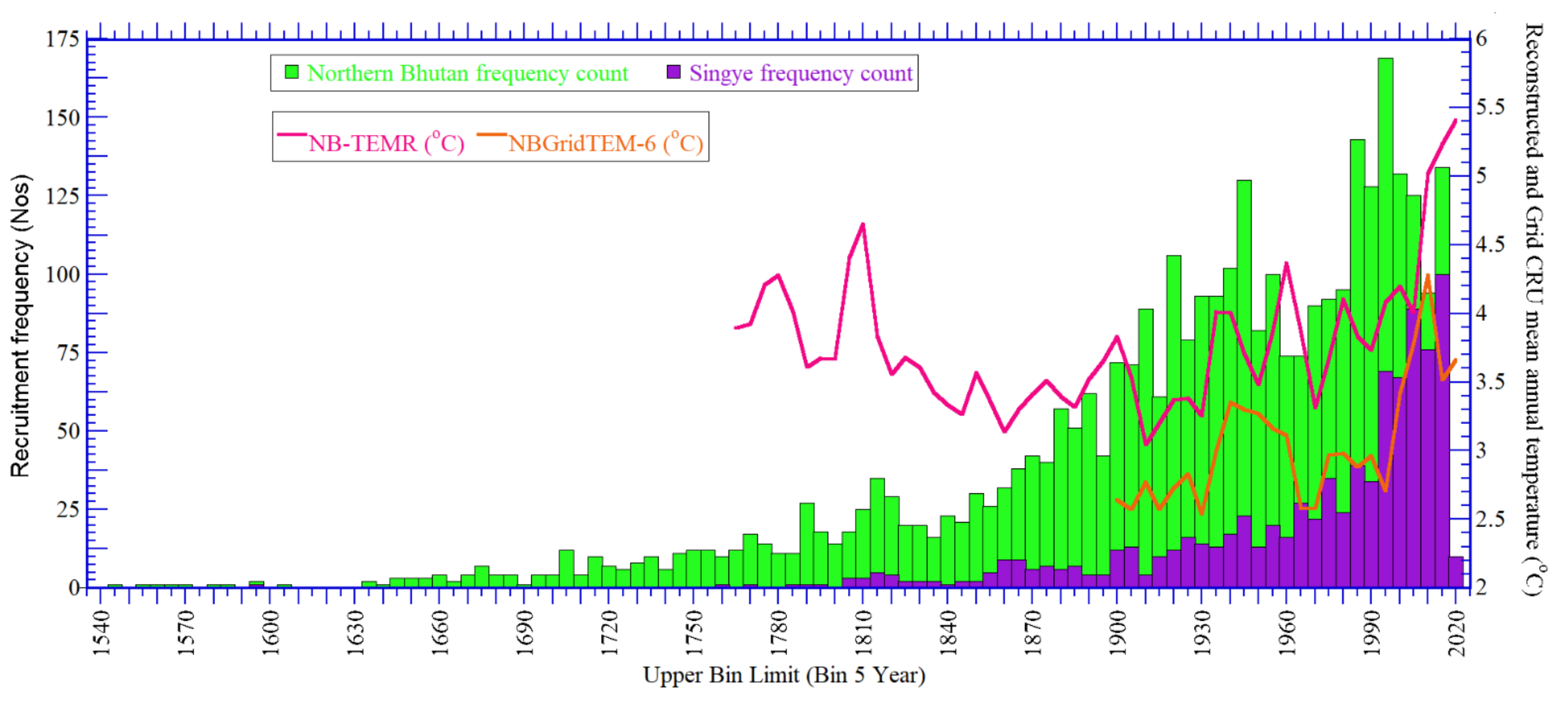
3.4.1. Tree Recruitment Frequency Distribution in Relation to Climate Change
3.4.2. Tree Recruitment Year Distribution in Relation to Climate Change and Elevation
3.4.3. Tree Regeneration and Age Distribution in the Upward Expansion of Treeline Ecotones Driven under Climate Change
- Upward regeneration expansion in Lunana treeline region
- b.
- Upward regeneration expansion in Laya treeline region
- c.
- Upward regeneration expansion in Singye treeline region
3.5. Geographical Position of Vegetation Margins in Response to Changes in Climate and Elevation

4. Discussion
4.1. Tree Demographic Size, Age and Population Distribution Triggered by Changes in Elevation
4.2. Tree Regeneration and Recruitment Dynamics Driven by Warming Mean Annual Temperatures
4.3. Regeneration Dynamics in Upward Expansion of Treeline Ecotones Driven by Climate Change
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hiller, B.; Muterthies, A. Humus forms and reforestation of an abandoned pasture at the alpine timberline (upper Engadine, Central Alps, Switzerland). In Mountain Ecosystems: Studies in Treeline Ecology; Broll, G., Keplin, B., Eds.; Springer Science+Business Media: Berlin, Germany, 2005; pp. 203–218. [Google Scholar]
- LaMarche, V.C. Paleoclimatic inferences from long tree-ring records: Intersite comparison shows climatic anomalies that may be linked to features of the general circulation. Science 1974, 183, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- LaMarche, V.C.J.; Mooney, H.A. Altithermal timberline advance in western United States. Nature 1967, 213, 980–982. [Google Scholar] [CrossRef]
- LaMarche, V.C., Jr.; Stockton, C.W. Chronologies from temperature -sensitive bristlecone pines at upper treeline in Western United States. Tree-Ring Bull. 1974, 4, 21–45. [Google Scholar]
- Holtmeier, F.-K.; Broll, G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Glob. Ecol. Biogeogr. 2005, 14, 395–410. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; Hansen-Speer, K., Ed.; University of Arizona Press: Tucson, AZ, USA, 2010; p. 521. [Google Scholar]
- Harsch, M.A. Treeline Dynamics: Pattern and Process At Multiple Spatial Scales; Research Lincoln University: Lincoln, New Zealand, 2010. [Google Scholar]
- Crawford, R.M.M. Peripheral Plant Population Survival in Polar Regions. In Mountain Ecosystems: Studies in Treeline Ecology; Broll, G., Keplin, B., Eds.; Springer Science, Business Media: Berlin, Germany, 2005; pp. 43–76. [Google Scholar]
- Tiwari, A.; Fan, Z.-X.; Jump, A.S.; Li, S.-F.; Zhou, Z.-K. Gradual expansion of moisture sensitive Abies spectabilis forest in the Trans-Himalayan zone of central Nepal associated with climate change. Dendrochronologia 2016, 41, 34–43. [Google Scholar] [CrossRef]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Carrer, M. Site- and species-specific treeline responses to climatic variability in eastern Nepal Himalaya. Dendrochronologia 2016, 13, 1. [Google Scholar] [CrossRef]
- Wilson, R.; Wiles, G.; D’Arrigo, R.; Zweck, C. Cycles and shifts: 1300 years of multi-decadal temperature variability in the Gulf of Alaska. Clim. Dynam. 2007, 28, 425–440. [Google Scholar] [CrossRef]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Borgaonkar, H.P. Treeline dynamics with climate change at the Central Nepal Himalaya. Clim. Past 2014, 10, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Sveinbjörnsson, B. North American and European Treelines: External Forces and Internal Processes Controlling Position. J. Human Environ. 2000, 29, 388–395. [Google Scholar] [CrossRef]
- Keenan, T.F.; Riley, W.J. Greening of the land surface in the world’s cold regions consistent with recent warming. Nat. Clim. Chang. 2018, 8, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, Q.B.; Ma, K.P. Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China. Glob. Ecol. Biogeogr. 2006, 15, 406–415. [Google Scholar] [CrossRef]
- Schickhoff, U. The upper timberline in the Himalayas, Hindu Kush and Karakorum: A review of geographical and ecological aspects. In Mountain Ecosystems: Studies in Treeline Ecology; Broll, G., Keplin, B., Eds.; Springer Science+Business Media: Berlin, Germany, 2005; pp. 275–354. [Google Scholar]
- Gratzer, G.; Rai, P.B.; Glatzel, G. The influence of the bamboo Yushania microphylla on regeneration of Abies densa in central Bhutan. Can. J. Forest Res. 1999, 29, 1518–1527. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chaudhary, V. Late-summer temperature reconstruction of the eastern Himalayan region based on tree-ring data of Abies densa. Tree Ring Res. 2003, 35, 196–202. [Google Scholar] [CrossRef]
- Edward, R.C.; Krusic, P.J.; Jones, P.D. Dendroclimatic Signals in Long Tree-Ring Chronologies from the Himalayas of Nepal. Int. J. Climatol. 2003, 23, 707–732. [Google Scholar] [CrossRef]
- Dawadi, B.; Liang, E.; Tian, L.; Devkota, L.P.; Yao, T. Pre-monsoon precipitation signal in tree rings of timberline Betula utilis in the central Himalayas. Quat. Int. 2013, 283, 72–77. [Google Scholar] [CrossRef]
- Tse-ring, K.; Sharma, E.; Chettri, N.; Shrestha, A. Climate Change Vulnerability of Mountain Ecosystems in the Eastern Himalayas; Climate Change Impact and Vulnerability in the Eastern Himalayas–Synthesis Report; International Centre for Integrated Mountain Development: Kathmandu, Nepal, 2010; p. 116. [Google Scholar]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T.; et al. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dynam. 2015, 6, 245–265. [Google Scholar] [CrossRef]
- Pollmann, W.; Hildebrand, R. Structure and composition of species in timberline ecotones of the southern Andes. In Mountain Ecosystems: Studies in Treeline Ecology; Broll, G., Keplin, B., Eds.; Springer Science+Business Media: Berlin, Germany, 2005; pp. 117–152. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press Inc. (London) Ltd.: New York, NY, USA, 1976; p. 571. [Google Scholar]
- Ahmed, M. The Science of Tree Rings: Dendrochronology; Qureshi Art Press: Karachi, Pakistan, 2014; p. 301. [Google Scholar]
- Camarero, J.J.; Gutiérrez, E. Pace and Pattern of recent Treeline Dynamics: Responseof ecotones to Climatic Variability in the Spanish Pyrenees. Clim. Chang. 2004, 63, 181–200. [Google Scholar] [CrossRef]
- Vittoz, P.; Rulence, B.; Largey, T.; Freléchoux, F. Effects of Climate and Land-Use Change on the Establishment and Growth of Cembran Pine (Pinus cembra L.) over the Altitudinal Treeline Ecotone in the Central Swiss Alps. Arctic Antarcticand Alpine Res. 2008, 40, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Gratzer, G.; Rai, P.B.; Schieler, K. Structure and regeneration dynamics of Abies densa forests in central Bhutan. Aust. J. Forest Sci. 2002, 3, 279–288. [Google Scholar]
- Eguchi, T. Regional and seasonal change of precipitation in Bhutan. In Life Zone Ecology of the Bhutan Himalayas; Ohsawa, M., Ed.; Laboratory of the Chiba University: Edinburgh, UK, 1991; pp. 1–20. [Google Scholar]
- Grierson, A.C.J.; Long, D.G. Flora of Bhutan; Royal Botanical Garden of Edinburgh: Edinburgh, UK, 1983. [Google Scholar]
- Krusic, P.J.; Cook, E.R.; Dukpa, D.; Putnam, A.E.; Rupper, S.; Schaefer, J. Six hundred thirty-eight years of summer temperature variability over the Bhutanese Himalaya. Geophys. Res. Lett. 2015, 42, 2988–2994. [Google Scholar] [CrossRef]
- Khandu, Y. Regeneration and Expansion of Native Pinewoods of Birse Forest in Scotland; Department of Plant and Soil Science, College of Life Science and Medicine, University of Aberdeen: Aberdeen, UK, 2007. [Google Scholar]
- Stokes, M.; Smiley, T. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Chen, F.; Wang, H.; Chen, F.-H.; Yuan, Y.; Zhang, R.-B. Tree-ring reconstruction of July–May precipitation (AD 1816–2010) in the northwestern marginal zone of the East Asian summer monsoon reveals the monsoon-related climate signals. Int. J. Climatol. 2015, 35, 2109–2121. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in Environmental Science; Cook, E.R., Kairiukstis, L.A., Eds.; International Institute for Applied System Analysis, Kluwer Academic Publisher: Dordrecht, The Netherlands, 1992; p. 394. [Google Scholar]
- Chen, F.; Shang, H.; Panyushkina, I.P.; Meko, D.M.; Yu, S.; Yuan, Y.; Chen, F. Tree-ring reconstruction of Lhasa River streamflow reveals 472 years of hydrologic change on southern Tibetan Plateau. J. Hydrol. 2019, 572, 169–178. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings: Basic and Application of Dendrochronology; D. Reidel Publishing Company: Dordrecht, The Netherland, 1988; p. 276. [Google Scholar]
- Prasad Gaire, N.; Fan, Z.-X.; Bräuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Gaire, N.P.; Bhuju, D.R.; Koirala, M.; Shah, S.K.; Carrer, M.; Timilsena, R. Tree-ring based spring precipitation reconstruction in western Nepal Himalaya since AD 1840. Dendrochronologia 2017, 42, 21–30. [Google Scholar] [CrossRef]
- Orvis, K.H.; Grissino-Mayer, H.D. Standardizing the reporting of abrasive papers used to surface tree-ring samples. Tree-Ring Res. 2002, 58, 47–50. [Google Scholar]
- Cook, E.R.; Kairiukstis, L. Methods of Dendrochronology; Kluwer: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bullet. 1983, 43, 69–78. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating cross-dating accuracy: A manual and turtorial for the computer program COFECHA. Tree Ring Res. 2001, 57, 205–221. [Google Scholar]
- Duncan, R.P. AN evaluation of errors in tree age estimates based on increment cores in kahikatea (Dacrycarpus dacrydioides). N. Z. Nat. Sci. 1989, 16, 31–37. [Google Scholar]
- Rozas, V. Tree age estimates in Fagus sylvatica and Quercus robur: Testing previous and improved methods. Plant Ecol. 2003, 167, 193–212. [Google Scholar] [CrossRef]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft: New York, NY, USA, 2020. [Google Scholar]
- Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations–the CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- British Atmospheric Data Centre. Climatic Research Unit (CRU) Time-Series (TS) Version 4.01 of High-Resolution Gridded Data of Month-By-Month Variation in Climate (January 1901–December 2016). Natural Environment Research Council (NERC), London, UK. 2016. Available online: http://badc.nerc.ac.uk (accessed on 23 February 2022).
- Khandu, Y.; Polthanee, A.; Aayutthaya, S.I.N. Dendroclimatic reconstruction of mean annual temperature over treeline regions of Northern Bhutan Himalaya. Dendrochronologia 2022. Submitted, under review. [Google Scholar]
- Synergy, S. Kaleida Graph Manual (Window); Synergy Software: Reading, PA, USA, 2020; p. 340. [Google Scholar]
- IBM Corporation. IBM SPSS Statistics for Windows, Version 23; IBM Corporation: Armonk, NY, USA, 2015. [Google Scholar]
- ESRI (Ed.) ArcGIS 10.3.1 for Desktop; Environmental Systems Research Institute, Inc.: Redlands, CA, USA, 2015; Available online: http://www.esri.com/ (accessed on 23 February 2022).
- Zhang, R.-B.; Yuan, Y.-J.; Wei, W.-S.; Gou, X.-H.; Yu, S.-L.; Shang, H.-M.; Chen, F.; Zhang, T.-W.; Qin, L. Dendroclimatic reconstruction of autumn–winter mean minimum temperature in the eastern Tibetan Plateau since 1600 AD. Dendrochronologia 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Recent treeline dynamics are similar between dry and mesic areas of Nepal, central Himalaya. J. Plant Ecol. 2014, 8, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.D.; Miller, R.F. Structure and development of expanding western juniper woodlands as influenced by two topographic variables. Forest Ecol. Manag. 2006, 229, 7–15. [Google Scholar] [CrossRef]
- Moen, J.; Cairns, D.M.; Lafon, C.W. Factors structuring the treeline ecotone in Fennoscandia. Plant Ecol. Divers. 2008, 1, 77–87. [Google Scholar] [CrossRef]
- Mainali, J.; All, J.; Jha, P.K.; Bhuju, D.R. Responses of Montane Forest to Climate Variability in the Central Himalayas of Nepal. Mt. Res. Dev. 2015, 35, 66–77. [Google Scholar] [CrossRef]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and Ecosystem Responses to Elevational Gradients: Processes, Mechanisms, and Insights for Global Change. Ann. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Mayor, J.R.; Sanders, N.J.; Classen, A.T.; Bardgett, R.D.; Clément, J.-C.; Fajardo, A.; Lavorel, S.; Sundqvist, M.K.; Bahn, M.; Chisholm, C.; et al. Elevation alters ecosystem properties across temperate treelines globally. Nature 2017, 542, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgaonkar, H.P.; Sikder, A.B.; Ram, S. High altitude forest sensitivity to the recent warming: A tree-ring analysis of conifers from Western Himalaya, India. Quat. Int. 2011, 236, 158–166. [Google Scholar] [CrossRef]
- Walther, G.-R.; Beißner, S.; Pott, R. Climate Change and High Mountain Vegetation Shifts. In Mountain Ecosystems: Studies in Treeline Ecology; Broll, G., Keplin, B., Eds.; Springer Science+Business Media: Berlin, Germany, 2005; pp. 77–96. [Google Scholar]
- Broll, G.; Keplin, B. Mountain Ecosystems: Studies in Treeline Ecology; Springer, Springer Science+Business Media: Berlin, Germany, 2005. [Google Scholar]
- Crawford, R.M.M. Plants at the Margins: Ecological Limits and Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2008; p. 496. [Google Scholar]
- Aakala, T.; Hari, P.; Dengel, S.; Newberry, S.L.; Mizunuma, T.; Grace, J. A prominent stepwise advance of the tree line in north-east Finland. J. Ecol. 2014, 102, 1582–1591. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khandu, Y.; Polthanee, A.; Isarangkool Na Ayutthaya, S. Ecological Dynamics and Regeneration Expansion of Treeline Ecotones in Response to Climate Change in Northern Bhutan Himalayas. Forests 2022, 13, 1062. https://doi.org/10.3390/f13071062
Khandu Y, Polthanee A, Isarangkool Na Ayutthaya S. Ecological Dynamics and Regeneration Expansion of Treeline Ecotones in Response to Climate Change in Northern Bhutan Himalayas. Forests. 2022; 13(7):1062. https://doi.org/10.3390/f13071062
Chicago/Turabian StyleKhandu, Yeshey, Anan Polthanee, and Supat Isarangkool Na Ayutthaya. 2022. "Ecological Dynamics and Regeneration Expansion of Treeline Ecotones in Response to Climate Change in Northern Bhutan Himalayas" Forests 13, no. 7: 1062. https://doi.org/10.3390/f13071062