
The Synergy of Patterns vs. Processes at Community Level: A Key Linkage for Subtropical Native Forests along the Urban Riparian Zone

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Investigation of Forest Communities
2.3. Parameter Calculations
2.3.1. Calculation of Importance Value
2.3.2. Identification of Dominant Species
2.3.3. Animal Community Parameters
2.4. Data Analysis
3. Results
3.1. The Species Composition of Forests and Their Dominant Species in Each Vertical Layer
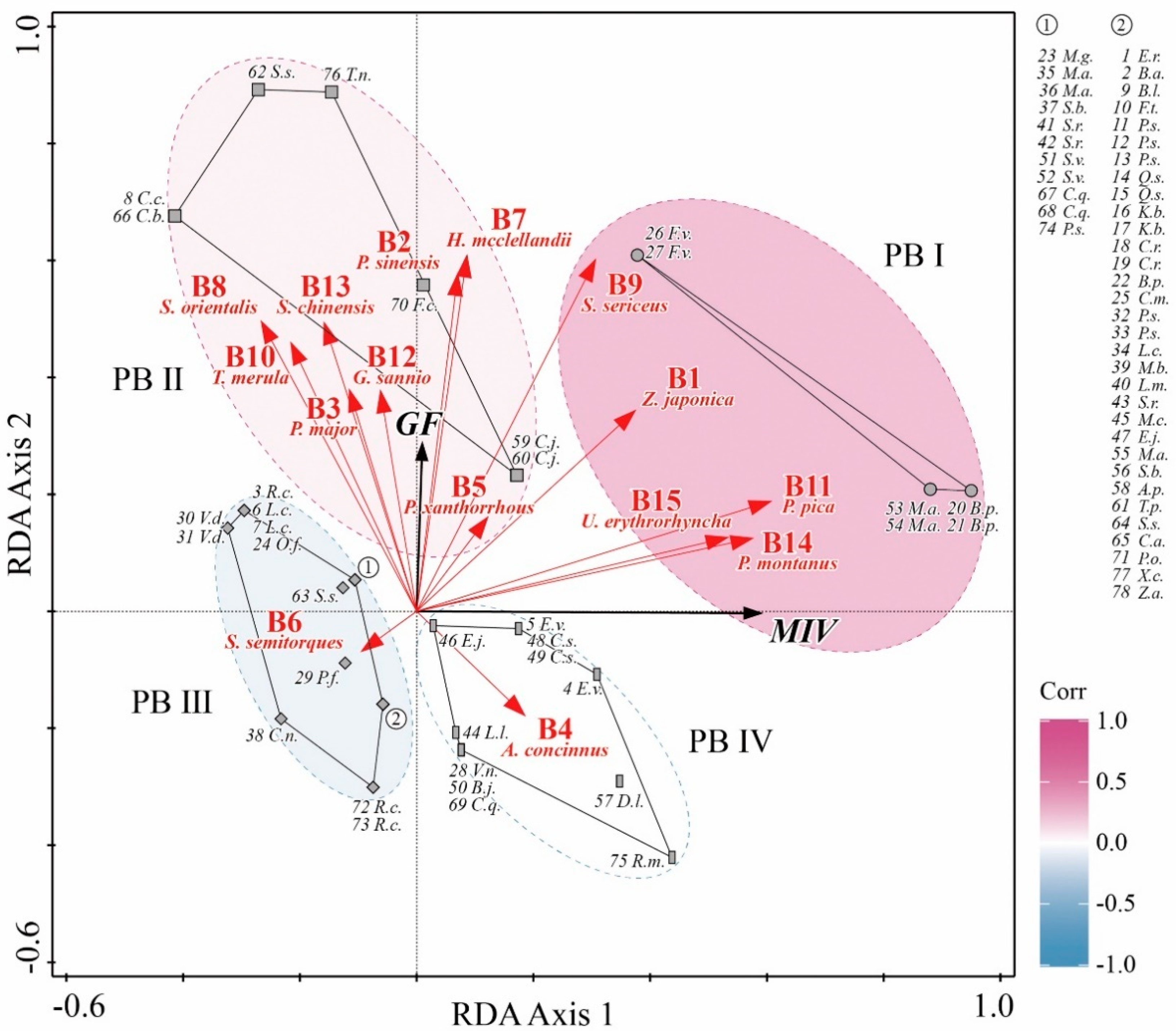
3.2. The Interspecies Associations of Plant Communities
3.3. The Associations and the Closeness between Plants and Birds
4. Discussion
4.1. Characteristics of the Structural Pattern of Forests and Its Significances
4.2. The Aggregation of Plant-Bird into Taxa
4.3. The Advantages of the PB Coevolution and the Significance for Riparian Ecosystem
5. Summary
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones: Focus on links between land and water. BioScience 2010, 41, 540–551. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Dobson, A.; Tilman, D.; Holt, R.D. Unsolved Problems in Ecology; Princeton University Press: Princeton, NJ, USA, 2020. [Google Scholar]
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M. Ecological assembly rules in plant communities—Approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Guimarães Jr, P.R.; Pires, M.M.; Jordano, P.; Bascompte, J.; Thompson, J.N. Indirect effects drive coevolution in mutualistic networks. Nature 2017, 550, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Van der Maarel, E.; Franklin, J. Vegetation Ecology, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Catford, J.A.; Jansson, R. Drowned, buried and carried away: Effects of plant traits on the distribution of native and alien species in riparian ecosystems. New Phytol. 2014, 204, 19–36. [Google Scholar] [CrossRef] [PubMed]
- Stella, J.C.; Rodríguez-González, P.M.; Dufour, S.; Bendix, J. Riparian vegetation research in Mediterranean-climate regions: Common patterns, ecological processes, and considerations for management. Hydrobiologia 2013, 719, 291–315. [Google Scholar] [CrossRef]
- Bennett, A.F.; Nimmo, D.G.; Radford, J.Q. Riparian vegetation has disproportionate benefits for landscape - Scale conservation of woodland birds in highly modified environments. Jour. App. Ecol. 2014, 51, 514–523. [Google Scholar] [CrossRef]
- Forman, R. Land Mosaics: The ecology of landscapes and regions (1995). In The Ecological Design and Planning Reader; Island Press: Washington, DC, USA, 2014; pp. 217–234. [Google Scholar]
- Palmer, M.A.; Hondula, K.L.; Koch, B.J. Ecological restoration of streams and rivers: Shifting strategies and shifting goals. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Naiman, R.J.; Décamps, H. The ecology of interfaces: Riparian zones. Annu. Rev. Ecol. Evol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef] [Green Version]
- Murgui, E.; Hedblom, M. Ecology and Conservation of Birds in Urban environments; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Lai, H.R.; Craven, D.; Hall, J.S.; Hui, F.K.C.; van Breugel, M. Successional syndromes of saplings in tropical secondary forests emerge from environment-dependent trait–demography relationships. Ecol. Lett. 2021, 24, 1776–1787. [Google Scholar] [CrossRef]
- De Bello, F.; Lavorel, S.; Díaz, S.; Harrington, R.; Cornelissen, J.H.; Bardgett, R.D.; Berg, M.P.; Cipriotti, P.; Feld, C.K.; Hering, D. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers. Conserv. 2010, 19, 2873–2893. [Google Scholar] [CrossRef]
- Assis, A.P.; Thompson, J.N.; Santana, P.C.; Jordano, P.; Bascompte, J.; Guimaraes, P., Jr. Genetic correlations and ecological networks shape coevolving mutualisms. Ecol. Lett. 2020, 23, 1789–1799. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.L.; Guzman, L.M.; Meester, D. A process-based metacommunity framework linking local and regional scale community ecology. Ecol. Lett. 2020, 23, 1314–1329. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M.; Jordano, P. Prunus mahaleb and Birds: The High-Efficiency Seed Dispersal System of a Temperate Fruiting Tree. Ecol. Monogr. 1981, 51, 203–218. [Google Scholar] [CrossRef]
- Warren, D.L.; Cardillo, M.; Rosauer, D.F.; Bolnick, D.I. Mistaking geography for biology: Inferring processes from species distributions. Trends Ecol. Evol. 2014, 29, 572–580. [Google Scholar] [CrossRef]
- Moor, H.; Rydin, H.; Hylander, K.; Nilsson, M.B.; Lindborg, R.; Norberg, J. Towards a trait-based ecology of wetland vegetation. J. Ecol. 2017, 105, 1623–1635. [Google Scholar] [CrossRef] [Green Version]
- Leslie, A.B.; Beaulieu, J.M.; Mathews, S. Variation in seed size is structured by dispersal syndrome and cone morphology in conifers and other nonflowering seed plants. New Phytol. 2017, 216, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Escobar, D.F.; Silveira, F.A.; Morellato, L.P.C. Timing of seed dispersal and seed dormancy in Brazilian savanna: Two solutions to face seasonality. Ann. Bot. 2018, 121, 1197–1209. [Google Scholar] [CrossRef]
- Naiman, R.J.; Johnston, C.A.; Kelley, J.C. Alteration of North American streams by beaver. BioSeience 1988, 38, 753–762. [Google Scholar] [CrossRef]
- Sakio, H. Long-Term Ecosystem Changes in Riparian Forests; Springer Nature Singapore Pte Ltd.: Singapore, 2020. [Google Scholar]
- Eriksson, O. Evolution of angiosperm seed disperser mutualisms: The timing of origins and their consequences for coevolutionary interactions between angiosperms and frugivores. Biol. Rev. 2016, 91, 168–186. [Google Scholar] [CrossRef]
- McClure, C.J.; Korte, A.C.; Heath, J.A.; Barber, J.R. Pavement and riparian forest shape the bird community along an urban river corridor. Glob. Ecol. Conserv. 2015, 4, 291–310. [Google Scholar] [CrossRef] [Green Version]
- Nimmo, D.G.; Haslem, A.; Radford, J.Q.; Hall, M.; Bennett, A.F. Riparian tree cover enhances the resistance and stability of woodland bird communities during an extreme climatic event. J. Appl. Ecol. 2016, 53, 449–458. [Google Scholar] [CrossRef]
- Thébault, E.; Fontaine, C. Stability of ecological communities and the architecture of mutualistic and trophic networks. Science 2010, 329, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.J. Urban Ecological Environments: Principles, Approaches and Optimization; China Architecture Publishing & Media Co., Ltd: Beijing, China, 2011. [Google Scholar]
- Pennington, D.N.; Blair, R.B. Habitat selection of breeding riparian birds in an urban environment: Untangling the relative importance of biophysical elements and spatial scale. Divers. Distrib. 2011, 17, 506–518. [Google Scholar] [CrossRef]
- Zhang, J.T. Numerical Ecology, 3rd ed.; Science Press: Beijing, China, 2018. [Google Scholar]
- Yang, C.; Xiong, J.; Zhong, S.; Wang, H.; Li, X. The Classification Table of Vascular Plants in Chongqing; Sichuan Science and Tecknology Press: Chengdu, China, 2009. [Google Scholar]
- Ma, J.H.; Liu, Y.; Lei, J.Y. Methods for Bird Surveys—A Practical Handbook; Hong Kong Bird Watching Society Limited & General: Hong Kong, China, 2006. [Google Scholar]
- Su, H.L.; Xiao, W.F. Status of wintering birdsalong the changjiang river of the Three Gorges. Chin. J. Zool. 2017, 52, 911–936. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University press: Cambridge, UK, 2014. [Google Scholar]
- Zhao, Z.J. A Handbook of the Birds of China; Jilin Science & Technology Press: Changchun, China, 2001. (In Chinese) [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Carlson, M.L.; Flagstad, L.A.; Gillet, F.; Mitchell, E.A. Community development along a proglacial chronosequence: Are above-ground and below-ground community structure controlled more by biotic than abiotic factors? J. Ecol. 2010, 98, 1084–1095. [Google Scholar] [CrossRef] [Green Version]
- Ter, B.; Cajo, J.; Šmilauer, P.; Dray, S. Algorithms and biplots for double constrained correspondence analysis. Environ. Ecol. Stat. 2018, 25, 171–197. [Google Scholar]
- Turner, M.G. Landscape ecology: What is the state of the science? Annu. Rev. Ecol. Evol. Syst. 2005, 36, 319–344. [Google Scholar] [CrossRef]
- Kassambara, A. ggcorrplot: Visualization of a Correlation Matrix Using ’ggplot2’. R Package Version 0.1.3. 2019. Available online: https://CRAN.R-project.org/package=ggcorrplot (accessed on 15 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Clark, J.S.; Lewis, M.; McLachlan, J.S.; HilleRisLambers, J. Estimating population spread: What can we forecast and how well? Ecology 2003, 84, 1979–1988. [Google Scholar] [CrossRef]
- Hendriks, A.J.; Mulder, C. Scaling of offspring number and mass to plant and animal size: Model and meta-analysis. Oecologia 2008, 155, 705–716. [Google Scholar] [CrossRef] [Green Version]
- Begon, M.; Townsend, C.R.; Harper, J.L. Ecology: From Individuals to Ecosystems; John Wiley & Sons: New York, NY, USA, 2006. [Google Scholar]
- Althoff, D.M.; Segraves, K.A.; Johnson, M.T. Testing for coevolutionary diversification: Linking pattern with process. Trends Ecol. Evol. 2014, 29, 82–89. [Google Scholar] [CrossRef]
- Duchenne, F.; Fontaine, C.; Teulière, E.; Thébault, E. Phenological traits foster persistence of mutualistic networks by promoting facilitation. Ecol. Lett. 2021, 24, 2088–2099. [Google Scholar] [CrossRef]
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef]
- Pugnaire, F.; Valladares, F. Functional Plant Ecology; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Rood, S.B.; Kaluthota, S.; Gill, K.M.; Hillman, E.J.; Woodman, S.G.; Pearce, D.W.; Mahoney, J.M. A twofold strategy for riparian restoration: Combining a functional flow regime and direct seeding to re-establish cottonwoods. River Res. Appl. 2016, 32, 836–844. [Google Scholar] [CrossRef]
- Madejon, P.; Ciadamidaro, L.; Maranon, T.; Murillo, J.M. Long-term biomonitoring of soil contamination using poplar trees: Accumulation of trace elements in leaves and fruits. Int. J. Phytoremediat. 2013, 15, 602–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brookshire, E.N.J.; Kauffman, J.B.; Lytjen, D.; Otting, N. Cumulative effects of wild ungulate and livestock herbivory on riparian willows. Oecologia 2002, 132, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Daru, B.H.; Elliott, T.L.; Park, D.S.; Davies, T.J. Understanding the processes underpinning patterns of phylogenetic regionalization. Trends Ecol. Evol. 2017, 32, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Lambert, F.R.; Marshall, A.G. Keystone characteristics of bird-dispersed Ficus in a Malaysian lowland rain forest. J. Ecol. 1991, 79, 793–809. [Google Scholar] [CrossRef] [Green Version]
- Bastolla, U.; Fortuna, M.A.; Pascual-García, A.; Ferrera, A.; Luque, B.; Bascompte, J. The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature 2009, 458, 1018–1020. [Google Scholar] [CrossRef]
- Chen, H.; Niklas, K.J.; Yang, D.; Sun, S. The effect of twig architecture and seed number on seed size variation in subtropical woody species. New Phytol. 2009, 183, 1212–1221. [Google Scholar] [CrossRef]
- Sinnott-Armstrong, M.A.; Donoghue, M.J.; Jetz, W.J. Dispersers and environment drive global variation in fruit colour syndromes. Ecol. Lett. 2021, 24, 1387–1399. [Google Scholar] [CrossRef]
- Yu, K.J.; Li, D.H.; Duan, T.W. Landscape approaches in biodiversity conservation. Chin. Biodivers. 1998, 6, 205–212. [Google Scholar]
- Munné, A.; Prat, N.; Solà, C.; Bonada, N.; Rieradevall, M. A simple field method for assessing the ecological quality of riparian habitat in rivers and streams: QBR index. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, 147–163. [Google Scholar] [CrossRef]
- Welsh, H.H.; Hodgson, G.R.; Karraker, N.E. Influences of the vegetation mosaic on riparian and stream environments in a mixed forest-grassland landscape in "Mediterranean" northwestern California. Ecography 2005, 28, 537–551. [Google Scholar] [CrossRef]
- Mamun, M.; An, K.G. Ecological health assessments of 72 streams and rivers in relation to water chemistry and land-use patterns in South Korea. Turk. J. Fish Aquat. Sci. 2018, 18, 871–880. [Google Scholar] [CrossRef]
- Yu, X.; Yang, J.; Liu, L.; Tian, Y.; Yu, Z. Effects of Spartina alterniflora invasion on biogenic elements in a subtropical coastal mangrove wetland. Environ. Sci. Pollut. R. 2015, 22, 3107–3115. [Google Scholar] [CrossRef]
- Gayen, J.; Ghosh, P.K.; Datta, D. Avifauna habitability of wetlands in an intensive cultural landscape: Insights from the Ichhamati floodplains, India. Environ. Monit. Assess. 2020, 192, 1–16. [Google Scholar] [CrossRef]
- Izzati, U.; Hasibuan, H.S. Riparian tree and bird diversity in Cisadane River, South Tangerang City, Indonesia. Biodivers. J. Biol. Divers. 2019, 20, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E.; Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef]
- Allesina, S.; Tang, S. Stability criteria for complex ecosystems. Nature 2012, 483, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Rohr, R.P.; Saavedra, S.; Bascompte, J. On the structural stability of mutualistic systems. Science 2014, 1253497. [Google Scholar] [CrossRef] [Green Version]
- Howe, H.F. Making dispersal syndromes and networks useful in tropical conservation and restoration. Glob. Ecol. Conserv. 2016, 6, 152–178. [Google Scholar] [CrossRef] [Green Version]
- Stouffer, P.C.; Jirinec, V.; Rutt, C.L.; Bierregaard, R.O., Jr.; Hernández-Palma, A.; Johnson, E.I.; Midway, S.R.; Powell, L.L.; Wolfe, J.D.; Lovejoy, T.E. Long-term change in the avifauna of undisturbed Amazonian rainforest: Ground-foraging birds disappear and the baseline shifts. Ecol. Lett. 2021, 24, 186–195. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
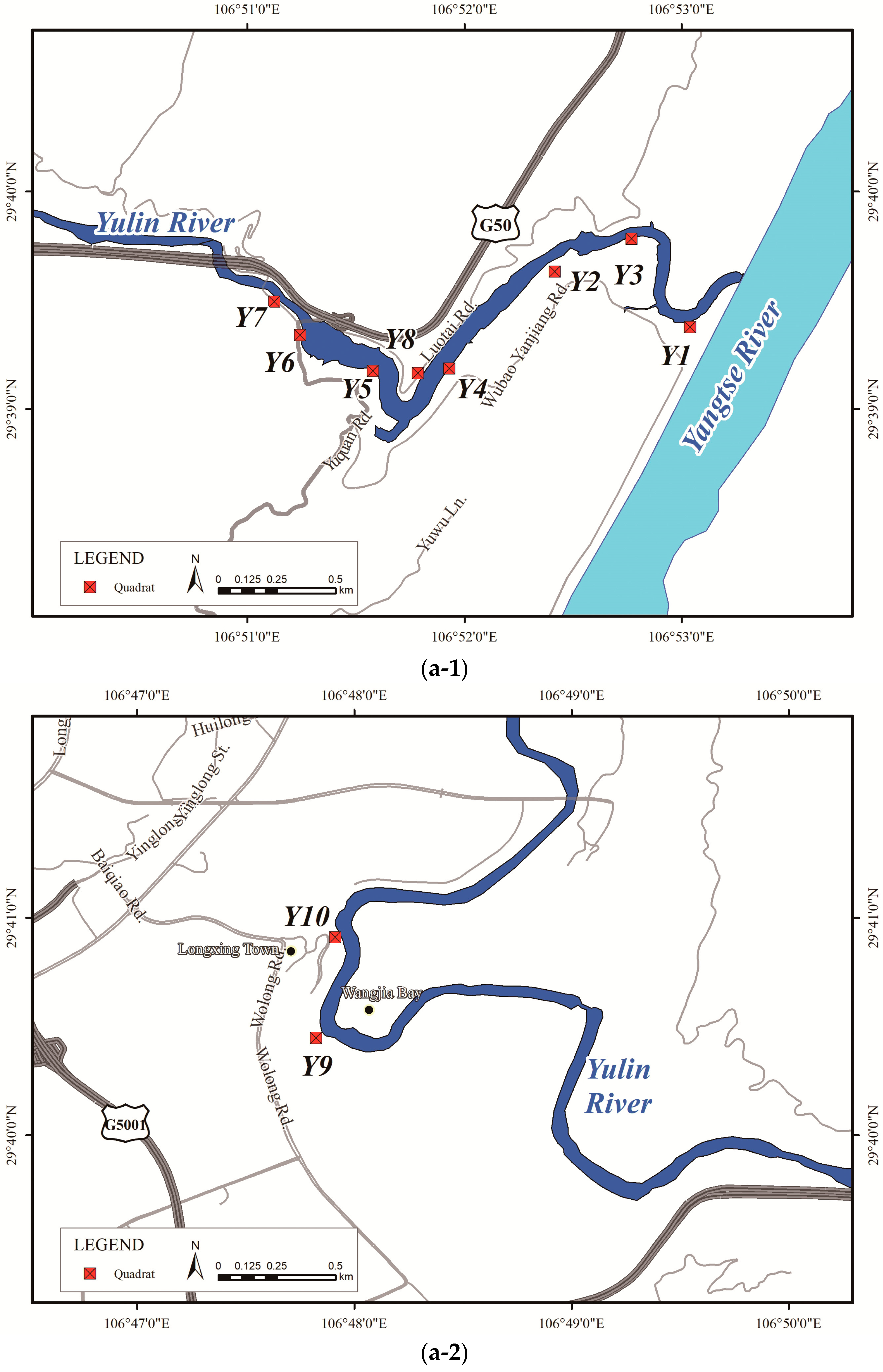
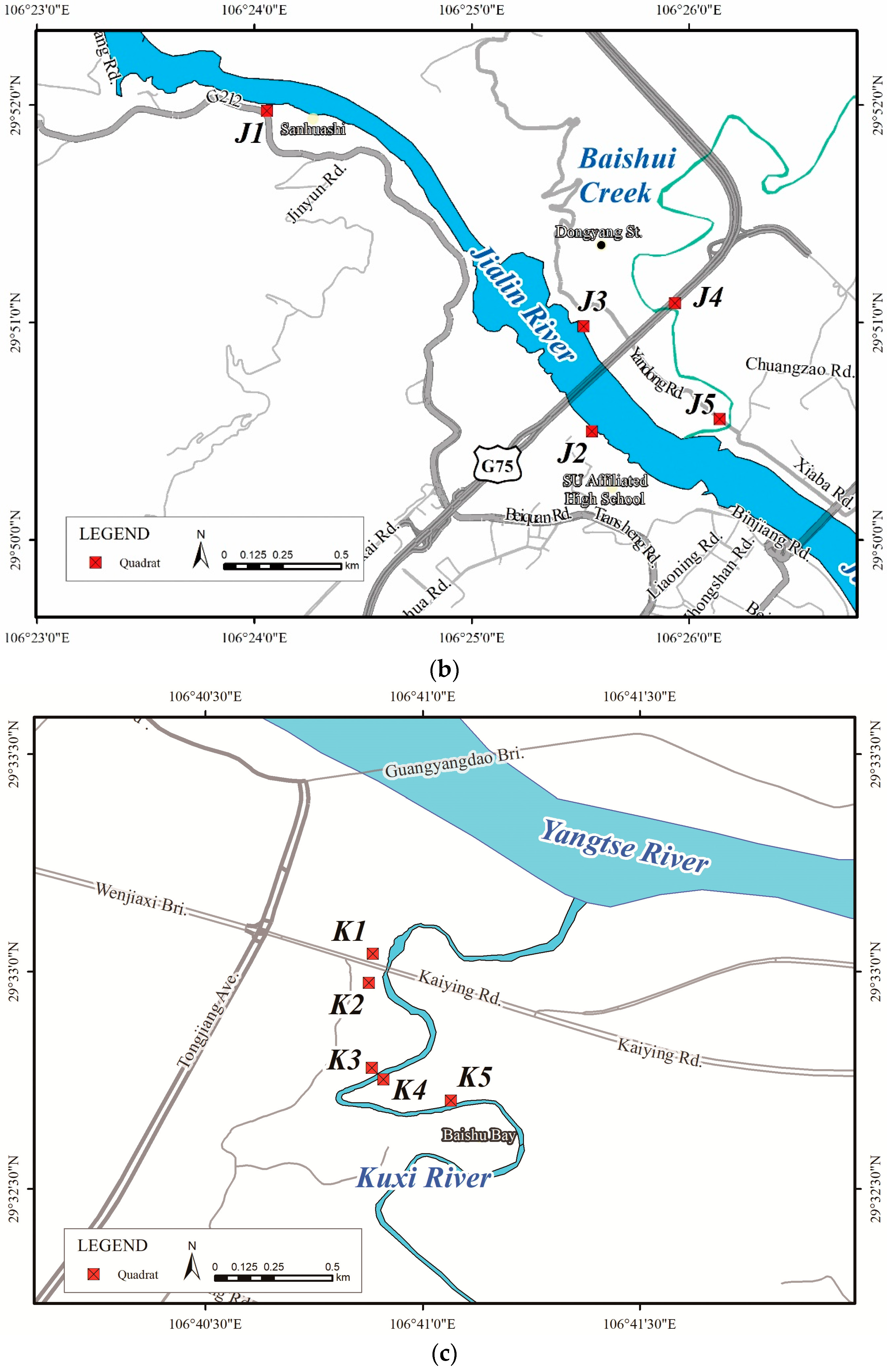
| River Name | Site No. | Site Location | Dry Water Level (m) | Elevation Range (m) | Horizontal Distance (m) | Longitude and Latitude | Slope Direction | Vegetation Coverage (%) | Surrounding Environment | Area Type | Description |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Jialing River | J1 | Near Sanhuashi, Beibei District | 186 | 30 | 90 | 106°24′18″ N, 29°51′49″ E | 26° NE | 60 | Farmland | Suburban | The soil layer near the riverbanks is thin and dominated by stones, while the soil layer from a higher place is thick; woody plants are natural and concentrated; the upper ground is mainly farmland. |
| J2 | Near Southwest University Affiliated High School, Beibei District | 181 | 30 | 90–155 | 106°25′48″ N, 29°50′21″ E | 55° NE | 63 | Residential area | Edge of built-up area | The soils are thick on highland, with sand and gravel dominating near the shores; the site is natural, with high herbaceous plant cover and robust woody plant growth. | |
| J3 | Near Dongyang Street, Beibei District | 181 | 20 | 190 | 106°25′45″ N, 29°50′49″ E | 185° S | 22 | Mulberry grove | Suburban | The sand and gravels are widely distributed, and the soil is poor; the natural vegetation is scattered along the gentle slopes; the higher areas are regularly concentrated and accompanied by farmland. | |
| J4 | Near Baishui Creek (a tributary of Jialing River), Beibei District | 191 | 6 | 30–50 | 106°25′45″ N, 29°50′49″ E | 192° S | 80 | Woodland | Suburban | The river is shallow and narrow; the subsidence area is gently sloping and silty near the waterfront; the bamboo cover is high; the woody plants are distributed in the high area above the steep slope. | |
| J5 | Near the inlet of Baishui Creek, Beibei District | 186 | 7 | 67 | 106°26′23″ N, 29°50′24″ E | 164° S | 75 | Woodland | Suburban | The soil layer of the bank is thick; the slope of the riverbank is high; the woody vegetation is patchy and robust. | |
| Yulin River | Y1 | Near the inlet, Jiangbei District | 175 | 10 | 31 | 106°53′16″ N, 29°39′12″ E | 339° NE | 90 | Highway | Suburban | A concave shore area with a gentle slope near the banks, and most cultivated. |
| Y2 | Near Wubao Yanjiang Rd., Jiangbei District | 173 | 20 | 50 | 106°52′40″ N, 29°39′27″ E | 314° NW | 50 | Highway | Suburban | The site is under reconstruction, with steeper slopes near the water shore, relatively high hardened surface ratio, and thicker soil layers; the woody plants are natural and distributed in a patchy pattern. | |
| Y3 | Near Wubao Yanjiang Rd., Jiangbei District | 167 | 18 | 88 | 106°53′0″ N, 29°39′35″ E | 25° N | 75 | Woodland | Suburban | The banks are convex and curved, with farmland on the banks; woody plants are intensive; fruit trees such as citrus and loquats are planted as well. | |
| Y4 | Under viaduct of Wubao Yanjiang Rd., Jiangbei District | 176 | 20 | 57 | 106°52′9″ N, 29°39′1″ E | 292° W | 65 | Highway | Suburban | The slope is steep, and the vegetation is patchy; fruit trees such as peach, plum, and citrus are planted on the banks. | |
| Y5 | Under viaduct of Yuquan Rd., Jiangbei District | 168 | 20 | 95 | 106°51′49″ N, 29°38′59″ E | 42° NE | 65 | Highway | Suburban | Small patches of farmland area on the banks, and the slope of the higher place is steep; woody plants are natural and distributed in patches, with a large herbaceous cover. | |
| Y6 | Near Yuquan Rd., Jiangbei District | 172 | 20 | 50 | 106°51′29″ N, 29°39′10″ E | 89° E | 70 | Highway | Suburban | The banks are steep and natural, with a high tall grass cover; woody plants (mostly maple poplar) are planted artificially near the waterfront. | |
| Y7 | Near Wujian Rd., Jiangbei District | 182 | 20 | 36 | 106°51′22″ N, 29°39′19″ E | 48° NE | 45 | Highway | Suburban | The banks are cliff-like and steep, with poor soil and scattered woody plants. | |
| Y8 | Near Luotai Rd., Yubei District | 176 | 10 | 24 | 106°52′2″ N, 29°38′59″ E | 110° E | 80 | Highway | Suburban | The soil layer is thick and fertile, while the slope is high; the banks are covered by both cultivated and natural woody plants with a high herbaceous cover under the forest. | |
| Y9 | Near Wangjia Bay, Jiangbei District | 177 | 10 | 123 | 106°48′5″ N, 29°40′17″ E | 69° E | 35 | Highway | Suburban | The shore is concave and will be under construction soon; woody plants are robust and concentrated along the river; higher bare land is covered by herbage. | |
| Y10 | Near Longxing Town, Yubei District | 172 | 5 | 90 | 106°48′9″ N, 29°40′45″ E | 75° E | 85 | Parks | Suburban | Patches of woody plants are concentrated along the gentle slope near the banks; the higher ground is a park under planning with artificial vegetation coverage. | |
| Kuxi River | K1 | Under bridge of Kaiying Rd., Nanan District | 176 | 10 | 51 | 106°41′8″ N, 29°32′52″ E | 127° SE | 85 | Woodland | Suburban | The land is steep and intact near the shores; the soil layer is thick on the gentle highland, covered by dense woody plants patches. |
| K2 | Under bridge of Kaiying Rd., Nanan District | 176 | 10 | 69 | 106°41′7″ N, 29°32′48″ E | 86° E | 80 | Highway | Suburban | Soil is fertile along the gentle riverbanks, and the area is largely used for agriculture; the vegetation cover from higher ground is natural and dense. | |
| K3 | Nanan District | 175 | 8 | 64 | 106°41′7″ N, 29°32′36″ E | 145° SE | 80 | Residential areas | Suburban | Woody plants patches are dense and distributed on the gentle and fertile banks. | |
| K4 | Nanan District | 175 | 8 | 31 | 106°41′9″ N, 29°32′35″ E | 326° NW | 85 | Woodland | Suburban | The banks are steep; woody plants are intact and patchy. | |
| K5 | Across Baishu Bay, Nanan District | 177 | 8 | 26 | 106°41′18″ N, 29°32′32″ E | 190° S | 70 | Residential area/woodland | Suburban | The banks are gentle and fertile; dense woody plants are distributed in patches, with a high herbage coverage under the forest. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wang, D.; Shi, Y.; Xu, Z.; Chen, H. The Synergy of Patterns vs. Processes at Community Level: A Key Linkage for Subtropical Native Forests along the Urban Riparian Zone. Forests 2022, 13, 1041. https://doi.org/10.3390/f13071041
Wang H, Wang D, Shi Y, Xu Z, Chen H. The Synergy of Patterns vs. Processes at Community Level: A Key Linkage for Subtropical Native Forests along the Urban Riparian Zone. Forests. 2022; 13(7):1041. https://doi.org/10.3390/f13071041
Chicago/Turabian StyleWang, Haiyang, Danrong Wang, Ya Shi, Zhiqing Xu, and Hong Chen. 2022. "The Synergy of Patterns vs. Processes at Community Level: A Key Linkage for Subtropical Native Forests along the Urban Riparian Zone" Forests 13, no. 7: 1041. https://doi.org/10.3390/f13071041