
Evidence for Alternate Stable States in an Ecuadorian Andean Cloud Forest

 , and
, and
Abstract
:1. Introduction
2. Methods
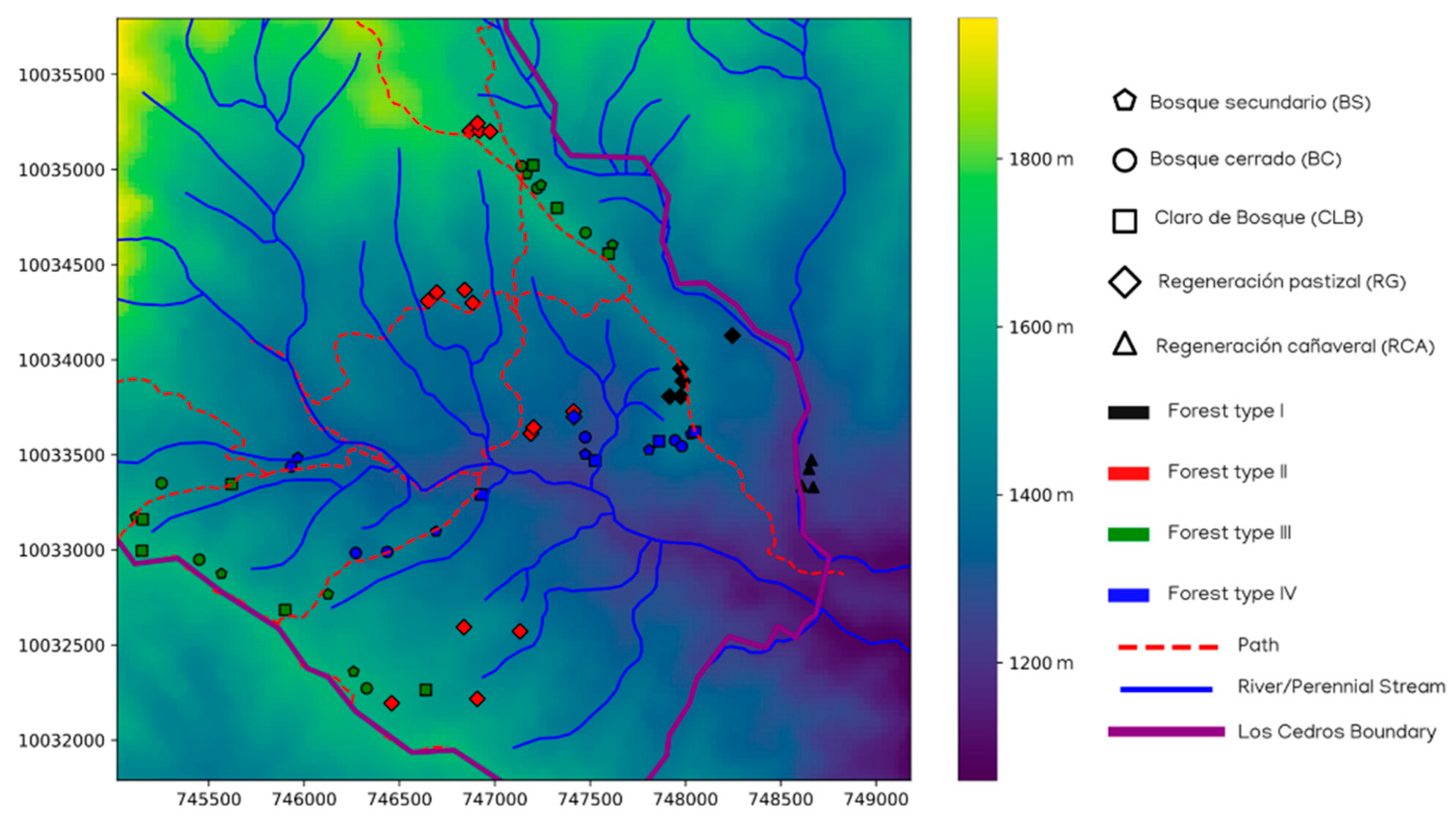
2.1. Site—Reserva Los Cedros
2.2. Tree Survey and Plant Identification
2.2.1. Selection of Sites and Categorization of Land-Use History
2.2.2. Survey Methods
2.3. Statistical and Informatic Methods
2.3.1. Species Accumulation Curves and Richness Estimators
2.3.2. Tree Community Turnover (Distance Decay or Beta Diversity)
Turnover by Physical Distance
Asymptotic Model
Linear Model
Model Comparison
Overall Turnover by Watershed Crossings
2.3.3. Ordination by Historical Land Use/Habitat
2.3.4. GIS Data and Additional Environmental Data
2.3.5. Hierarchical Clustering of Sites
2.3.6. Prediction of Current Ecological State by Land Use History and Elevation
2.3.7. Indicator Species Analysis
2.3.8. Spatial Analyses
2.3.9. Juvenile Communities
2.3.10. Deforestation in the Region
3. Results
3.1. Species Accumulation Curves and Richness Estimators
3.2. Tree Community Turnover (Distance Decay)
3.2.1. Turnover by Physical Distance
- Asymptotic model
- 2.
- Linear model
3.2.2. Overall Turnover by Watershed Crossings
3.3. Hierarchical Clustering of Sites and Prediction of Current Ecological State by Land-Use History and Elevation
3.4. Indicator Species Analysis
3.5. Spatial Analyses
3.6. Juvenile Communities
3.7. Deforestation in the Region
4. Discussion
4.1. Stable States in the Andean Cloud Forest
4.1.1. Soil Structure
4.1.2. Soil Seed Bank Depletion
4.1.3. Large Seeded Plants and Animal Dispersal
4.1.4. Relative Fluxes of Local vs. Exotic Plant Types
4.2. Juvenile Tree Community
4.3. Beta Diversity and Spatial Heterogeneity in the Andean Cloud Forest
4.4. Conservation Value of Los Cedros
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Bunin, G. Directionality and Community-Level Selection. Oikos 2021, 130, 489–500. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Negron-Juarez, R.I.; Marra, D.M.; Di Vittorio, A.; Tews, J.; Roberts, D.; Ribeiro, G.H.; Trumbore, S.E.; Higuchi, N. The Steady-State Mosaic of Disturbance and Succession across an Old-Growth Central Amazon Forest Landscape. Proc. Natl. Acad. Sci. USA 2013, 110, 3949–3954. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Turner, B.L. Ecological Succession in a Changing World. J. Ecol. 2019, 107, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; Carnegie Institution of Washington: Washington, DC, USA, 1916; ISBN 0-598-48534-1. [Google Scholar]
- Connell, J.H.; Slatyer, R.O. Mechanisms of Succession in Natural Communities and Their Role in Community Stability and Organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Cowles, H.C. The Ecological Relations of the Vegetation on the Sand Dunes of Lake Michigan, Part I—Geographical Relations of the Dune Floras. Bot. Gaz. 1899, 27, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Curtis, J.T.; Greene, H.C. A Study of Relic Wisconsin Prairies by the Species-Presence Method. Ecology 1949, 30, 83–92. [Google Scholar] [CrossRef]
- Gleason, H.A. The Individualistic Concept of the Plant Association. Bull. Torrey Bot. Club 1926, 53, 7–26. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, R.H. Vegetation of the Great Smoky Mountains. Ecol. Monogr. 1956, 26, 1–80. [Google Scholar] [CrossRef]
- Norden, N.; Angarita, H.A.; Bongers, F.; Martínez-Ramos, M.; Granzow-de la Cerda, I.; Van Breugel, M.; Lebrija-Trejos, E.; Meave, J.A.; Vandermeer, J.; Williamson, G.B.; et al. Successional Dynamics in Neotropical Forests Are as Uncertain as They Are Predictable. Proc. Natl. Acad. Sci. USA 2015, 112, 8013–8018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, P.W. The Tropical Rain Forest; Cambridge University Press: London, UK, 1952. [Google Scholar]
- HilleRisLambers, J.; Adler, P.B.; Harpole, W.S.; Levine, J.M.; Mayfield, M.M. Rethinking Community Assembly through the Lens of Coexistence Theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 227–248. [Google Scholar] [CrossRef] [Green Version]
- Young, T.P.; Chase, J.M.; Huddleston, R.T. Community Succession and Assembly Comparing, Contrasting and Combining Paradigms in the Context of Ecological Restoration. Ecol. Rest. 2001, 19, 5–18. [Google Scholar] [CrossRef]
- Fisher, C.K.; Mehta, P. The Transition between the Niche and Neutral Regimes in Ecology. Proc. Natl. Acad. Sci. USA 2014, 111, 13111–13116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbell, S.P. Approaching Ecological Complexity from the Perspective of Symmetric Neutral Theory. Trop. For. Community Ecol. 2008, 143, 159. [Google Scholar]
- Keddy, P.A. Assembly and Response Rules: Two Goals for Predictive Community Ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, D.C.; Laughlin, D.E. Advances in Modeling Trait-Based Plant Community Assembly. Trends Plant Sci. 2013, 18, 584–593. [Google Scholar] [CrossRef]
- Ulrich, W.; Zaplata, M.K.; Winter, S.; Schaaf, W.; Fischer, A.; Soliveres, S.; Gotelli, N.J. Species Interactions and Random Dispersal Rather than Habitat Filtering Drive Community Assembly during Early Plant Succession. Oikos 2016, 125, 698–707. [Google Scholar] [CrossRef] [Green Version]
- Bhaskar, R.; Dawson, T.E.; Balvanera, P. Community Assembly and Functional Diversity along Succession Post-Management. Funct. Ecol. 2014, 28, 1256–1265. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Meiners, S.J. Ever since Clements: From Succession to Vegetation Dynamics and Understanding to Intervention. Appl. Veg. Sci. 2009, 12, 9–21. [Google Scholar] [CrossRef]
- Norden, N.; Chazdon, R.L.; Chao, A.; Jiang, Y.-H.; Vílchez-Alvarado, B. Resilience of Tropical Rain Forests: Tree Community Reassembly in Secondary Forests. Ecol. Lett. 2009, 12, 385–394. [Google Scholar] [CrossRef]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s Sixth Mass Extinction Already Arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H. (Ted); et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Carmody, M.; Waszczak, C.; Idänheimo, N.; Saarinen, T.; Kangasjärvi, J. ROS Signalling in a Destabilised World: A Molecular Understanding of Climate Change. J. Plant Physiol. 2016, 203, 69–83. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Day, J.W.; Christian, R.R.; Boesch, D.M.; Yáñez-Arancibia, A.; Morris, J.; Twilley, R.R.; Naylor, L.; Schaffner, L.; Stevenson, C. Consequences of Climate Change on the Ecogeomorphology of Coastal Wetlands. Estuaries Coasts 2008, 31, 477–491. [Google Scholar] [CrossRef]
- Doney, S.C.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate Change Impacts on Marine Ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [Green Version]
- Fadrique, B.; Báez, S.; Duque, Á.; Malizia, A.; Blundo, C.; Carilla, J.; Osinaga-Acosta, O.; Malizia, L.; Silman, M.; Farfán-Ríos, W. Widespread but Heterogeneous Responses of Andean Forests to Climate Change. Nature 2018, 564, 207. [Google Scholar] [CrossRef]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fulé, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread Increase of Tree Mortality Rates in the Western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel Ecosystems: Implications for Conservation and Restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.C.; Goldewijk, K.K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic Transformation of the Biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19, 589–606. [Google Scholar] [CrossRef]
- Hautier, Y.; Tilman, D.; Isbell, F.; Seabloom, E.W.; Borer, E.T.; Reich, P.B. Anthropogenic Environmental Changes Affect Ecosystem Stability via Biodiversity. Science 2015, 348, 336–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. Forest Soils and Carbon Sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Odum, E.P. The Strategy of Ecosystem Development. In The Ecological Design and Planning Reader; Springer: Berlin/Heidelberg, Germany, 2014; pp. 203–216. [Google Scholar]
- Gundersen, P.; Thybring, E.E.; Nord-Larsen, T.; Vesterdal, L.; Nadelhoffer, K.J.; Johannsen, V.K. Old-Growth Forest Carbon Sinks Overestimated. Nature 2021, 591, E21–E23. [Google Scholar] [CrossRef] [PubMed]
- Luyssaert, S.; Schulze, E.-D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-Growth Forests as Global Carbon Sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Lewis, S.L.; Wheeler, C.E.; Mitchard, E.T.A.; Koch, A. Restoring Natural Forests Is the Best Way to Remove Atmospheric Carbon. Nature 2019, 568, 25–28. [Google Scholar] [CrossRef]
- Moomaw, W.R.; Masino, S.A.; Faison, E.K. Intact Forests in the United States: Proforestation Mitigates Climate Change and Serves the Greatest Good. Front. For. Glob. Chang. 2019, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Sacco, A.D.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.S.; Breman, E.; Rebola, L.C.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten Golden Rules for Reforestation to Optimize Carbon Sequestration, Biodiversity Recovery and Livelihood Benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Evans, T.; Venter, O.; Williams, B.; Tulloch, A.; Stewart, C.; Thompson, I.; Ray, J.C.; Murray, K.; Salazar, A.; et al. The Exceptional Value of Intact Forest Ecosystems. Nat. Ecol. Evol. 2018, 2, 599–610. [Google Scholar] [CrossRef]
- Harmon, M.E.; Ferrell, W.K.; Franklin, J.F. Effects on Carbon Storage of Conversion of Old-Growth Forests to Young Forests. Science 1990, 247, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of Tree Carbon Accumulation Increases Continuously with Tree Size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Mitchard, E.T.A. The Tropical Forest Carbon Cycle and Climate Change. Nature 2018, 559, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Girardin, C.A.J.; Espejob, J.E.S.; Doughty, C.E.; Huasco, W.H.; Metcalfe, D.B.; Durand-Baca, L.; Marthews, T.R.; Aragao, L.E.O.C.; Farfán-Rios, W.; García-Cabrera, K.; et al. Productivity and Carbon Allocation in a Tropical Montane Cloud Forest in the Peruvian Andes. Plant Ecol. Divers. 2014, 7, 107–123. [Google Scholar] [CrossRef]
- Spracklen, D.V.; Righelato, R. Tropical Montane Forests Are a Larger than Expected Global Carbon Store. Biogeosciences 2014, 11, 2741–2754. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.A.; Zorrilla, M.; Endara, L.; Thomas, D.C.; Vandegrift, R.; Rubenstein, J.M.; Policha, T.; Rios-Touma, B.; Read, M. New Mining Concessions Could Severely Decrease Biodiversity and Ecosystem Services in Ecuador. Trop. Conserv. Sci. 2018, 11, 1940082918780427. [Google Scholar] [CrossRef]
- Draper, F.C.; Asner, G.P.; Coronado, E.N.H.; Baker, T.R.; García-Villacorta, R.; Pitman, N.C.A.; Fine, P.V.A.; Phillips, O.L.; Gómez, R.Z.; Guerra, C.A.A.; et al. Dominant Tree Species Drive Beta Diversity Patterns in Western Amazonia. Ecology 2019, 100, e02636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencia, R.; Balslev, H.; Miño, G.P.Y. High Tree Alpha-Diversity in Amazonian Ecuador. Biodivers. Conserv. 1994, 3, 21–28. [Google Scholar] [CrossRef]
- Brehm, G.; Hebert, P.D.N.; Colwell, R.K.; Adams, M.-O.; Bodner, F.; Friedemann, K.; Möckel, L.; Fiedler, K. Turning Up the Heat on a Hotspot: DNA Barcodes Reveal 80% More Species of Geometrid Moths along an Andean Elevational Gradient. PLoS ONE 2016, 11, e0150327. [Google Scholar] [CrossRef] [PubMed]
- Churchill, S.P. Moss diversity and endemism of the tropical andes. Ann. Mo. Bot. Gard. 2009, 96, 434–449. [Google Scholar] [CrossRef]
- Gentry, A.H.; Dodson, C.H. Diversity and Biogeography of Neotropical Vascular Epiphytes. Ann. Mo. Bot. Gard. 1987, 74, 205. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.E. The Tropical Andean Plant Diversity Powerhouse. New Phytol. 2016, 210, 1152–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küper, W.; Kreft, H.; Nieder, J.; Köster, N.; Barthlott, W. Large-Scale Diversity Patterns of Vascular Epiphytes in Neotropical Montane Rain Forests. J. Biogeogr. 2004, 31, 1477–1487. [Google Scholar] [CrossRef]
- Olson, D.M. The Distribution of Leaf Litter Invertebrates Along a Neotropical Altitudinal Gradient. J. Trop. Ecol. 1994, 10, 129–150. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Barahona, S.; Luna-Vega, I.; Tejero-Díez, D. Species Richness, Endemism, and Conservation of American Tree Ferns (Cyatheales). Biodivers. Conserv. 2011, 20, 59–72. [Google Scholar] [CrossRef]
- Ríos-Touma, B.; Holzenthal, R.W.; Huisman, J.; Thomson, R.; Rázuri-Gonzales, E. Diversity and Distribution of the Caddisflies (Insecta: Trichoptera) of Ecuador. PeerJ 2017, 5, e2851. [Google Scholar] [CrossRef]
- Smith, S.A.; Oca, A.N.M.D.; Reeder, T.W.; Wiens, J.J. A Phylogenetic Perspective on Elevational Species Richness Patterns in Middle American Treefrogs: Why so Few Species in Lowland Tropical Rainforests? Evolution 2007, 61, 1188–1207. [Google Scholar] [CrossRef] [PubMed]
- Borchsenius, F. Patterns of Plant Species Endemism in Ecuador. Biodivers. Conserv. 1997, 6, 379–399. [Google Scholar] [CrossRef]
- Young, K.R.; Ulloa, C.U.; Luteyn, J.L.; Knapp, S. Plant Evolution and Endemism in Andean South America: An Introduction. Bot. Rev. 2002, 68, 4–21. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Janzen, D.H. Why Mountain Passes Are Higher in the Tropics. Am. Nat. 1967, 101, 233–249. [Google Scholar] [CrossRef]
- Von Humboldt, A.; Bonpland, A. Essay on the Geography of Plants; University of Chicago Press: Chicago, IL, USA, 1807; ISBN 0-226-36068-7. [Google Scholar]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, Water, and Broad-Scale Geographic Patterns of Species Richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Arita, H.T.; Vázquez-Domínguez, E. The Tropics: Cradle, Museum or Casino? A Dynamic Null Model for Latitudinal Gradients of Species Diversity. Ecol. Lett. 2008, 11, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, D.; Roy, K.; Valentine, J.W. Out of the Tropics: Evolutionary Dynamics of the Latitudinal Diversity Gradient. Science 2006, 314, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Gentry, A.H. Tropical Forest Biodiversity: Distributional Patterns and Their Conservational Significance. Oikos 1992, 63, 19. [Google Scholar] [CrossRef]
- Särkinen, T.; Pennington, R.T.; Lavin, M.; Simon, M.F.; Hughes, C.E. Evolutionary Islands in the Andes: Persistence and Isolation Explain High Endemism in Andean Dry Tropical Forests. J. Biogeogr. 2012, 39, 884–900. [Google Scholar] [CrossRef]
- Corbin, J.D.; Holl, K.D. Applied Nucleation as a Forest Restoration Strategy. For. Ecol. Manag. 2012, 265, 37–46. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Holl, K.D.; Cole, R.J.; Reid, J.L. Testing Applied Nucleation as a Strategy to Facilitate Tropical Forest Recovery. J. Appl. Ecol. 2013, 50, 88–96. [Google Scholar] [CrossRef]
- Loughlin, N.J.D.; Gosling, W.D.; Mothes, P.; Montoya, E. Ecological Consequences of Post-Columbian Indigenous Depopulation in the Andean–Amazonian Corridor. Nat. Ecol. Evol. 2018, 2, 1233–1236. [Google Scholar] [CrossRef]
- Tobler, W.R. A Computer Movie Simulating Urban Growth in the Detroit Region. Econ. Geogr. 1970, 46, 234–240. [Google Scholar] [CrossRef]
- Londoño-Murcia, M.C.; Tellez-Valdés, O.; Sánchez-Cordero, V. Environmental Heterogeneity of World Wildlife Fund for Nature Ecoregions and Implications for Conservation in Neotropical Biodiversity Hotspots. Environ. Conserv. 2010, 37, 116–127. [Google Scholar] [CrossRef]
- Pérez-Escobar, O.A.; Lucas, E.; Jaramillo, C.; Monro, A.; Morris, S.K.; Bogarín, D.; Greer, D.; Dodsworth, S.; Aguilar-Cano, J.; Sanchez Meseguer, A.; et al. The Origin and Diversification of the Hyperdiverse Flora in the Chocó Biogeographic Region. Front. Plant Sci. 2019, 10, 1328. [Google Scholar] [CrossRef] [PubMed]
- Stadtmüller, T. Cloud Forests in the Humid Tropics: A Bibliographic Review; United Nations University Press: Tokyo, Japan; Turrialba, Costa Rica, 1987; ISBN 92-808-0670-X. [Google Scholar]
- Foster, P. The Potential Negative Impacts of Global Climate Change on Tropical Montane Cloud Forests. Earth-Sci. Rev. 2001, 55, 73–106. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Whitmore, T.C. A Comparison of Montane and Lowland Rain Forest in Ecuador I. The Forest Structure, Physiognomy, and Floristics. J. Ecol. 1963, 51, 567. [Google Scholar] [CrossRef]
- Grubb, P.J.; Whitmore, T.C. A Comparison of Montane and Lowland Rain Forest in Ecuador: II. The Climate and Its Effects on the Distribution and Physiognomy of the Forests. J. Ecol. 1966, 54, 303. [Google Scholar] [CrossRef]
- Policha, T. Pollination Biology of the Mushroom-Mimicking Orchid Genus Dracula; University of Oregon: Eugene, OR, USA, 2014. [Google Scholar]
- Información meteo e hidro|Productos Y Servicios Inamhi. Available online: https://inamhi.wixsite.com/inamhi/novedades (accessed on 25 March 2022).
- Wunder, S. Economics of Deforestation: The Example of Ecuador; St Antony’s Series; Palgrave Macmillan: London, UK, 2000; ISBN 978-0-333-73146-8. [Google Scholar]
- Dodson, C.H.; Gentry, A.H. Biological Extinction in Western Ecuador. Ann. Mo. Bot. Gard. 1991, 78, 273. [Google Scholar] [CrossRef]
- Lippi, R.D. Paleotopography and Phosphate Analysis of a Buried Jungle Site in Ecuador. J. Field Archaeol. 1988, 15, 85–97. [Google Scholar] [CrossRef]
- Lippi, R.; Gudiño, A. Inkas and Yumbos at Palmitopamba in Northwestern Ecuador. In Distant Provinces in the Inka Empire; University of Iowa Press: Iowa City, IA, USA, 2010; pp. 260–278. [Google Scholar]
- Watling, J.; Iriarte, J.; Mayle, F.E.; Schaan, D.; Pessenda, L.C.R.; Loader, N.J.; Street-Perrott, F.A.; Dickau, R.E.; Damasceno, A.; Ranzi, A. Impact of Pre-Columbian “Geoglyph” Builders on Amazonian Forests. Proc. Natl. Acad. Sci. USA 2017, 114, 1868–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goosem, S.P.; Tucker, N.I.J.; Queensland; Wet Tropics Management Authority. Repairing the Rainforest; Wet Tropics Management Authority: Cairns, Australia, 2013; ISBN 978-1-921591-66-2.
- Espírito-Santo, F.D.; Keller, M.M.; Linder, E.; Oliveira Junior, R.C.; Pereira, C.; Oliveira, C.G. Gap Formation and Carbon Cycling in the Brazilian Amazon: Measurement Using High-Resolution Optical Remote Sensing and Studies in Large Forest Plots. Plant Ecol. Divers. 2014, 7, 305–318. [Google Scholar] [CrossRef]
- Franklin, J.F.; Spies, T.A.; Pelt, R.V.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and Structural Development of Natural Forest Ecosystems with Silvicultural Implications, Using Douglas-Fir Forests as an Example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Hunter, M.O.; Keller, M.; Morton, D.; Cook, B.; Lefsky, M.; Ducey, M.; Saleska, S.; de Oliveira, R.C., Jr.; Schietti, J. Structural Dynamics of Tropical Moist Forest Gaps. PLoS ONE 2015, 10, e0132144. [Google Scholar] [CrossRef] [PubMed]
- Brokaw, N.Y.L. The Definition of Treefall Gap and Its Effect on Measures of Forest Dynamics. Biotropica 1982, 14, 158–160. [Google Scholar] [CrossRef]
- Crausbay, S.D.; Martin, P.H. Natural Disturbance, Vegetation Patterns and Ecological Dynamics in Tropical Montane Forests. J. Trop. Ecol. 2016, 32, 384–403. [Google Scholar] [CrossRef]
- Persson, C.; Eriksson, R.; Ståhl, B.; Romoleroux, K. (Eds.) Flora of Ecuador|University of Gothenburg; University of Gothenburg: Gothenburg, Sweden, 2011. [Google Scholar]
- Python Core Team Python Software Foundation. Available online: https://www.python.org/ (accessed on 27 January 2018).
- McKinney, W. Data Structures for Statistical Computing in Python. In Proceedings of the 9th Python in Science Conference, Stéfan van der Walt, Jarrod Millman, Austin, TX, USA, 28 June–3 July 2010; pp. 51–56. [Google Scholar]
- Reback, J.; McKinney, W.; jbrockmendel; den Bossche, J.V.; Augspurger, T.; Cloud, P.; gfyoung; Sinhrks; Hawkins, S.; Klein, A.; et al. Pandas 1.1.3. Zenodo, Online Software Record. 2020. Available online: https://zenodo.org/record/4067057#.YpYM9FRByUk (accessed on 5 October 2020).
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; DPLR: Vienna, Austria, 2020. [Google Scholar]
- Salvatier, J.; Wiecki, T.V.; Fonnesbeck, C. Probabilistic Programming in Python Using PyMC3. PeerJ Comput. Sci. 2016, 2, e55. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.M.; et al. Vegan: Community Ecology Package. R Package Version 2.4-3; The Comprehensive R Archive Network: Online, 2017. [Google Scholar]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and Estimators Linking Individual-Based and Sample-Based Rarefaction, Extrapolation and Comparison of Assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Legendre, L. Developments in Environmental Modelling. In Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, ISBN 978-0-444-53868-0. [Google Scholar]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Carroll, C.; Hartikainen, A.; Martin, O. ArviZ a Unified Library for Exploratory Analysis of Bayesian Models in Python. J. Open Source Softw. 2019, 4, 1143. [Google Scholar] [CrossRef]
- Ashour, S.K.; Abdel-hameed, M.A. Approximate Skew Normal Distribution. J. Adv. Res. 2010, 1, 341–350. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, A.; Leonard, T. Bayes Estimation Subject to Uncertainty About Parameter Constraints. Biometrika 1976, 63, 201–203. [Google Scholar] [CrossRef]
- Tachikawa, T.; Kaku, M.; Iwasaki, A.; Gesch, D.B.; Oimoen, M.J.; Zhang, Z.; Danielson, J.J.; Krieger, T.; Curtis, B.; Haase, J.; et al. ASTER Global Digital Elevation Model Version 2—Summary of Validation Results, 2nd ed.; NASA: Washington, DC, USA, 2011; p. 27.
- Bartos, M. Pysheds. Software Package 2021. Available online: https://github.com/mdbartos/pysheds (accessed on 1 October 2020).
- Dijkstra, E.W. A Note on Two Problems in Connexion with Graphs. Numer. Math. 1959, 1, 269–271. [Google Scholar] [CrossRef] [Green Version]
- Hagberg, A.; Swart, P.; Chult, D.S. Exploring Network Structure, Dynamics, and Function Using Networkx; Los Alamos National Lab.: Los Alamos, NM, USA, 2008.
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving Indicator Species Analysis by Combining Groups of Sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Dray, S.; Pélissier, R.; Couteron, P.; Fortin, M.-J.; Legendre, P.; Peres-Neto, P.R.; Bellier, E.; Bivand, R.; Blanchet, F.G.; Cáceres, M.D.; et al. Community Ecology in the Age of Multivariate Multiscale Spatial Analysis. Ecol. Monogr. 2012, 82, 257–275. [Google Scholar] [CrossRef]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY Plant Trait Database—Enhanced Coverage and Open Access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [Green Version]
- Parr, C.S.; Wilson, N.; Leary, P.; Schulz, K.; Lans, K.; Walley, L.; Hammock, J.; Goddard, A.; Rice, J.; Studer, M.; et al. The Encyclopedia of Life v2: Providing Global Access to Knowledge About Life on Earth. Biodivers. Data J. 2014, 2, e1079. [Google Scholar] [CrossRef] [Green Version]
- Gentry, A.H. A Field Guide to the Families and Genera of Woody Plants of Northwest South America (Colombia, Ecuador, Peru), with Supplementary Notes on Herbaceous Taxa; The University of Chicago Press: Chicago, IL, USA, 1993; ISBN 0-226-28944-3. [Google Scholar]
- GDAL/OGR contributors. GDAL/OGR Geospatial Data Abstraction Software Library; Open Source Geospatial Foundation: Beaverton, OR, USA, 2021. [Google Scholar]
- Grubb, P.J. Control of Forest Growth and Distribution on Wet Tropical Mountains: With Special Reference to Mineral Nutrition. Annu. Rev. Ecol. Syst. 1977, 8, 83–107. [Google Scholar] [CrossRef]
- Leigh, E.G. Structure and Climate in Tropical Rain Forest. Annu. Rev. Ecol. Syst. 1975, 6, 67–86. [Google Scholar] [CrossRef]
- Sohng, J.; Singhakumara, B.M.P.; Ashton, M.S. Effects on Soil Chemistry of Tropical Deforestation for Agriculture and Subsequent Reforestation with Special Reference to Changes in Carbon and Nitrogen. For. Ecol. Manag. 2017, 389, 331–340. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, Q.; Hui, D.; Wu, J.; Xiong, X.; Zhao, J.; Zhao, M.; Chu, G.; Zhou, G.; Zhang, D. Recovery in Soil Carbon Stock but Reduction in Carbon Stabilization after 56-Year Forest Restoration in Degraded Tropical Lands. For. Ecol. Manag. 2019, 441, 1–8. [Google Scholar] [CrossRef]
- Hartmann, M.; Howes, C.G.; VanInsberghe, D.; Yu, H.; Bachar, D.; Christen, R.; Henrik Nilsson, R.; Hallam, S.J.; Mohn, W.W. Significant and Persistent Impact of Timber Harvesting on Soil Microbial Communities in Northern Coniferous Forests. ISME J. 2012, 6, 2199–2218. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Niklaus, P.A.; Zimmermann, S.; Schmutz, S.; Kremer, J.; Abarenkov, K.; Lüscher, P.; Widmer, F.; Frey, B. Resistance and Resilience of the Forest Soil Microbiome to Logging-Associated Compaction. ISME J. 2014, 8, 226–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; He, N.; Xu, L.; Zhou, X. Microbial Properties Regulate Spatial Variation in the Differences in Heterotrophic Respiration and Its Temperature Sensitivity between Primary and Secondary Forests from Tropical to Cold-Temperate Zones. Agric. For. Meteorol. 2018, 262, 81–88. [Google Scholar] [CrossRef]
- Bart, D.; Davenport, T. The Influence of Legacy Impacted Seed Banks on Vegetation Recovery in a Post-Agricultural Fen Complex. Wetl. Ecol Manag. 2015, 23, 405–418. [Google Scholar] [CrossRef]
- Young, K.R.; Ewel, J.J.; Brown, B.J. Seed Dynamics during Forest Succession in Costa Rica. Vegetatio 1987, 71, 157–173. [Google Scholar] [CrossRef]
- Myster, R.W. Early Successional Pattern and Process after Sugarcane, Banana, and Pasture Cultivation in Ecuador. N. Z. J. Bot. 2007, 45, 101–110. [Google Scholar] [CrossRef]
- Myster, R.W. Seed Predation, Pathogens and Germination in Primary vs. Secondary Cloud Forest at Maquipucuna Reserve, Ecuador. J. Trop. Ecol. 2015, 31, 375–378. [Google Scholar] [CrossRef]
- Fragaszy, D.M.; Visalberghi, E.; Fedigan, L.M. The Complete Capuchin: The Biology of the Genus Cebus; Cambridge University Press: London, UK, 2004; ISBN 0-521-66768-2. [Google Scholar]
- Russo, S.E.; Campbell, C.J.; Dew, J.L.; Stevenson, P.R.; Suarez, S.A. A Multi-Forest Comparison of Dietary Preferences and Seed Dispersal by Ateles spp. Int. J. Primatol. 2005, 26, 1017–1037. [Google Scholar] [CrossRef] [Green Version]
- Chanderbali, A. Flora Neotropica; publ. for Organization for flora neotropica. In Lauraceae; The New York Botanical Garden: New York, NY, USA, 2004; ISBN 978-0-89327-454-2. [Google Scholar]
- Bufalo, F.S.; Galetti, M.; Culot, L. Seed Dispersal by Primates and Implications for the Conservation of a Biodiversity Hotspot, the Atlantic Forest of South America. Int. J. Primatol. 2016, 37, 333–349. [Google Scholar] [CrossRef] [Green Version]
- Morelos-Juárez, C.; Tapia, A.; Conde, G.; Peck, M. Diet of the Critically Endangered Brown-Headed Spider Monkey (Ateles Fusciceps Fusciceps) in the Ecuadorian Chocó: Conflict between Primates and Loggers over Fruiting Tree Species; PeerJ Inc.: Corte Madera, CA, USA, 2015. [Google Scholar]
- Howe, H.F.; Smallwood, J. Ecology of Seed Dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Cubiña, A.; Aide, T.M. The Effect of Distance from Forest Edge on Seed Rain and Soil Seed Bank in a Tropical Pasture. Biotropica 2001, 33, 260–267. [Google Scholar] [CrossRef]
- Holl, K.D. Factors Limiting Tropical Rain Forest Regeneration in Abandoned Pasture: Seed Rain, Seed Germination, Microclimate, and Soil1. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Mariscal, A.; Tigabu, M.; Savadogo, P.; Odén, P.C. Regeneration Status and Role of Traditional Ecological Knowledge for Cloud Forest Ecosystem Restoration in Ecuador. Forests 2022, 13, 92. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Planchuelo, G.; Kowarik, I.; von der Lippe, M. Endangered Plants in Novel Urban Ecosystems Are Filtered by Strategy Type and Dispersal Syndrome, Not by Spatial Dependence on Natural Remnants. Front. Ecol. Evol. 2020, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Chapman, H.; Cordeiro, N.J.; Dutton, P.; Wenny, D.; Kitamura, S.; Kaplin, B.; Melo, F.P.L.; Lawes, M.J. Seed-Dispersal Ecology of Tropical Montane Forests. J. Trop. Ecol. 2016, 32, 437–454. [Google Scholar] [CrossRef]
- Nunez-Iturri, G.; Olsson, O.; Howe, H.F. Hunting Reduces Recruitment of Primate-Dispersed Trees in Amazonian Peru. Biol. Conserv. 2008, 141, 1536–1546. [Google Scholar] [CrossRef]
- van der Werff, H. A New Species of Caryodaphnopsis (Lauraceae) from Peru. Syst. Bot. 1986, 11, 415–418. [Google Scholar] [CrossRef]
- Romo, M. Seasonal Variation in Fruit Consumption and Seed Dispersal by Canopy Bats (Artibeus spp.) in a Lowland Forest in Peru. Vida Silv. Neotrop. 1996, 5, 110–119. [Google Scholar]
- Acero-Murcia, A.; Almario, L.J.; García, J.; Defler, T.R.; López, R. Diet of the Caquetá Titi (Plecturocebus caquetensis) in a Disturbed Forest Fragment in Caquetá, Colombia. Primate Conserv. 2018, 32, 1–7. [Google Scholar]
- Knogge, C.; Heymann, E.W. Seed Dispersal by Sympatric Tamarins, Saguinus Mystax and Saguinus Fuscicollis: Diversity and Characteristics of Plant Species. Folia Primatol. 2003, 74, 33–47. [Google Scholar] [CrossRef]
- Giraldo, P.; Gómez-Posada, C.; Martínez, J.; Kattan, G. Resource Use and Seed Dispersal by Red Howler Monkeys (Alouatta seniculus) in a Colombian Andean Forest. Neotrop. Primates 2007, 14, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Willson, M.F.; Crome, F.H.J. Patterns of Seed Rain at the Edge of a Tropical Queensland Rain Forest. J. Trop. Ecol. 1989, 5, 301–308. [Google Scholar] [CrossRef]
- Cramer, J.M.; Mesquita, R.C.G.; Bruce Williamson, G. Forest Fragmentation Differentially Affects Seed Dispersal of Large and Small-Seeded Tropical Trees. Biol. Conserv. 2007, 137, 415–423. [Google Scholar] [CrossRef]
- Peck, M.; Thorn, J.; Mariscal, A.; Baird, A.; Tirira, D.; Kniveton, D. Focusing Conservation Efforts for the Critically Endangered Brown-Headed Spider Monkey (Ateles fusciceps) Using Remote Sensing, Modeling, and Playback Survey Methods. Int. J. Primatol. 2011, 32, 134–148. [Google Scholar] [CrossRef]
- Peres, C.A.; Emilio, T.; Schietti, J.; Desmoulière, S.J.M.; Levi, T. Dispersal Limitation Induces Long-Term Biomass Collapse in Overhunted Amazonian Forests. Proc. Natl. Acad. Sci. USA 2016, 113, 892–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calle-Rendón, B.R.; Peck, M.; Bennett, S.E.; Morelos-Juarez, C.; Alfonso, F. Comparison of Forest Regeneration in Two Sites with Different Primate Abundances in Northwestern Ecuador. Rev. De Biol. Trop. 2016, 64, 493–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morante-Filho, J.C.; Arroyo-Rodríguez, V.; Pessoa, M.D.; Cazetta, E.; Faria, D. Direct and Cascading Effects of Landscape Structure on Tropical Forest and Non-Forest Frugivorous Birds. Ecol. Appl. 2018, 28, 2024–2032. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.B.; Bovo, A.A.A.; Alexandrino, E.R.; Oliveira, V.C.; Pizo, M.A.; Ferraz, K.M.P.M.B. Fruit Availability at the Individual and Local Levels Influences Fruit Removal in Cecropia pachystachya. Braz. J. Biol. 2018, 79, 758–759. [Google Scholar] [CrossRef] [PubMed]
- Skutch, A.F. The Most Hospitable Tree. Sci. Mon. 1945, 60, 5–17. [Google Scholar]
- Baldeck, C.A.; Harms, K.E.; Yavitt, J.B.; John, R.; Turner, B.L.; Valencia, R.; Navarrete, H.; Bunyavejchewin, S.; Kiratiprayoon, S.; Yaacob, A. Habitat Filtering across Tree Life Stages in Tropical Forest Communities. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130548. [Google Scholar] [CrossRef] [Green Version]
- Ledo, A.; Cayuela, L.; Manso, R.; Condés, S. Recruitment Patterns and Potential Mechanisms of Community Assembly in an Andean Cloud Forest. J. Veg. Sci. 2015, 26, 876–888. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest Disturbances under Climate Change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative Stable States in Ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Clerici, N.; Cote-Navarro, F.; Escobedo, F.J.; Rubiano, K.; Villegas, J.C. Spatio-Temporal and Cumulative Effects of Land Use-Land Cover and Climate Change on Two Ecosystem Services in the Colombian Andes. Sci. Total Environ. 2019, 685, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J.; Silman, M.R.; Bush, M.B.; Farfan, W.; Cabrera, K.G.; Malhi, Y.; Meir, P.; Revilla, N.S.; Quisiyupanqui, M.N.R.; Saatchi, S. Upslope Migration of Andean Trees. J. Biogeogr. 2011, 38, 783–791. [Google Scholar] [CrossRef]
- Rehm, E.M.; Feeley, K.J. The Inability of Tropical Cloud Forest Species to Invade Grasslands above Treeline during Climate Change: Potential Explanations and Consequences. Ecography 2015, 38, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Stork, N.E. Re-Assessing Current Extinction Rates. Biodivers. Conserv. 2010, 19, 357–371. [Google Scholar] [CrossRef]
- Tovar, C.; Arnillas, C.A.; Cuesta, F.; Buytaert, W. Diverging Responses of Tropical Andean Biomes under Future Climate Conditions. PLoS ONE 2013, 8, e63634. [Google Scholar] [CrossRef] [Green Version]
- Myers, N. Threatened Biotas:” Hot Spots” in Tropical Forests. Environmentalist 1988, 8, 187–208. [Google Scholar] [CrossRef]
- Mosandl, R.; Günter, S.; Stimm, B.; Weber, M. Ecuador Suffers the Highest Deforestation Rate in South America. In Gradients in a Tropical Mountain Ecosystem of Ecuador; Beck, E., Bendix, J., Kottke, I., Makeschin, F., Mosandl, R., Eds.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2008; pp. 37–40. ISBN 978-3-540-73526-7. [Google Scholar]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; de la Cruz, M. Deforestation and Forest Fragmentation in South Ecuador since the 1970s—Losing a Hotspot of Biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef] [Green Version]
- Guayasamin, J.M.; Vandegrift, R.; Policha, T.; Encalada, A.C.; Greene, N.; Ríos-Touma, B.; Endara, L.; Cárdenas, R.E.; Larreátegui, F.; Baquero, L.; et al. Tipping Point Towards Biodiversity Conservation? Local and Global Consequences of the Application of ‘Rights of Nature’ by Ecuador. Soc. Sci. 2021. [Google Scholar] [CrossRef]
- Vallejo Galárraga, M.C.; Freslon, W.S. Ecuador: Mineral Policy. In Encyclopedia of Mineral and Energy Policy; Tiess, G., Majumder, T., Cameron, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–8. ISBN 978-3-642-40871-7. [Google Scholar]
- Filotas, E.; Parrott, L.; Burton, P.J.; Chazdon, R.L.; Coates, K.D.; Coll, L.; Haeussler, S.; Martin, K.; Nocentini, S.; Puettmann, K.J.; et al. Viewing Forests through the Lens of Complex Systems Science. Ecosphere 2014, 5, art1. [Google Scholar] [CrossRef]
- Messier, C.; Puettmann, K.; Chazdon, R.; Andersson, K.P.; Angers, V.A.; Brotons, L.; Filotas, E.; Tittler, R.; Parrott, L.; Levin, S.A. From Management to Stewardship: Viewing Forests As Complex Adaptive Systems in an Uncertain World. Conserv. Lett. 2015, 8, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Loehle, C. Challenges of Ecological Complexity. Ecol. Complex. 2004, 1, 3–6. [Google Scholar]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and Woodland Stand Structural Complexity: Its Definition and Measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Ripple, W.J.; Estes, J.A.; Schmitz, O.J.; Constant, V.; Kaylor, M.J.; Lenz, A.; Motley, J.L.; Self, K.E.; Taylor, D.S.; Wolf, C. What Is a Trophic Cascade? Trends Ecol. Evol. 2016, 31, 842–849. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001; Volume 1, ISBN 0-691-08836-5. [Google Scholar]
- Ross, K.A.; Fox, B.J.; Fox, M.D. Changes to Plant Species Richness in Forest Fragments: Fragment Age, Disturbance and Fire History May Be as Important as Area. J. Biogeogr. 2002, 29, 749–765. [Google Scholar] [CrossRef]
- Michaels, T.K.; Eppinga, M.B.; Bever, J.D. A Nucleation Framework for Transition between Alternate States: Short-Circuiting Barriers to Ecosystem Recovery. Ecology 2020, 101, e03099. [Google Scholar] [CrossRef] [PubMed]
- Fernández, N.; Navarro, L.M.; Pereira, H.M. Rewilding: A Call for Boosting Ecological Complexity in Conservation. Conserv. Lett. 2017, 10, 276–278. [Google Scholar] [CrossRef]
- Balmford, A.; Bruner, A.; Cooper, P.; Costanza, R.; Farber, S.; Green, R.E.; Jenkins, M.; Jefferiss, P.; Jessamy, V.; Madden, J.; et al. Economic Reasons for Conserving Wild Nature. Science 2002, 297, 950–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocian, M.; Batker, D.; Harrison-Cox, J. An Ecological Study of Ecuador’s Intag Region: The Environmental Impacts and Potential Rewards of Mining; Earth Economics: Tacoma, WA, USA, 2011. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Size | Species Observed | Chao | Chao.Se | Jack1 | Jack1.Se | Jack2 | Boot | Boot.Se | |
|---|---|---|---|---|---|---|---|---|---|
| Survey Plots | 61 | 343 | 516.4 | 39.2 | 483.7 | 21.6 | 566.8 | 404.4 | 10.5 |
| Permanent Plot (Trees) | 121 | 43 | 65.4 | 13.8 | 61.8 | 4.3 | 72.7 | 51.4 | 2.2 |
| Watersheds Crossed | Sample Size | Mean BC | Std. dev BC |
|---|---|---|---|
| 0 | 215 | 0.820 | 0.114 |
| 1 | 630 | 0.880 | 0.089 |
| 2 | 559 | 0.872 | 0.085 |
| 3 | 346 | 0.891 | 0.079 |
| 4 | 80 | 0.881 | 0.096 |
| Group1 | Group2 | Meandiff | p-Adj | Lower | Upper | Reject |
|---|---|---|---|---|---|---|
| 0 | 1 | 0.0601 | 0.001 | 0.0408 | 0.0795 | TRUE |
| 0 | 2 | 0.0516 | 0.001 | 0.0319 | 0.0713 | TRUE |
| 0 | 3 | 0.0708 | 0.001 | 0.0495 | 0.0921 | TRUE |
| 0 | 4 | 0.0609 | 0.001 | 0.0287 | 0.093 | TRUE |
| 1 | 2 | −0.0085 | 0.4759 | −0.0228 | 0.0057 | FALSE |
| 1 | 3 | 0.0106 | 0.393 | −0.0058 | 0.0271 | FALSE |
| 1 | 4 | 0.0007 | 0.9 | −0.0284 | 0.0299 | FALSE |
| 2 | 3 | 0.0192 | 0.0158 | 0.0024 | 0.036 | TRUE |
| 2 | 4 | 0.0093 | 0.9 | −0.0201 | 0.0386 | FALSE |
| 3 | 4 | −0.0099 | 0.9 | −0.0404 | 0.0205 | FALSE |
| Indicator Species by Land-Use-History/Habitat-Type | |||
| SGroup BC | |||
| Indicator Species | Rpb.G | p-Value | Sig Code |
| Pseudolmedia rigida (Moraceae) | 0.423 | 0.0317 | * |
| Ficus subandina (Moraceae) | 0.406 | 0.049 | * |
| Group BS | |||
| Rpb.G | p-Value | Sig Code | |
| Psychotria paeonia (Rubiaceae) | 0.478 | 0.0148 | * |
| Persea pseudofasciculata (Lauraceae) | 0.432 | 0.0351 | * |
| Myrcia aff. aliena (Myrtaceae) | 0.425 | 0.0377 | * |
| Group CLB | |||
| Rpb.G | p-Value | Sig Code | |
| Endlicheria cf. chalisea (Lauraceae) | 0.431 | 0.0268 | * |
| Group RCA | |||
| Rpb.G | p-Value | Sig Code | |
| Cordia colombiana (Boraginaceae) | 0.894 | 0.0001 | *** |
| Saurauia sp. 1 (Actinidaceae) | 0.694 | 0.0003 | *** |
| Miconia aff. brevitheca (Melastomataceae) | 0.683 | 0.0004 | *** |
| Ficus caldasiana (Moraceae) | 0.667 | 0.0033 | ** |
| Turpinia occidentalis (Staphyleaceae) | 0.663 | 0.0006 | *** |
| Clarisia biflora (Moraceae) | 0.645 | 0.0022 | ** |
| Nectandra membranacea (Lauraceae) | 0.61 | 0.0027 | ** |
| Aegiphila alba (Verbenaceae) | 0.594 | 0.002 | ** |
| Ficus andicola (Moraceae) | 0.539 | 0.0191 | * |
| Caryodaphnopsis theobromifolia (Lauraceae) | 0.535 | 0.0099 | ** |
| Nectandra aff. membranacea (Lauraceae) | 0.513 | 0.0106 | * |
| Group RG | |||
| Rpb.G | p-Value | Sig Code | |
| Cecropia andina (Cecropiaceae) | 0.662 | 0.0018 | ** |
| Cecropia ficifolia (Cecropiaceae) | 0.58 | 0.0035 | ** |
| Cecropia sp. 2 (Cecropiaceae) | 0.427 | 0.031 | * |
| Meriania tomentosa (Melastomataceae) | 0.415 | 0.0426 | * |
| Group BC + BS | |||
| Rpb.G | p-Value | Sig Code | |
| Aniba aff. hostmanniana (Lauraceae) | 0.411 | 0.0471 | * |
| Group RCA + RG | |||
| Rpb.G | p-Value | Sig Code | |
| Solanum lepidotum (Solanaceae) | 0.529 | 0.006 | ** |
| Cestrum megalophyllum (Solanaceae) | 0.423 | 0.0268 | * |
| Group BC + BS + CLB | |||
| Rpb.G | p-Value | Sig Code | |
| Alsophila erinacea (Cyatheaceae) | 0.42 | 0.0423 | * |
| Indicator Species by Current Ecological State (Forest Type) | |||
| Group I | |||
| Rpb.G | p-Value | Sig Code | |
| Cordia colombiana (Boraginaceae) | 0.661 | 0.0001 | *** |
| Meriania tomentosa (Melastomataceae) | 0.641 | 0.0001 | *** |
| Saurauia sp. 1 (Actinidaceae) | 0.545 | 0.0003 | *** |
| Cyathea halonata (Cyatheaceae) | 0.458 | 0.0033 | ** |
| Senna dariensis (Fab. Caesalpiniaceae) | 0.449 | 0.0046 | ** |
| Ficus caldasiana (Moraceae) | 0.42 | 0.0206 | * |
| Leandra subseriata (Melastomataceae) | 0.37 | 0.015 | * |
| Miconia aff. brevitheca (Melastomataceae) | 0.368 | 0.027 | * |
| Piper fuliginosum (Piperaceae) | 0.356 | 0.05 | * |
| Turpinia occidentalis (Staphyleaceae) | 0.346 | 0.0426 | * |
| Group II | |||
| Rpb.G | p-Value | Sig Code | |
| Cecropia andina (Cecropiaceae) | 0.811 | 0.0001 | *** |
| Cecropia sp. 2 (Cecropiaceae) | 0.5 | 0.0009 | *** |
| Melastomataceae sp. 1 | 0.468 | 0.0051 | ** |
| Dussia lehmannii (Fab. Faboideae) | 0.334 | 0.0444 | * |
| Group III | |||
| Rpb.G | p-Value | Sig Code | |
| Otoba gordoniifolia (Myristicaceae) | 0.524 | 0.0007 | *** |
| Alsophila erinacea (Cyatheaceae) | 0.515 | 0.0015 | ** |
| Persea aff. pseudofasciculata (Lauraceae) | 0.494 | 0.0022 | ** |
| Vismia lauriformis (Clusiaceae) | 0.44 | 0.0065 | ** |
| Conostegia aff. centronioides (Melastomataceae) | 0.418 | 0.0083 | ** |
| Wettinia aff. oxycarpa (Arecaceae) | 0.403 | 0.0115 | * |
| Persea pseudofasciculata (Lauraceae) | 0.386 | 0.0217 | * |
| Psychotria paeonia (Rubiaceae) | 0.373 | 0.0264 | * |
| Ficus dulciaria (Rubiaceae) | 0.354 | 0.03 | * |
| Group IV | |||
| Rpb.G | p-Value | Sig Code | |
| Dacryodes cupularis (Burseraceae) | 0.754 | 0.0001 | *** |
| Protium ecuadorense (Burseraceae) | 0.638 | 0.0001 | *** |
| Garcinia macrophylla (Clusiaceae) | 0.613 | 0.0001 | *** |
| .Beilschmiedia aff. costaricensis (Lauraceae) | 0.448 | 0.0018 | ** |
| Conostegia superba (Melastomataceae) | 0.444 | 0.0024 | ** |
| Ocotea stenoneura (Lauraceae) | 0.369 | 0.0454 | * |
| Gustavia dodsonii (Lecythidaceae) | 0.362 | 0.029 | * |
| Styrax weberbaueri (Styracaceae) | 0.361 | 0.0408 | * |
| Pseudolmedia rigida (Moraceae) | 0.355 | 0.0263 | * |
| Group I + II | |||
| Rpb.G | p-Value | Sig Code | |
| Cecropia ficifolia (Cecropiaceae) | 0.52 | 0.0005 | *** |
| Solanum lepidotum (Solanaceae) | 0.447 | 0.0055 | ** |
| Cecropia reticulata (Cecropiaceae) | 0.371 | 0.0244 | * |
| Aegiphila alba (Verbenaceae) | 0.348 | 0.0465 | * |
| Urera caracasana (Urticaceae) | 0.341 | 0.037 | * |
| Group I + IV | |||
| Rpb.G | p-Value | Sig Code | |
| Caryodaphnopsis theobromifolia (Lauraceae) | 0.416 | 0.0092 | ** |
| Clarisia biflora (Moraceae) | 0.403 | 0.0157 | * |
| Group II + III | |||
| Rpb.G | p-Value | Sig Code | |
| Guatteria megalophylla (Annonaceae) | 0.411 | 0.0133 | * |
| Ficus cuatrecasana (Moraceae) | 0.364 | 0.0299 | * |
| Group III + IV | |||
| Rpb.G | p-Value | Sig Code | |
| Aniba aff. hostmanniana (Lauraceae) | 0.401 | 0.0138 | * |
| Capparis sp. (Capparaceae) | 0.385 | 0.0267 | * |
| Hieronyma asperifolia (Euphorbiaceae) | 0.383 | 0.0226 | * |
| Eschweilera integrifolia (Lecythidaceae) | 0.379 | 0.0199 | * |
| Ocotea insularis (Lauraceae) | 0.366 | 0.0295 | * |
| Faramea oblongifolia (Rubiaceae) | 0.362 | 0.0291 | * |
| Helicostylis tovarensis (Moraceae) | 0.346 | 0.0444 | * |
| Slope | Dem | Aspect | Exposure (Eastern) | Exposure (Northen) | Tostream | BC | BS | CLB | RCA | RG | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MEM1 | −0.403 | 0.287 | |||||||||
| MEM2 | −0.344 | −0.636 | 0.308 | ||||||||
| MEM3 | |||||||||||
| MEM4 | 0.349 | ||||||||||
| MEM5 | −0.503 | ||||||||||
| MEM6 | 0.306 | ||||||||||
| MEM7 | 0.385 | ||||||||||
| MEM8 | 0.404 | 0.460 | |||||||||
| MEM9 | −0.451 | ||||||||||
| MEM10 | −0.340 | ||||||||||
| MEM11 | 0.282 | ||||||||||
| MEM12 | −0.410 | ||||||||||
| MEM13 | 0.345 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariscal, A.; Thomas, D.C.; Haffenden, A.; Manobanda, R.; Defas, W.; Angel Chinchero, M.; Simba Larco, J.D.; Jaramillo, E.; Roy, B.A.; Peck, M. Evidence for Alternate Stable States in an Ecuadorian Andean Cloud Forest. Forests 2022, 13, 875. https://doi.org/10.3390/f13060875
Mariscal A, Thomas DC, Haffenden A, Manobanda R, Defas W, Angel Chinchero M, Simba Larco JD, Jaramillo E, Roy BA, Peck M. Evidence for Alternate Stable States in an Ecuadorian Andean Cloud Forest. Forests. 2022; 13(6):875. https://doi.org/10.3390/f13060875
Chicago/Turabian StyleMariscal, Ana, Daniel Churchill Thomas, Austin Haffenden, Rocío Manobanda, William Defas, Miguel Angel Chinchero, José Danilo Simba Larco, Edison Jaramillo, Bitty A. Roy, and Mika Peck. 2022. "Evidence for Alternate Stable States in an Ecuadorian Andean Cloud Forest" Forests 13, no. 6: 875. https://doi.org/10.3390/f13060875