
Allometric Equation for Aboveground Biomass Estimation of Mixed Mature Mangrove Forest

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Sample Preparation
2.3. Approach in Biomass Estimation
- M = Aboveground biomass
- D = Diameter at breast height
- H = Stem height
- a and b = constant
- e = error term
- ρ = wood density
- Theoretical = Known value
- Experimental = Calculated value
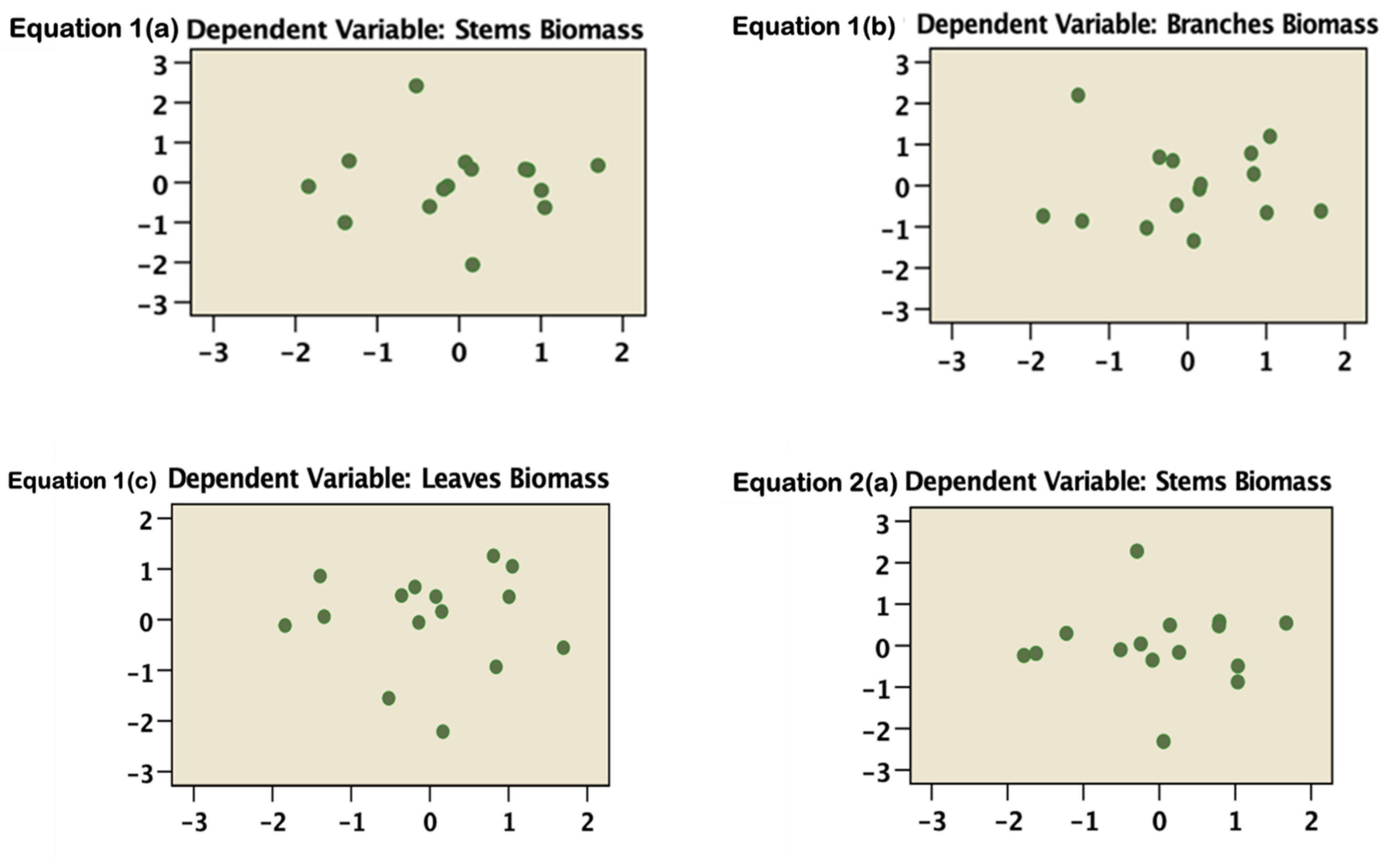
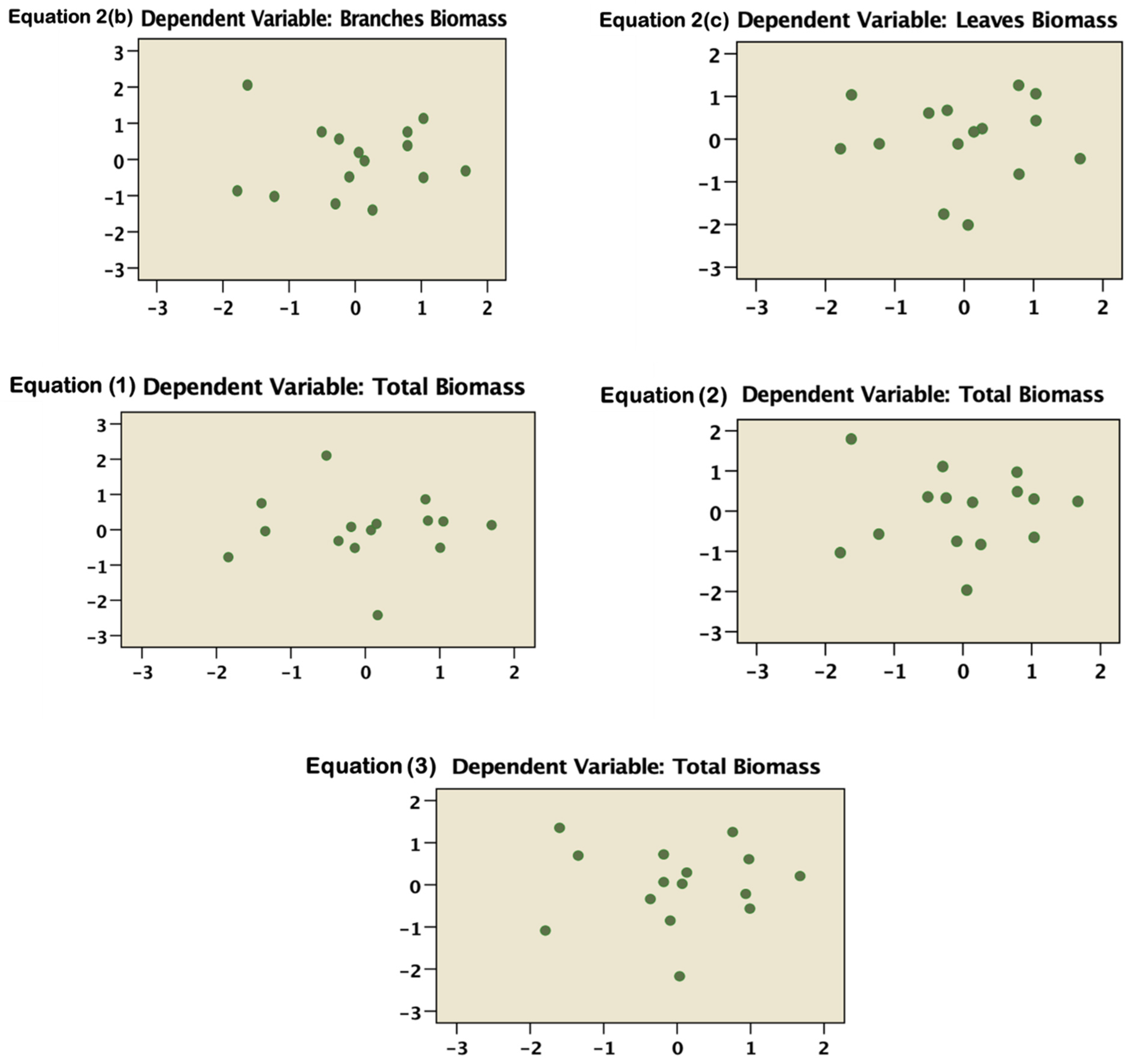
2.4. Statistical Analysis
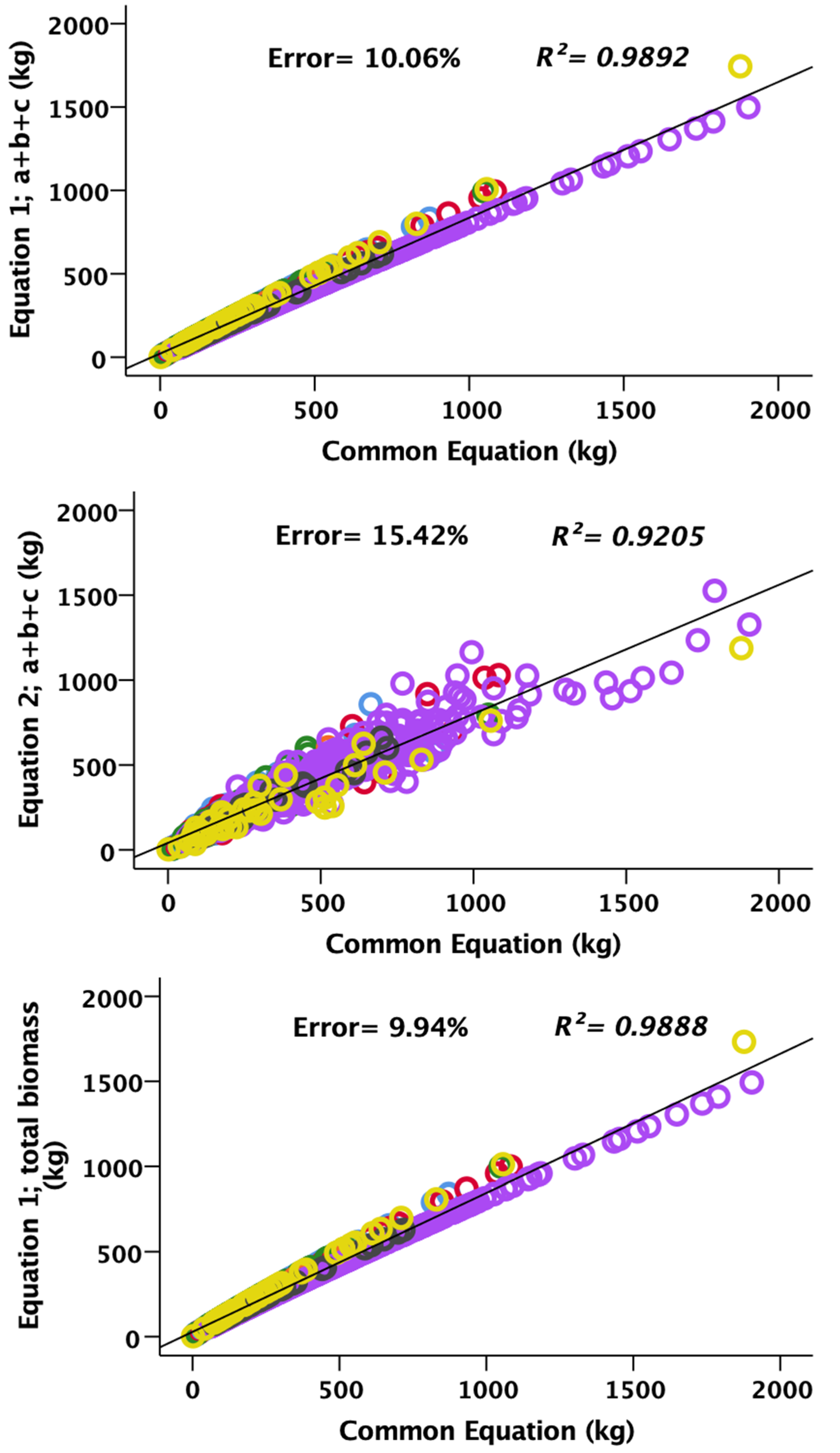
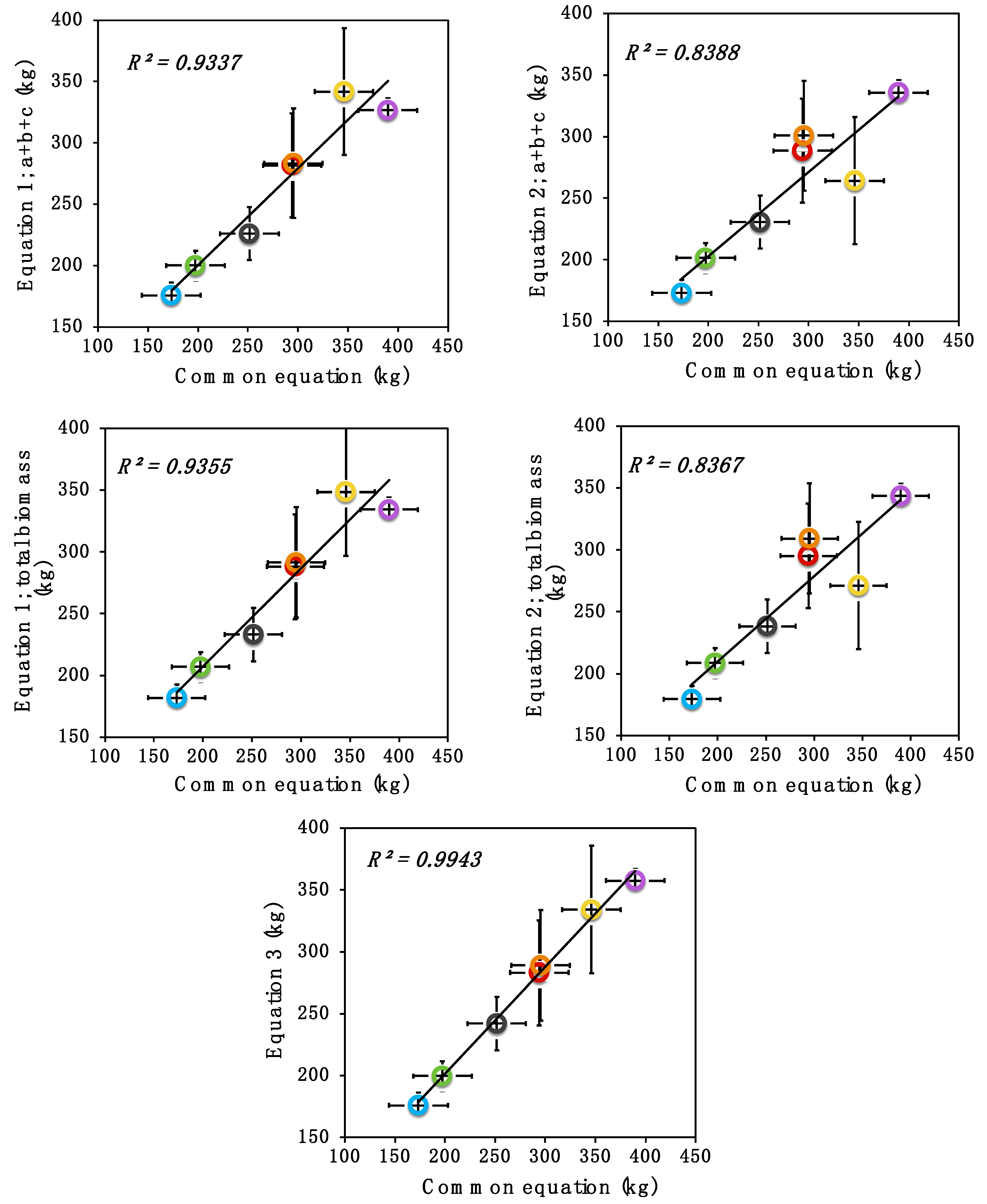
3. Results and Discussion
3.1. Allometric Equations Derived from 15 Destructive Trees
3.2. Allometric Equations Applied to 1000 Standing Trees
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Group | Equation | R2 | N | Data Origin | D Max (cm) | Source |
|---|---|---|---|---|---|---|
| General equation | B = ρ × exp [−1.349 + 1.980 × ln(D) + 0.207 × (ln(D))2 − 0.0281 × (ln(D))3] | unknown | 84 | Americas | 42.0 | Chave et al. [41] |
| General equation | B = 0.168 × ρ × (D)2.471 | 0.99 | 84 | Americas | 42.0 | Chave et al. [41]; Komiyama et al. [11] |
| General equation | B = 0.251 ρ (D)2.46 | 0.98 | 104 | Asia | 49.0 | Komiyama et al. [29] |
| Specific tree equations—Asia-Pacific region | ||||||
| Rhizophora apiculata | B = 0.1709D2.516 | 0.98 | 20 | Malaysia | 30.0 | Putz & Chan [27] |
| Rhizophora apiculata | B = 0.043D2.63 | 0.97 | 34 | Indonesia | 40.0 | Amira [63] |
| Rhizophora apiculata (wood mass) | Bwood = 0.0695D2.644 × ρ | 0.89 | 191 | Micronesia | 60.0 | Modified from Cole et al. [64]; Kauffman & Cole [65] |
| Xylocapus granatum | B = 0.1832D2.21 | 0.95 | 30 | Indonesia | 41.0 | Tarlan [66] |
Appendix B

References
- Ball, M.C. Ecophysiology of mangroves. Trees 1988, 2, 129–142. [Google Scholar] [CrossRef]
- Twilley, R.R. Coupling of mangroves to the productivity of estuarine and coastal waters. In Coastal-Offshore Ecosystem Interactions: Lecture Notes on Coastal and Estuarine Studies; Jansson, B.O., Ed.; American Geophysical Union Publisher: Washington, DC, USA, 1988; pp. 165–202. [Google Scholar] [CrossRef]
- Baran, E.; Hambrey, J. Mangrove conservation and coastal management in Southeast Asia: What impact on fishery resources? Mar. Pollut. Bull. 1999, 37, 431–440. [Google Scholar] [CrossRef]
- Primavera, J.H. Tropical shrimp farming and its sustainability. Trop. Maric. 1998, 8, 257–289. [Google Scholar] [CrossRef]
- Aburto-Oropeza, O.; Ezcurra, E.; Danemann, G.; Valdez, V.; Murray, J.; Sala, E. Mangroves in the Gulf of California increase fishery yields. Proc. Natl. Acad. Sci. USA 2008, 105, 10456–10459. [Google Scholar] [CrossRef] [Green Version]
- Dahdouh-Guebas, F.; Mathenge, C.; Kairo, J.G.; Koedam, N. Utilization of mangrove wood products around Mida Creek (Kenya) amongst subsistence and commercial users. Econ. Bot. 2000, 54, 513–527. [Google Scholar] [CrossRef]
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, H.; Li, Y.; Xu, H.; Shen, J.; Rhome, J.; Smith, T.J. The role of mangroves in attenuating storm surges. Estuar. Coast. Shelf Sci. 2012, 102–103, 11–23. [Google Scholar] [CrossRef]
- Cahoon, D.R.; Hensel, P.; Rybczyk, J.; McKee, K.L.; Proffitt, C.E.; Perez, B.C. Mass tree mortality leads to mangrove peat collapse at Bay Islands, Honduras after Hurricane Mitch. J. Ecol. 2003, 91, 1093–1105. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castañeda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middleburg, J.J.; et al. Mangrove production and carbon sinks: A revision of global budget estimates. Glob. Biogeochem. Cycles 2008, 22, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Ministry of Natural Resources and Environment (MNRE), Malaysia. Malaysia’s Submission on Reference Levels for REDD+ Results Based Payments under UNFCCC, 2015. Available online: https://redd.unfccc.int/files/2018_frel_submission_malaysia.pdf (accessed on 7 January 2022).
- Nomura, K.; Mitchard, E.T.A.; Bowers, S.J.; Patenaude, G. Missed carbon emissions from forests: Comparing countries’ estimates submitted to UNFCCC to biophysical estimates. Environmental Res. Lett. 2019, 14, 4015. [Google Scholar] [CrossRef]
- The Economics of Ecosystem and Biodiversity, TEEB. An Interim Report; European Communities: Brussel, Belgium, 2008. [Google Scholar]
- De Groot, R.; Brander, L.; Van Der Ploeg, S.; Costanza, R.; Bernard, F.; Braat, L.; Christie, M.; Crossman, N.; Ghermandi, A.; Hein, L.; et al. Global estimates of the value of ecosystems and their services in monetary units. Ecosyst. Serv. 2012, 1, 50–61. [Google Scholar] [CrossRef]
- Zolkos, S.; Goetz, S.; Dubayah, R. A meta-analysis of terrestrial aboveground biomass estimation using Lidar remote sensing. Remote Sens. Environ. 2013, 128, 289–298. [Google Scholar] [CrossRef]
- Duncanson, L.; Dubayar, R.; Cook, B.; Rosette, J.; Parker, G. The importance of spatial detail: Assessing the utility of individual crown information and scaling approaches for Lidar-based biomass density estimation. Remote Sens. Environ. 2015, 168, 102–112. [Google Scholar] [CrossRef]
- Taylor, P.; Asner, G.; Dahlin, K.; Anderson, K.; Knapp, D.; Martin, R.; Mascaro, J.; Chazdon, R.; Cole, R.; Wanek, W. Landscape-scale controls on aboveground forest carbon stocks on the Osa Peninsula, Costa Rica. PLoS ONE 2015, 10, e0126748. [Google Scholar] [CrossRef] [Green Version]
- Fatoyinbo, T.E.; Simard, M. Height and biomass of mangroves in Africa from ICESat/ GLAS and SRTM. Int. J. Remote Sens. 2013, 34, 668–681. [Google Scholar] [CrossRef]
- Brown, S.; Gillespie, A.J.R.; Lugo, A.E. Biomass estimation methods for tropical forests with applications to forest inventory data. For. Sci. 1989, 35, 881–902. [Google Scholar] [CrossRef]
- Nelson, B.W.; Mesquita, R.; Pereira, J.L.G.; De-Souza, S.G.A.; Batista, G.T.; Couto, L.B. Allometric regressions for improved estimate of secondary forest biomass in the central Amazon. For. Ecol. Manag. 1999, 117, 149–167. [Google Scholar] [CrossRef]
- Basuki, T.M.; Van-Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the aboveground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Djomo, A.N.; Ibrahima, A.; Saborowski, J.; Gravenhorst, G. Allometric equations for biomass estimations in Cameroon and pan moist tropical equation including biomass data from Africa. For. Ecol. Manag. 2010, 260, 1873–1885. [Google Scholar] [CrossRef]
- Fayolle, A.; Doucet, J.L.; Gillet, J.F.; Bourland, N.; Lejeune, P. Tree allometry in Central Africa: Testing the validity of pantropical multi-species allometric equations for estimating biomass and carbon stocks. For. Ecol. Manag. 2013, 305, 29–37. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Change Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.E.; Gong, W.K.; Wong, C.H. Allometry and partitioning of the mangrove, Rhizophora apiculata. For. Ecol. Manag. 2004, 188, 395–408. [Google Scholar] [CrossRef]
- Putz, F.E.; Chan, H.T. Tree Growth, dynamics, and productivity in a mature mangrove forest in Malaysia. For. Ecol. Manag. 1986, 17, 211–230. [Google Scholar] [CrossRef]
- Hazandy, A.H.; Ahmad-Ainuddin, N.; Zaiton, S.; Arifin, A.; Tuan-Marina, T.I.; Lydia-Suzieana, M. Quantifying aboveground carbon stock of 30-year-old mangrove forest and its economic value in Matang Mangrove Forest. In Proceedings of the National Conference on “Forest Resource Economics Assessment”, Putrajaya, Malaysia, 2–5 September 2014; pp. 188–213. [Google Scholar]
- Komiyama, A.; Poungparn, S.; Kato, S. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Jachowski, N.R.A.; Quak, M.S.Y.; Friess, D.A.; Duangnamon, D.; Webb, E.L.; Ziegler, A.D. Mangrove biomass estimation in Southwest Thailand using machine learning. Appl. Geogr. 2013, 45, 311–321. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Donato, D.C. Protocols for the Measurement, Monitoring and Reporting of Structure, Biomass and Carbon Stocks in Mangrove Forests; Working Paper 86; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 2012; 37p. [Google Scholar]
- Clough, B.F.; Scott, K. Allometric relationships for estimating above-ground biomass in six mangrove species. For. Ecol. Manag. 1989, 27, 117–127. [Google Scholar] [CrossRef]
- Doruska, P.F.; Patterson, D.W.; Posey, T.E. Variation in wood density by stand origin and log position for Loblolly Pine sawtimber in the Coastal Plain of Arkansas. In Proceedings of the 13th Biennial Southern Silvicultural Research Conference on General Technical Report, SRS-92; Kristina, C.F., Ed.; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2006; pp. 341–343. [Google Scholar]
- Preston, K.A.; Cornwell, W.K.; De-Noyer, J.L. Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms. New Phytol. Trust. 2006, 170, 807–818. [Google Scholar] [CrossRef]
- Iida, Y.; Lourens, P.; Sterk, F.J.; Kassim, A.R.; Kubo, T.; Potts, M.D.; Kohyama, T.S. Wood density explains architectural differentiation across 145 co-occuring tropical tree species. Funct. Ecol. 2012, 26, 274–282. [Google Scholar] [CrossRef]
- Medeiros, T.C.C.; Sampaio, E.V.S.B. Allometry of aboveground biomasses in mangrove species in Itamaracá, Pernambuco, Brazil. Wetl. Ecol. Manag. 2008, 16, 323–330. [Google Scholar] [CrossRef]
- Fearnside, P.M. Wood density for estimating forest biomass in Brazilian Amazonia. For. Ecol. Manag. 1997, 90, 59–87. [Google Scholar] [CrossRef]
- Banaticla, M.R.N.; Sales, R.F.; Lasco, R.D. Biomass equation for tropical tree plantation species in young stands using secondary data from the Philippines. Ann. Trop. Res. 2007, 29, 73–90. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Liu, Q.; Zhang, M.; Saeed, S. Effect of measurement errors on the estimation of tree biomass. Can. J. For. Res. 2019, 49, 1371–1378. [Google Scholar] [CrossRef]
- Bellasen, V.; Stephan, N. Accounting for Carbon; Cambridge University Press: Cambridgr, UK, 2015; 540p. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Folster, H.; Fromard, F.; Giguchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in Tropical Forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Komiyama, A.; Jintana, V.; Sangtiean, T.; Kato, S. A common allometric equation for predicting stem weight of mangroves growing in secondary forests. Ecol. Res. 2002, 17, 415–418. [Google Scholar] [CrossRef]
- Gong, W.K.; Ong, J.E. Plant biomass and nutrient flux in a managed mangrove forest in Malaysia. Estuar. Coast. Shelf Sci. 1990, 31, 519–530. [Google Scholar] [CrossRef]
- Kahn, M.N.I.; Suwa, R.; Hagihara, A. Allometric relationships for estimating the aboveground phytomass and leaf area of mangrove Kandelia candel (L.) Druce trees in the Manko Wetland, Okinawa Island, Japan. Trees 2005, 19, 266–272. [Google Scholar] [CrossRef]
- Soares, M.L.G.; Schaeffer-Novelli, Y. Above-ground biomass of mangrove species. I. Analysis of models. Estuar. Coast. Shelf Sci. 2005, 65, 1–18. [Google Scholar] [CrossRef]
- Jalil, A.-M.; Abdul-Hamid, H.; Mohamad-Ali, N.-A.; Mohamed, J. Allometric models for estimating aboveground biomass and carbon stock in planted Aquilaria malaccensis stand. J. Trop. For. Sci. 2021, 33, 240–246. [Google Scholar] [CrossRef]
- Clough, B.F. Primary Productivity and Growth of Mangrove Forests. In Tropical Mangrove Ecosystem; Robertson, A.I., Alongi, D.M., Eds.; American Geophysical Union: Washington, DC, USA, 1992; pp. 225–250. [Google Scholar]
- Fromard, F.; Puig, H.; Mougin, E.; Marty, G.; Betoulle, J.L.; Cadamuro, L. Structure, above-ground biomass and dynamics of mangrove ecosystems: New data from French Guiana. Oecologia 1998, 115, 39–53. [Google Scholar] [CrossRef]
- King, D.A. Linking tree form, allocation and growth with an allometrically explicit model. Ecol. Model. 2005, 185, 77–91. [Google Scholar] [CrossRef]
- Smith, T.J., III; Whelan, K.R.T. Development of allometric relations for three mangrove species in South Florida for use in the Greater Everglades Ecosystem restoration. Wetl. Ecol. Manag. 2006, 14, 409–419. [Google Scholar] [CrossRef]
- Henry, H.A.L.; Aarssen, L.W. The interpretation of stem diameter–height allometry in trees: Biomechanical constraints, neighbour effects, or biased regressions? Ecol. Lett. 1999, 2, 89–97. [Google Scholar] [CrossRef]
- Latiff, A. Conservation strategies for endangered mangrove swamp forests in Malaysia. Pak. J. Bot. 2012, 44, 27–36. [Google Scholar]
- Norhayati, A.; Latiff, A. Biomass and species composition of a mangrove forest in Pulau Langkawi, Malaysia. Malays. Appl. Biol. 2001, 30, 75–80. [Google Scholar]
- Chandra, I.A.; Seca, G.; Hena, M.K.A. Aboveground biomass production of Rhizophora apiculata Blume in Sarawak mangrove forest. Am. J. Agric. Biol. Sci. 2011, 6, 469–474. [Google Scholar]
- Norilani, W.W.I.; Juliana, W.W.A.; Salam, M.R.; Latiff, A. Community structure at two compartments of a disturbed mangrove forests at Pulau Langkawi. Am. Inst. Phys. AIP Conf. Proc. 2014, 1614, 790–794. [Google Scholar] [CrossRef] [Green Version]
- Rozainah, M.Z.; Nazri, M.N.; Sofawi, A.B.; Hemati, Z.; Juliana, W.A. Estimation of carbon pool in soil, above and below ground vegetation at different types of mangrove forests in Peninsular Malaysia. Mar. Pollut. Bull. 2018, 137, 237–245. [Google Scholar] [CrossRef]
- Haase, R.; Haase, P. Above-ground biomass estimates for invasive trees and shrubs in the Pantanal of Mato Grosso, Brazil. For. Ecol. Manag. 1995, 73, 29–35. [Google Scholar] [CrossRef]
- Rayachhetry, M.B.; Van, T.K.; Center, T.D.; Laroche, F. Dry weight estimation of the aboveground components of Melaleuca quinquenervia trees in southern Florida. For. Ecol. Manag. 2001, 142, 281–290. [Google Scholar] [CrossRef]
- Novitzky, P. Analysis of Mangrove Structure and Latitudinal Relationships on the Gulf Coast of Peninsular Florida. Master’s Thesis, University of South Florida, Tampa, FL, USA, 2010. [Google Scholar]
- Kodikara, K.A.S.; Jayatissa, L.P.; Huxham, M.; Dahdouh-Guebas, F.; Koedam, N. The effects of salinity on growth and survival of mangrove seedlings changes with age. Acta Bot. Bras. 2018, 32, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Phan-Khanh, L. Wave Attenuation in Coastal Mangroves: Mangrove Squeeze in the Mekong Delta. Doctoral’s Thesis, Delft University of Technology, Delf, Netherlands, 2019. [Google Scholar]
- Vinh, T.V.; Marchand, C.; Linh, T.V.K.; Vinh, D.D.; Allenbach, M. Allometric models to estimate above-ground biomass and carbon stocks in Rhizophora apiculata tropical managed mangrove forests (Southern Viet Nam). For. Ecol. Manag. 2019, 434, 131–141. [Google Scholar] [CrossRef]
- Amira, S. Estimation of Rhizophora apiculata B1 Biomass in Mangrove Forest in Batu Ampar Kubu Raya Regency, West Kalimantan. Honor’s Thesis, Bogor Agricultural University, Bogor, Indonesia, 2008. [Google Scholar]
- Cole, T.G.; Ewel, K.C.; Devoe, N.N. Structure of mangrove trees and forests in Micronesia. For. Ecol. Manag. 1999, 117, 95–109. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Cole, T.G. Micronesian mangrove forest structure and tree responses to a severe typhoon. Wetlands 2010, 30, 1077–1084. [Google Scholar] [CrossRef]
- Tarlan, M.A. Biomass Estimation of Nyirih (Xylocarpus granatum Koenig. 1784) in Primary Mangrove Forest in Batu Ampar, West Kalimantan. Undergraduate Thesis, Bogor Agricultural University, Bogor, Indonesia, 2008. [Google Scholar]







| Species | Local Name | Sample Trees | D (cm) | H (m) | Tree Density |
|---|---|---|---|---|---|
| Bruguiera cylindrica (L.) Blume | Bakau putih | 3 | 17.13 ± 3.85 | 17.17 ± 1.23 | 715.62 ± 10.2 |
| Bruguiera parviflora (R.) Wight | Lenggadai | 1 | 18.7 ± 0 | 18.0 ± 0 | 736.81 ± 0 |
| Bruguiera sexangula (L.) Poir | Tumu putih | 2 | 24.9 ± 1.10 | 20.50 ± 0.50 | 713.72 ± 7.87 |
| Ceriops tagal (P.) Rob | Tengar | 1 | 15.5 ± 0 | 14.5 ± 0 | 745.45 ± 0 |
| Rhizophora apiculata (L.) Blume | Bakau minyak | 5 | 20.08 ± 4.21 | 19.92 ± 1.69 | 842.22 ± 28.5 |
| Rhizophora mucronata (L.) Lam | Bakau kurap | 1 | 16.8 ± 0 | 18.1 ± 0 | 801.41 ± 0 |
| Xylocarpus granatum (J.) Koenig | Nyireh bunga | 2 | 14.70 ± 4.10 | 13.88 ± 2.63 | 708.65 ± 17.9 |
| Model | No. of Equation | Component | Equation | R2 | Standardized and Unstandardized Residual Normality | |
|---|---|---|---|---|---|---|
| M = aDb | 1 | (a) | M of stem | 0.1761D2.3769 | 0.9223 | 0.107 |
| (b) | M of branches and twigs | 0.0553D2.3055 | 0.7792 | 0.499 | ||
| (c) | M of leaves | 0.0347D1.9762 | 0.5834 | 0.206 | ||
| M = aD2Hb | 2 | (a) | M of stem | 0.0355D2H0.9778 | 0.9579 | 0.833 |
| (b) | M of branches and twigs | 0.0221D2H0.8745 | 0.6880 | 0.853 | ||
| (c) | M of leaves | 0.0125D2H0.7767 | 0.5530 | 0.246 | ||
| M = aDb | 1 | Total biomass | 0.2999D2.3001 | 0.9543 | 0.117 | |
| M = aD2Hb | 2 | Total biomass | 0.0739D2H0.9291 | 0.9556 | 0.813 | |
| M = aρb1Db2 | 3 | Total biomass | 0.00475ρ0.6309D2.28787 | 0.9697 | 0.724 | |
| No. of Equation | N | Mean | SD | SE | t-Value | p-Value | 95% Lower Bound | 95% Upper Bound | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | (a) | 15 | 5.0923 | 0.8731 | 0.1914 | 12.4186 | 1.3858 × 10−8 | 1.9634 | 2.7904 |
| (b) | 15 | 3.7284 | 0.8470 | 0.3404 | 6.7726 | 1.3161 × 10−5 | 1.5701 | 3.0410 | |
| (c) | 15 | 2.3163 | 0.7260 | 0.4632 | 4.2667 | 0.0009 | 0.9756 | 2.9768 | |
| 2 | (a) | 15 | 5.0923 | 0.8898 | 0.0569 | 17.1905 | 2.5414 × 10−10 | 0.8549 | 1.1007 |
| (b) | 15 | 3.7293 | 0.7942 | 0.1636 | 5.3369 | 0.0001 | 0.5216 | 1.2274 | |
| (c) | 15 | 2.3163 | 0.7068 | 0.1937 | 4.0105 | 0.0015 | 0.3583 | 1.1951 | |
| 1 | 15 | 5.4042 | 0.8449 | 0.1395 | 16.4836 | 4.2895 × 10−10 | 1.9986 | 2.6015 | |
| 2 | 15 | 5.4042 | 0.8455 | 0.0555 | 16.7277 | 3.572 × 10−10 | 0.8091 | 1.0491 | |
| 3 | 15 | 5.4042 | 0.8517 | 0.1184; 0.2555 | 19.3310; 2.4701 | 2.0743 × 10−10; 0.0295 | 1.9838; −1.2527 | 1.6592; 2.6144 | |
| Species | No. of Tree | Diameter (cm) | Height (m) | ||||
|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | ||
| B. cylindrica | 185 | 5 | 31.5 | 1 15.09 ± 10.38 | 4.2 | 30 | 15.99 ± 0.33 |
| B. parviflora | 40 | 6.9 | 34 | 18.05 ± 1.13 | 6.5 | 28 | 18.75 ± 0.77 |
| B. sexangula | 131 | 6.1 | 34 | 16.35 ± 0.39 | 4.1 | 28 | 17.25 ± 0.30 |
| C. tagal | 8 | 13 | 25.2 | 19.45 ± 1.40 | 15 | 26 | 19.32 ± 1.26 |
| R. apiculata | 553 | 9.9 | 40.5 | 20.03 ± 0.24 | 10 | 35 | 19.51 ± 0.16 |
| R. mucronata | 44 | 11 | 27.8 | 17.38 ± 0.65 | 11 | 24 | 18.16 ± 0.42 |
| X. granatum | 39 | 2 2.4 | 43.2 | 19.74 ± 1.21 | 5.2 | 24 | 14.83 ± 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul-Hamid, H.; Mohamad-Ismail, F.-N.; Mohamed, J.; Samdin, Z.; Abiri, R.; Tuan-Ibrahim, T.-M.; Mohammad, L.-S.; Jalil, A.-M.; Naji, H.-R. Allometric Equation for Aboveground Biomass Estimation of Mixed Mature Mangrove Forest. Forests 2022, 13, 325. https://doi.org/10.3390/f13020325
Abdul-Hamid H, Mohamad-Ismail F-N, Mohamed J, Samdin Z, Abiri R, Tuan-Ibrahim T-M, Mohammad L-S, Jalil A-M, Naji H-R. Allometric Equation for Aboveground Biomass Estimation of Mixed Mature Mangrove Forest. Forests. 2022; 13(2):325. https://doi.org/10.3390/f13020325
Chicago/Turabian StyleAbdul-Hamid, Hazandy, Fatin-Norliyana Mohamad-Ismail, Johar Mohamed, Zaiton Samdin, Rambod Abiri, Tuan-Marina Tuan-Ibrahim, Lydia-Suzieana Mohammad, Abdul-Majid Jalil, and Hamid-Reza Naji. 2022. "Allometric Equation for Aboveground Biomass Estimation of Mixed Mature Mangrove Forest" Forests 13, no. 2: 325. https://doi.org/10.3390/f13020325