
Analysis of Plant Water Transport Mechanism and Water Requirement for Growth Based on the Effect of Thermal Environment


Abstract
:1. Introduction
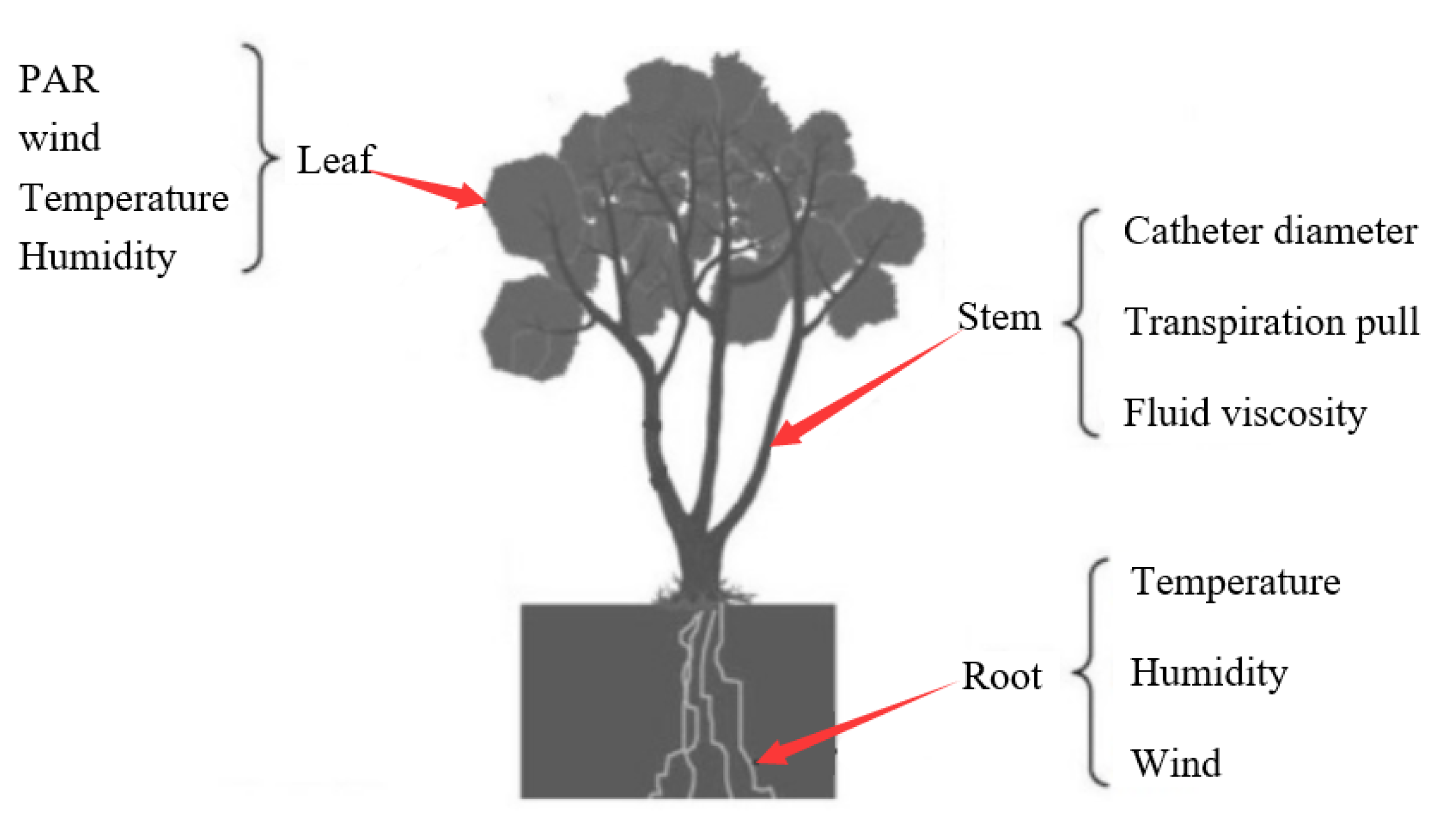
2. Methods
2.1. Transpiration Model
2.2. Stem Water Delivery Model
2.3. Root Water Uptake Model
2.4. Plant Water Requirement Model
3. Results and Discussion
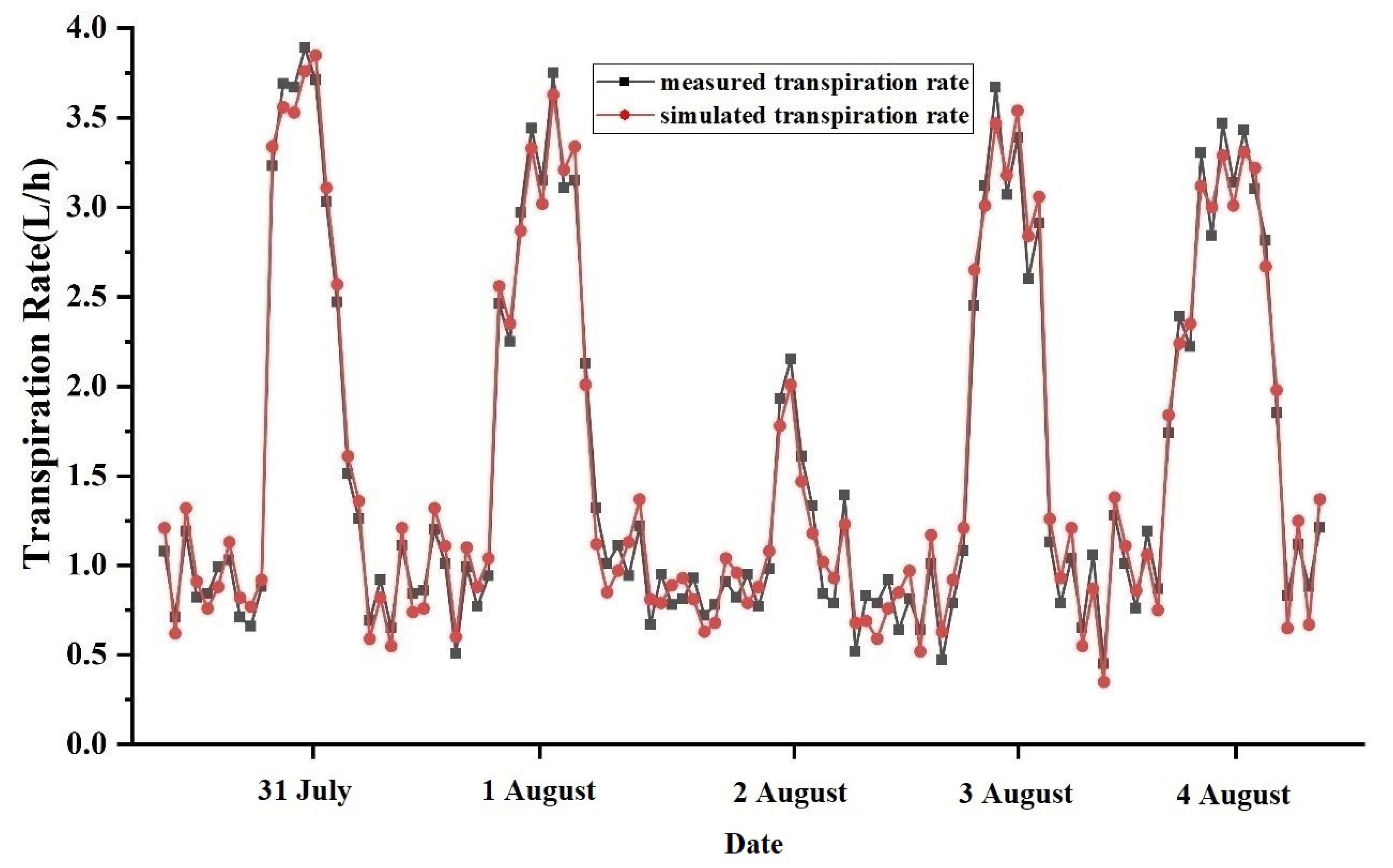
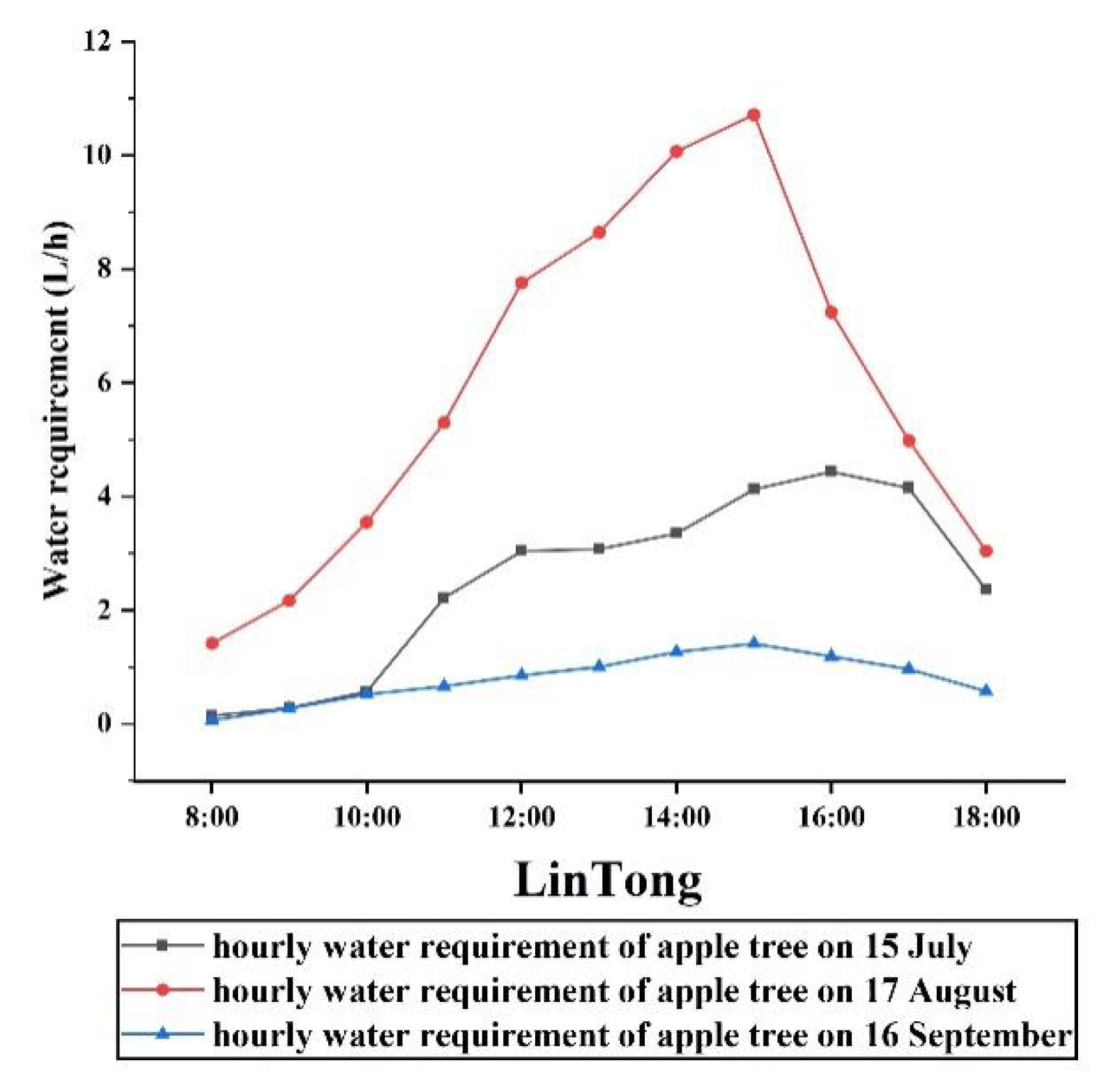
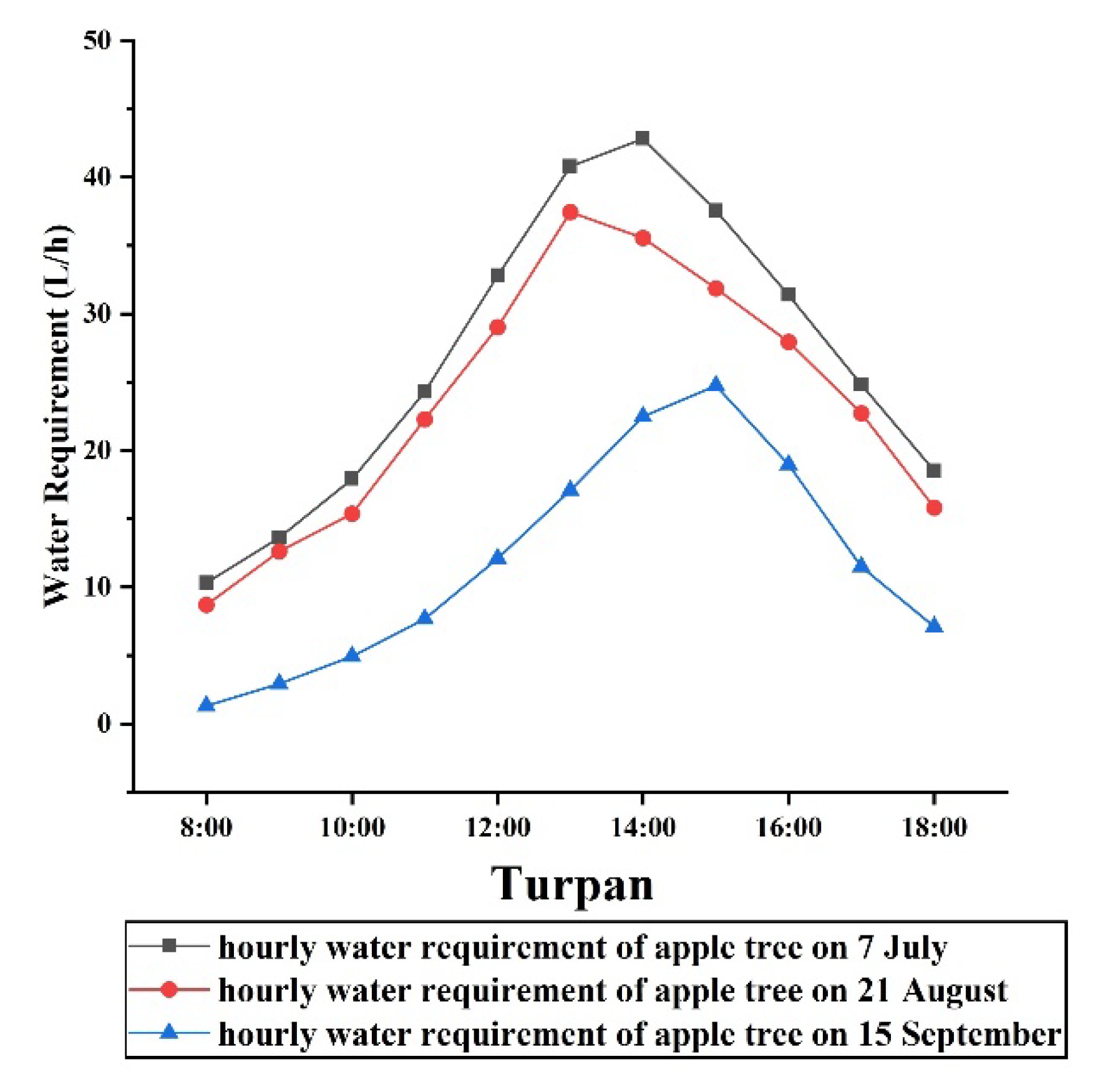
3.1. Model Validation
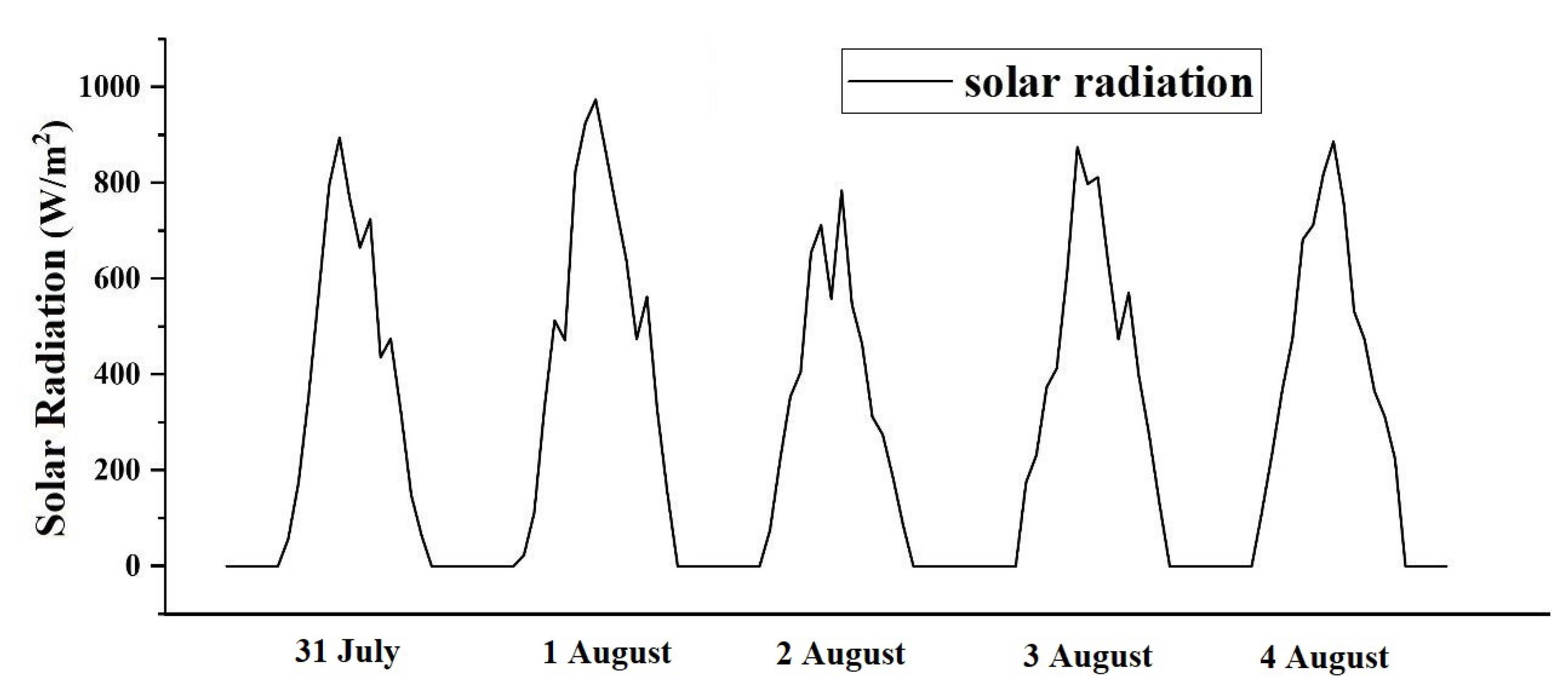
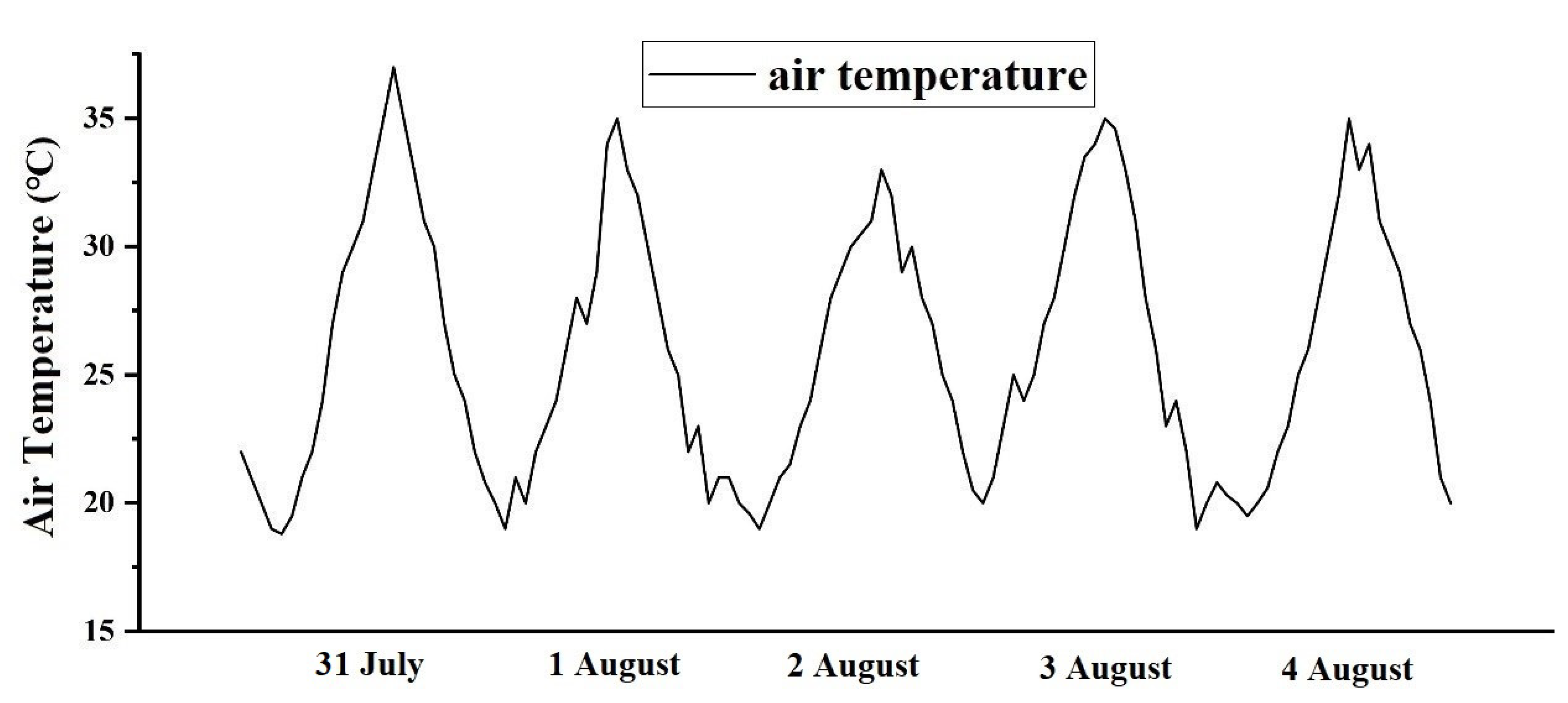
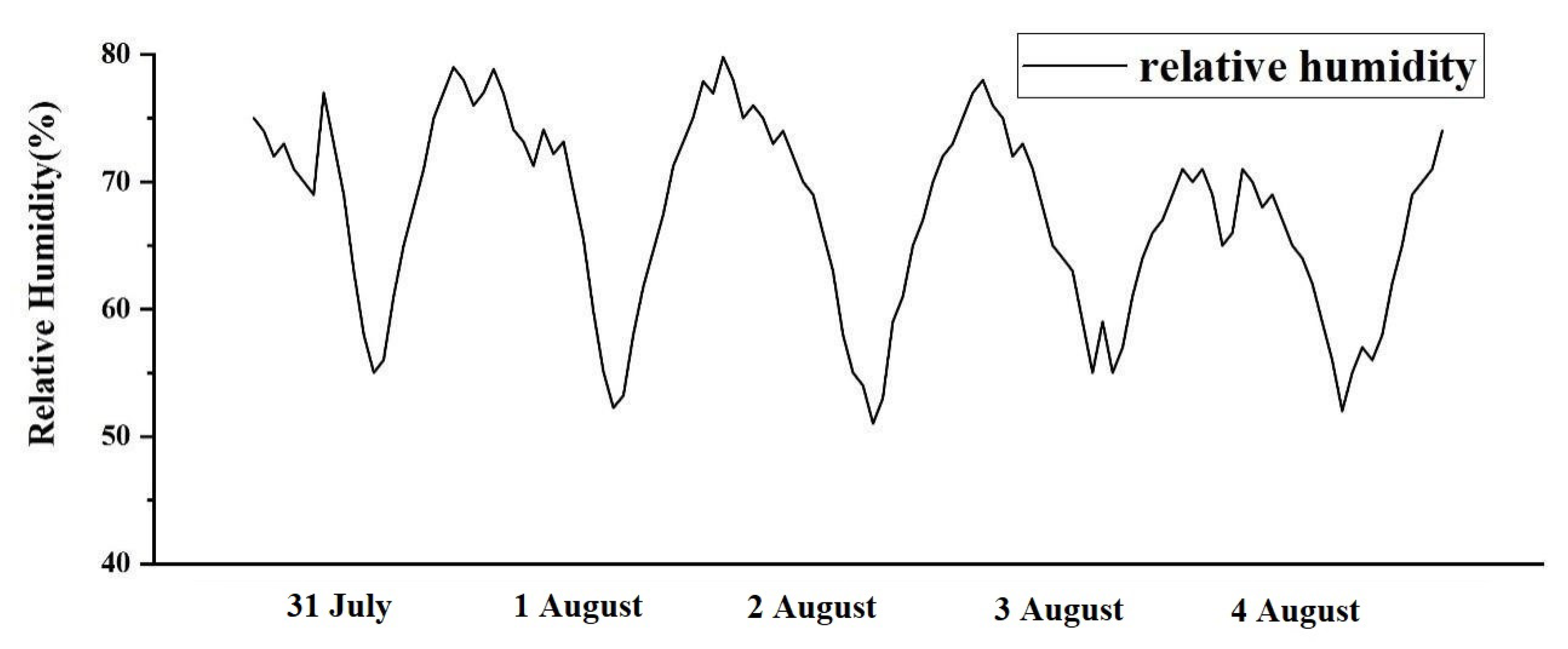
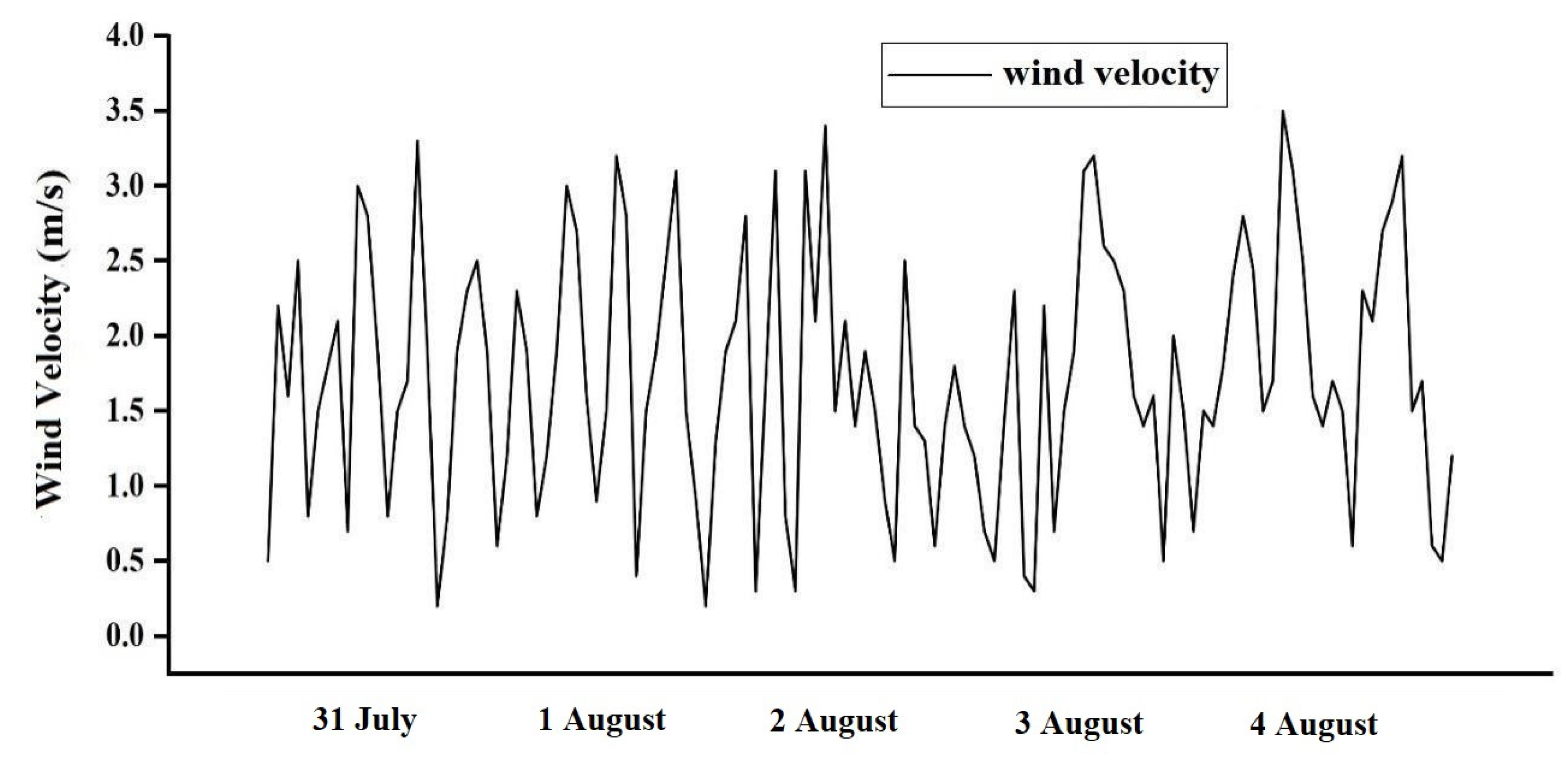
3.1.1. Materials and Methods
3.1.2. Validation
3.2. Discussion
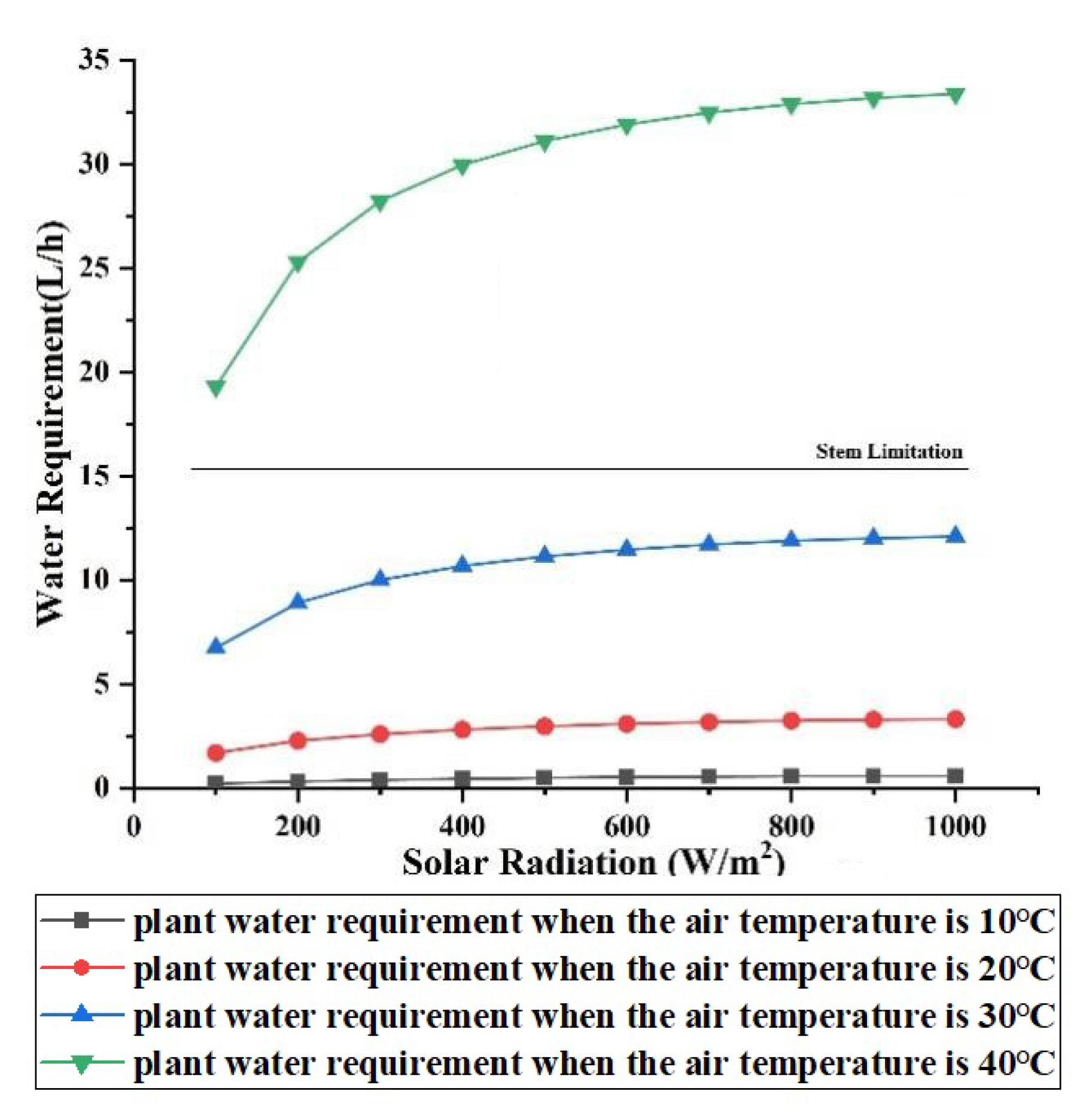
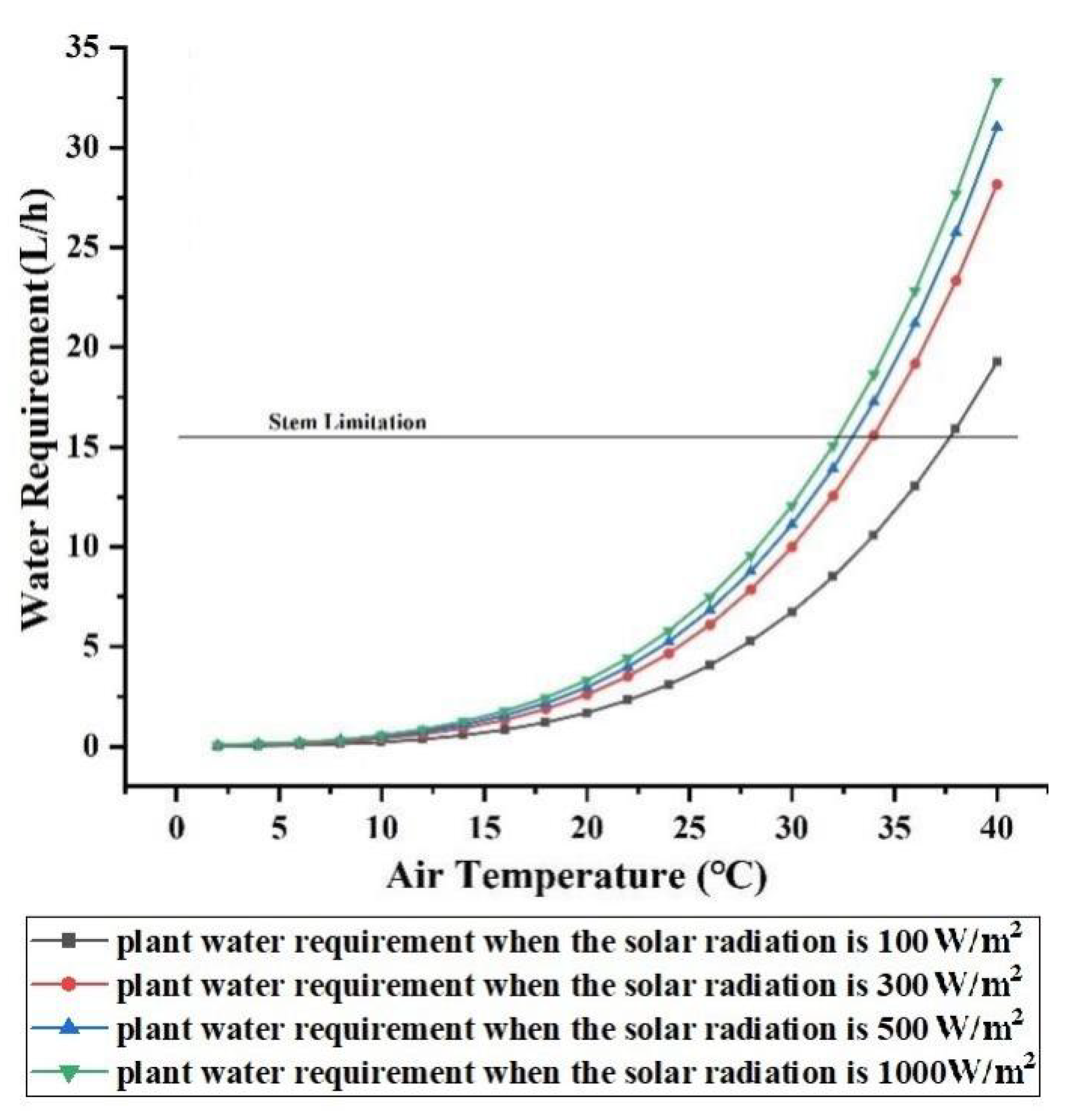
3.2.1. Effect of Solar Radiation
3.2.2. Effect of Air Temperature
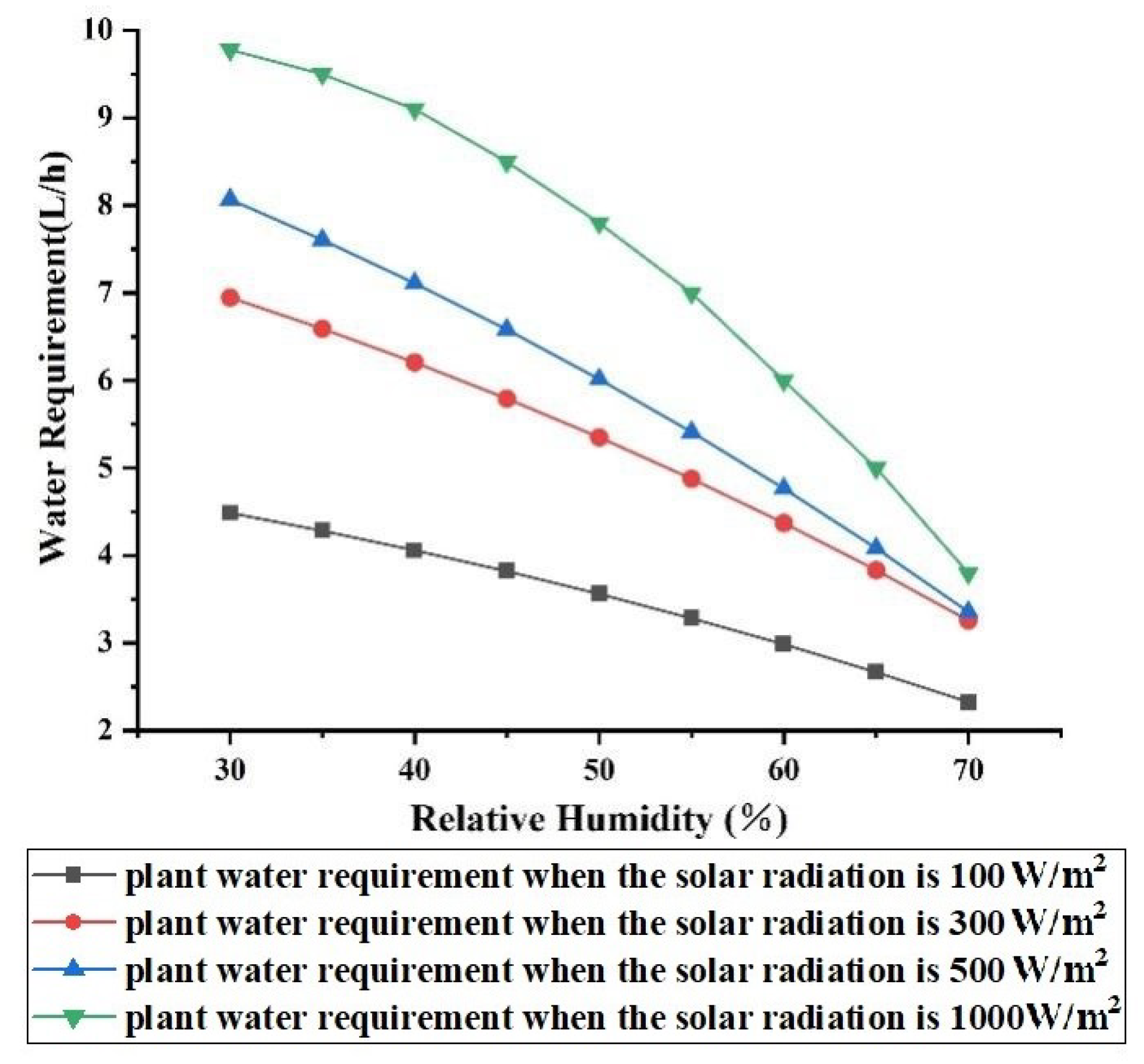
3.2.3. Effect of Relative Humidity
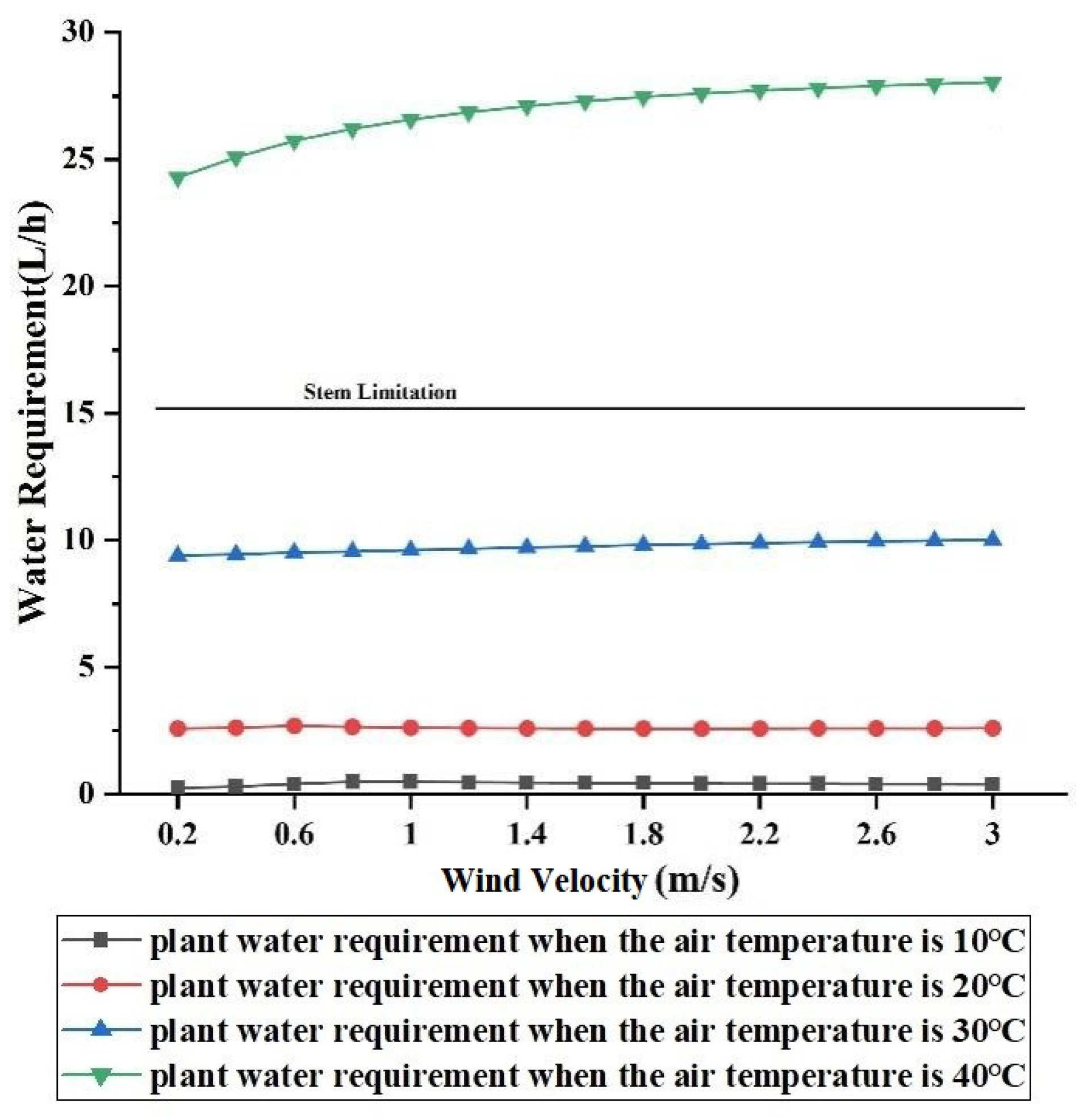
3.2.4. Effect of Wind Velocity
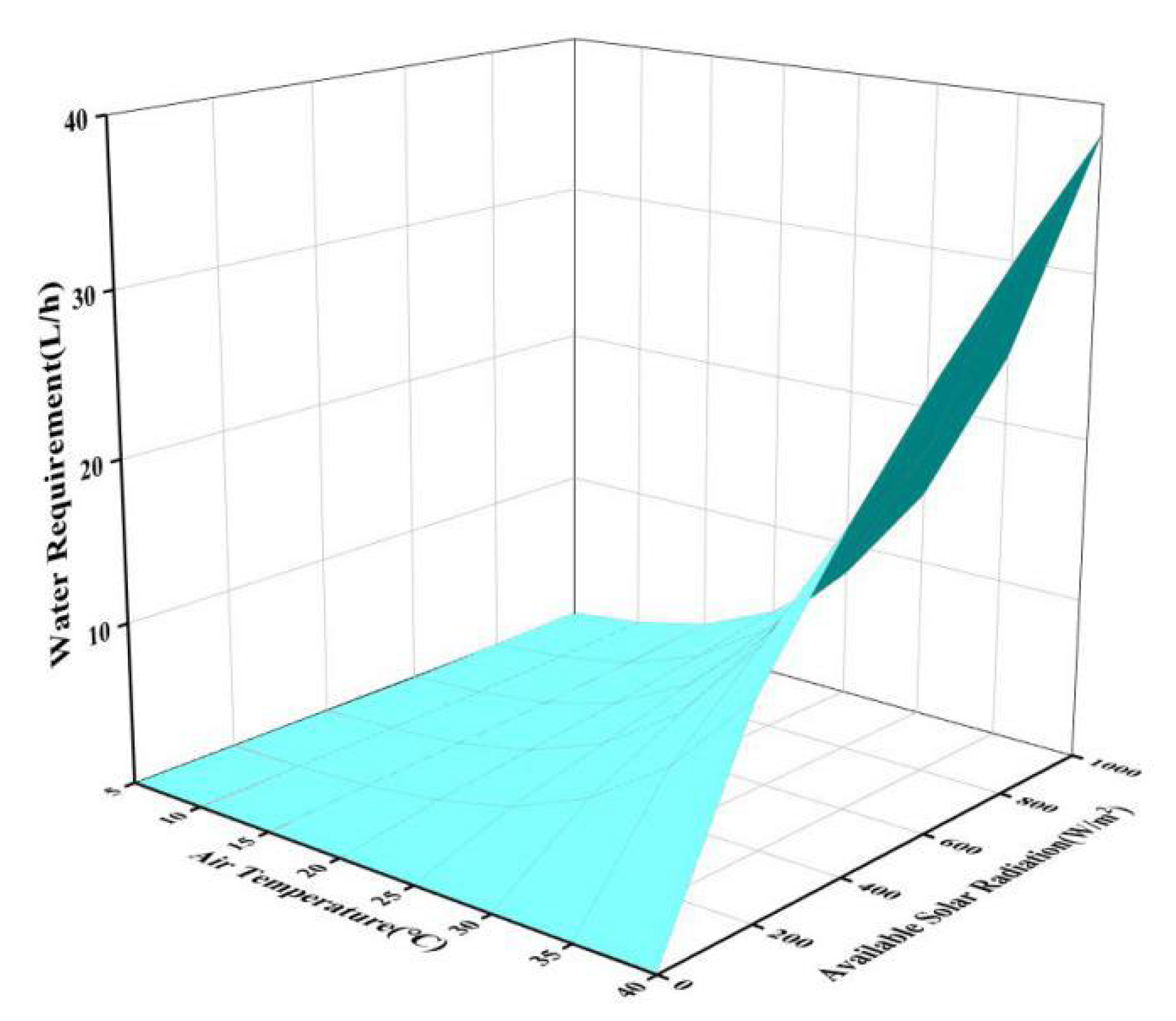
3.2.5. Effect of Multiple Parameters
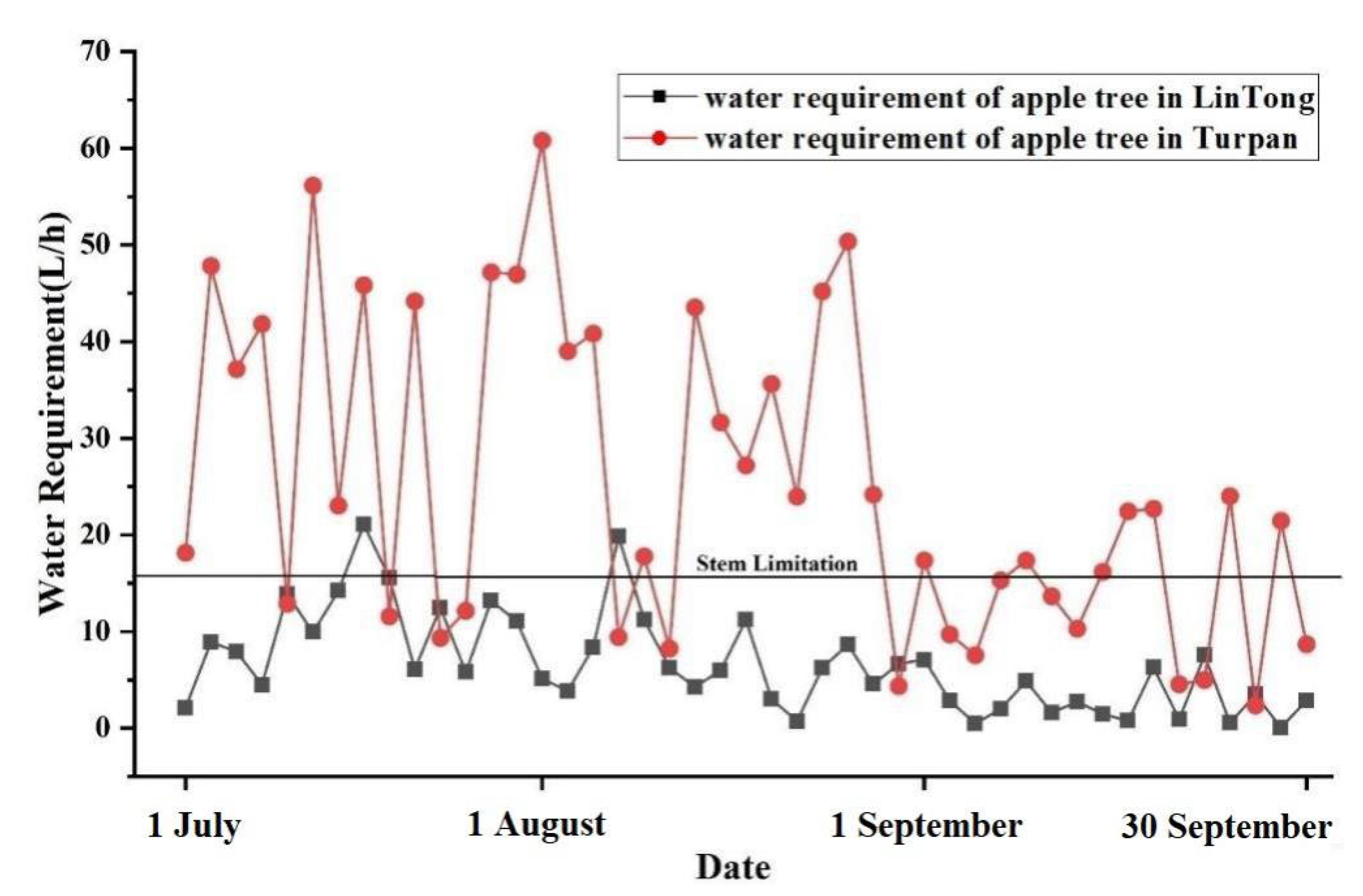
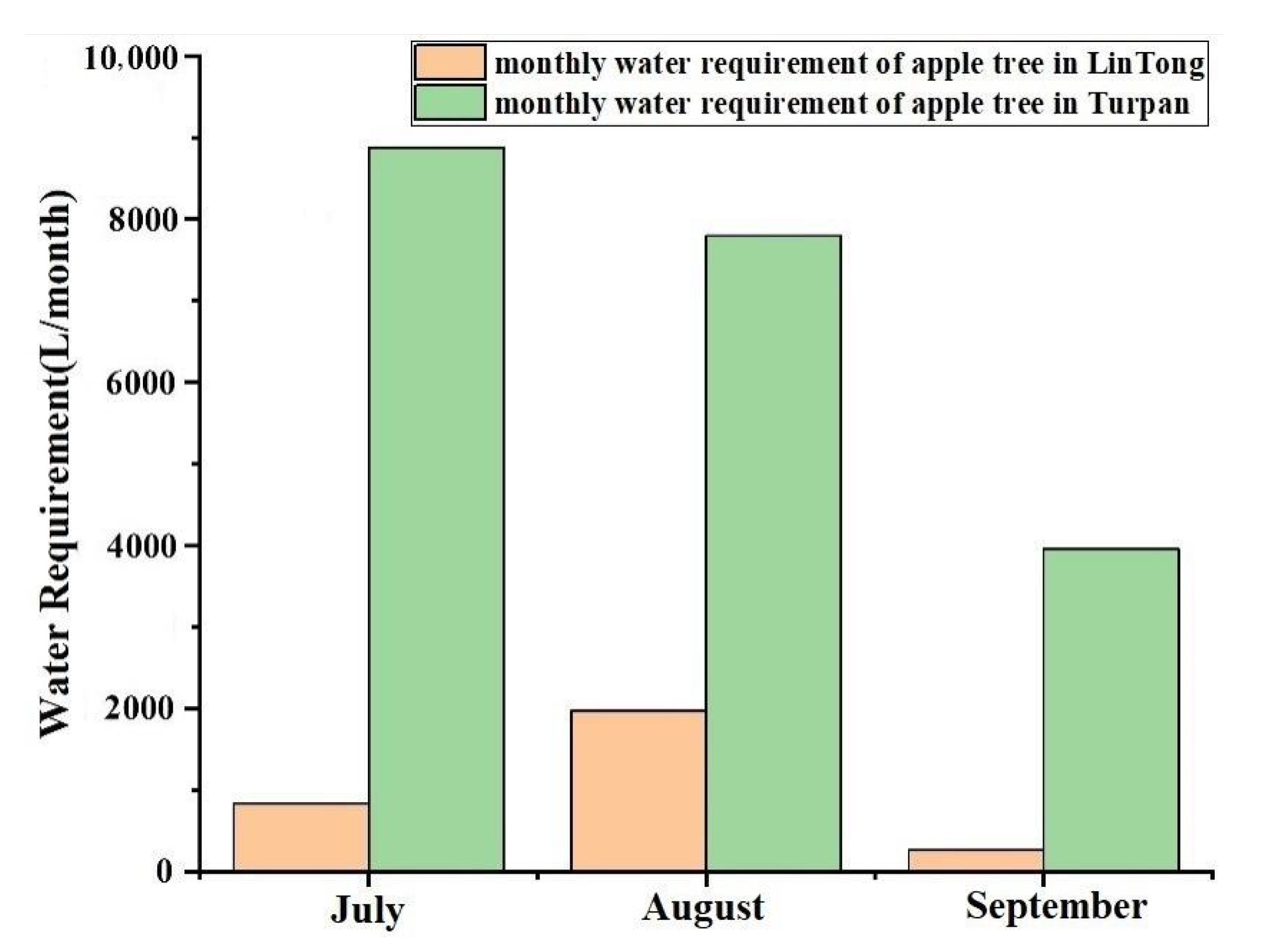
4. Application and Analysis
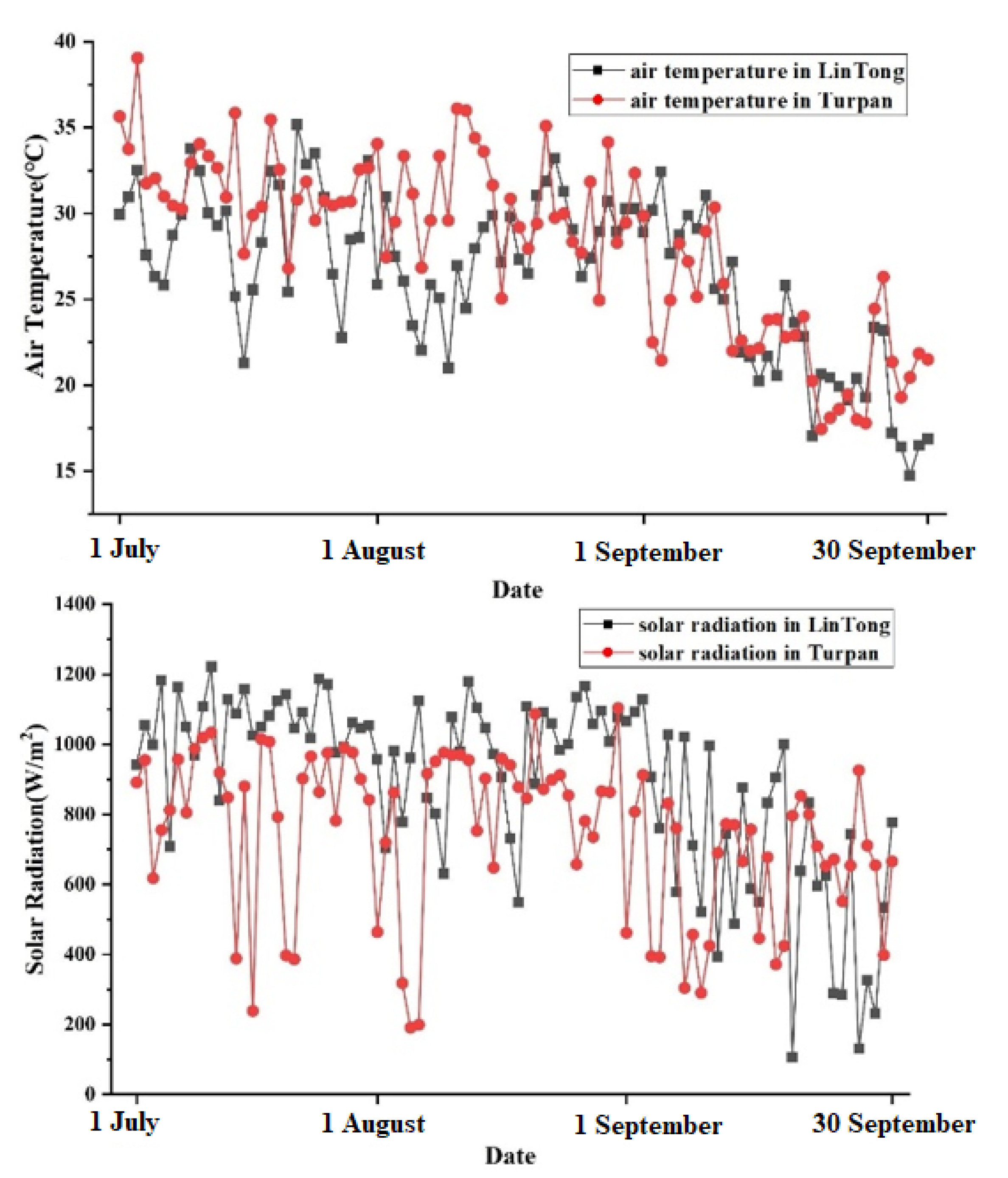
4.1. The Characteristics of the Study Plots
4.2. Analysis on Irrigation Strategy and Thermal Environment Regulation Ability of Plant
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, S.; Cao, Y.; Li, S.; Zhou, W.; Liu, S.; Bai, Z. Ecological Restoration of a Loess Open-Cast Mining Area in China: Perspective from an Ecological Security Pattern. Forests 2022, 13, 269. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Wilson, K.B.; Hanson, P.J. Environmental control of whole-plant transpiration, canopy conductance and estimates of the decoupling coefficient for large red maple trees. Agric. For. Meteorol. 2000, 104, 157–168. [Google Scholar] [CrossRef]
- Stock, S.C.; Koester, M.; Boy, J.; Godoy, R.; Dippold, M.A. Plant carbon investment in fine roots and arbuscular mycorrhizal fungi: A cross-biome study on nutrient acquisition strategies. Sci. Total Environ. 2021, 781, 146748. [Google Scholar] [CrossRef]
- Schlaepfer, D.R.; Ewers, B.E.; Shuman, B.N.; Williams, D.G.; Frank, J.M.; Massman, W.J.; Lauenroth, W.K. Terrestrial water fluxes dominated by transpiration: Comment. Ecosphere 2016, 5, 61. [Google Scholar] [CrossRef]
- Zhi, Y. The Simulation of the Transpiration of the Plant Leaf; University of Science and Technology of China: HeFei, China, 2013. [Google Scholar]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Sperry, J.S.; Venturas, M.D.; Anderegg, W.R.; Mencuccini, M.; Mackay, D.S.; Wang, Y.; Love, D.M. Predicting stomatal responses to the environment from the optimization of photosynthetic gain and hydraulic cost. Plant Cell Environ. 2016, 40, 816–830. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C.M. Drought Stress Effects and Olive Tree Acclimation under a Changing Climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Wang, H. Regulation mechanism of woody plant transport tissue on water transport. In Proceedings of the Symposium on Plant Water Demand and Regulation Technology, Xiamen, China, 1 May 2010; pp. 125–131. [Google Scholar]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of Plant Responses to Water Stress and Related Genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon T, M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Momii, K.; Hiyama, H.; Takeuchi, S. Field sugarcane transpiration based on sap flow measurements and root water uptake simulations: Case study on Tanegashima Island, Japan. Agric. Water Manag. 2021, 250, 106836. [Google Scholar] [CrossRef]
- Smethurst, P.J.; McVicar, T.R.; Huth, N.I.; Bradshaw, B.P.; Stewart, S.B.; Baker, T.G.; Benyon, R.G.; McGrath, J.F.; Van Niel, T.G. Nitrate Uptake from an Aquifer by Two Plantation Forests: Plausibility Strengthened by Process-Based Modelling. Forests 2022, 13, 184. [Google Scholar] [CrossRef]
- Chi, D.C.; Wang, X.; Xia, G.M. A Study on Water-uptake Model of Paddy Root. J. Irrig. Drain. 2004, 23, 56–58. [Google Scholar]
- Baena-Moreno, F.M.; Rodriguez-Galan, M.; Vega, F.; Reina, T.R.; Vilches, L.F.; Navarrete, B. Synergizing carbon capture storage and utilization in a biogas upgrading lab-scale plant based on calcium chloride: Influence of precipitation parameters. Sci. Total Environ. 2019, 670, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Haiming, F.; Yufang, D. Prediction and analysis of vegetation leaf temperature and canopy temperature. Mod. Hortic. 2021, 44, 5–8. [Google Scholar]
- Jarvis, P.G. The Interpretation of the Variations in Leaf Water Potential and Stomatal Conductance Found in Canopies in the Field. Philos. Trans. R. Soc. Lond. 1976, 273, 593–610. [Google Scholar]
- Kim, J.; Verma, S.B. Modeling canopy stomatal conductance in a temperate grassland ecosystem. Agric. For. Meteorol. 1991, 55, 149–166. [Google Scholar] [CrossRef]
- Gollan, T.; Schulze, T.D. The responses of stomata and leaf gas exchange to vapour pressure deficits and soil water content. Oecologia 1985, 65, 356–362. [Google Scholar] [CrossRef]
- Gang, L. Modelling Stomatal Conductance, Transpiration and Photosynthesis of Cut Lilium Grown in Greenhouses; Nanjing Agricultural University: Nanjing, China, 2012. [Google Scholar]
- Hofstra, G.; Hesketh, J.D. The effect of temperature on stomatal aperture in different species. Can. J. Bot. 1969, 47, 1307–1310. [Google Scholar] [CrossRef]
- Feddes, R.A.; Hoff, H.; Bruen, M.; Da Wson, T.E.; Pitman, A.J. Modeling Root Water Uptake in Hydrological and Climate Models. Bull. Am. Meteorol. Soc. 2001, 82, 2797–2810. [Google Scholar] [CrossRef] [Green Version]
- Ball, J.T.; Woodrow, I.E.; Berry, J.A. A Model Predicting Stomatal Conductance and its Contribution to the Control of Photosynthesis under Different Environmental Conditions. In Progress in Photosynthesis Research; Springer: Dordrecht, The Netherlands, 1987; pp. 221–224. [Google Scholar] [CrossRef]
- Hales, K.; Neelin, J.D.; Zeng, N. Sensitivity of Tropical Land Climate to Leaf Area Index: Role of Surface Conductance versus Albedo. J. Clim. 2004, 17, 1459–1473. [Google Scholar] [CrossRef]
- Lu, P. Estimation of whole-plant transpiration of bananas using sap flow measurements. J. Exp. Bot. 2002, 53, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Sperry, J.S.; Love, D.M. What plant hydraulics can tell us about responses to climate-change droughts. New Phytol. 2015, 207, 14. [Google Scholar] [CrossRef] [PubMed]
- Gong, D. Dynamic Mechanism of Water Transport in Soil-Plant-Atmosphere Continnum (SPAC) of Apple Orchard and Its Simulation; Northwest A & F University: Lintong, China, 2005. [Google Scholar]
- Venturas, M.D.; Sperry, J.S.; Love, D.M.; Frehner, E.H.; Allred, M.G.; Wang, Y.; Anderegg, W.R. A stomatal control model based on optimization of carbon gain versus hydraulic risk predicts aspen sapling responses to drought. New Phytol. 2018, 220, 836–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuzet, A.; Perrier, A.; Leuning, R. A coupled model of stomatal conductance, photosynthesis and transpiration. Plant Cell Environ. 2003, 26, 1097–1116. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a1 | a2 | a3 | a4 | b1 | b2 | b3 | c1 | c2 |
|---|---|---|---|---|---|---|---|---|
| 88.55 | 8.98 | 0.0025 | 2.36475 | 0.025357 | −2.89317 | 133.9857 | 0.00658 | 0.003744 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, H.; Sun, H.; Long, J. Analysis of Plant Water Transport Mechanism and Water Requirement for Growth Based on the Effect of Thermal Environment. Forests 2022, 13, 583. https://doi.org/10.3390/f13040583
Lu H, Sun H, Long J. Analysis of Plant Water Transport Mechanism and Water Requirement for Growth Based on the Effect of Thermal Environment. Forests. 2022; 13(4):583. https://doi.org/10.3390/f13040583
Chicago/Turabian StyleLu, Haolin, Hongfa Sun, and Jibo Long. 2022. "Analysis of Plant Water Transport Mechanism and Water Requirement for Growth Based on the Effect of Thermal Environment" Forests 13, no. 4: 583. https://doi.org/10.3390/f13040583