1. Introduction
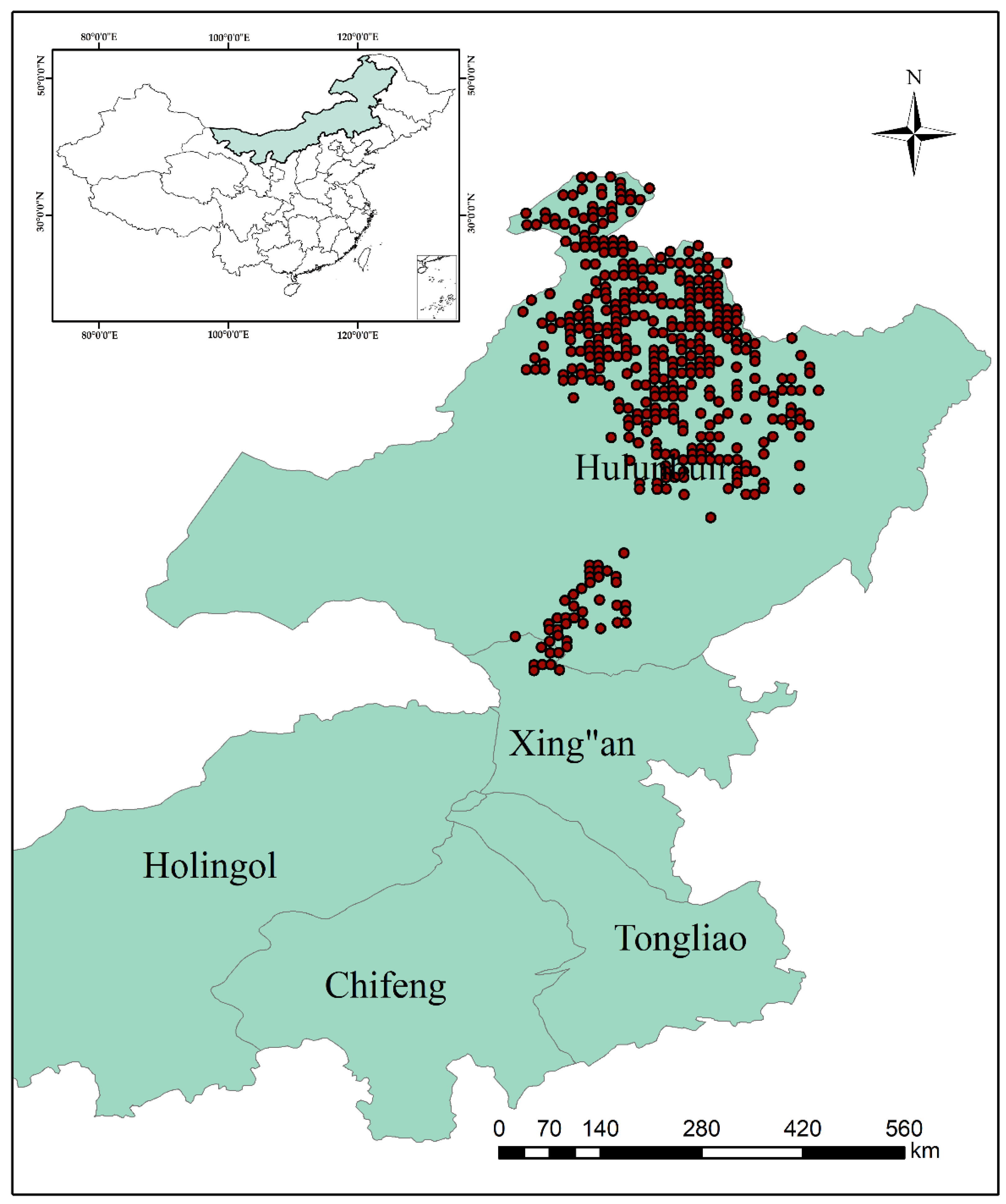
Larix gmelinii natural forests are a typical vegetation community in the Xing’an Mountains area in northeast China [
1,
2,
3]. They cover an area of 40,421 ha with a stocking of 4,654,312 m
3, representing 49% of the total forest area and 49% of stocking in the Xing’an Mountains [
4]. It is noteworthy that they are distributed in the southern edge of the global northern coniferous forest range and hence play a vital role in ecological security for northeast China [
5,
6].
Larix gmelinii is also of economic importance and is widely used for furniture, construction, and shipbuilding because of its toughness [
7,
8].
Like other natural forests,
Larix gmelinii natural forests have been suffering from deforestation and degradation due to the old forest policy (1950–1997) in China, which was designed to maximize timber production for economic development. Zhang [
9] documented that from 1950 to 1995 natural forests declined to 30% of the total forest area in China and the unit-area stocking of natural forests decreased by 32%. In 1998, China established Natural Forest Conservation Program (NFCP), articulating the new forest policy, which focused on forest ecological services other than wood production [
9,
10]. In 2017, China further banned all commercial logging in natural forests [
11] and encouraged scientific forest management strategies for forest restoration [
12].
Formulating forest management strategies requires accurate prediction and comparison of forest growth and yield under different management scenarios [
13,
14]. Forest growth models plays an important role in prescribing forest management strategies. For instance, Sterba [
15] employed the distance-independent individual tree growth model PROGNAUS to determine the equilibrium curve for mixed-species forests in Austria. Lars, et al. [
16] used a single-tree growth model to investigate the possibility of transforming normal, young Norway spruce forests to develop more heterogeneous stand structures, aiming for multi-layered forests in the long run. Normally, forest growth models are categorized into whole-stand models, size-class models, and individual-tree models according to the relevant modeling units [
17]. The whole stand model cannot capture variability regarding individual tree size and species, resulting in incapability of predicting forest dynamics with complex forest structures [
18]. Tree size-class models and individual-tree models, which employ a finer modeling resolution, i.e., diameter class and individual tree, respectively, are used to assess more complex species compositions and structures [
18,
19].
Individual-tree models use an individual tree as the modeling unit and are thus capable of a better mimicking of reality and integrating spatial heterogeneity [
17,
20]. However, they are challenging to develop as they require large amounts of individual tree-level data, which is especially the case for distance-dependent individual-tree models [
14,
17]. By comparison, tree-size models are easier to develop and have shown robust performance in modeling forest dynamics of unevenly aged, mixed-species forests [
18,
21].
As the largest terrestrial ecosystem type, forests play a significant role in combating climate change by absorbing CO
2. As much as 30% (2 petagrams of carbon per year; Pg C year−C) of annual global anthropogenic CO
2 emissions are sequestrated by the world’s forests [
22,
23]. Climate change, in turn, can also influence forest productivity [
24,
25], tree mortality [
26,
27,
28], and tree recruitment [
29,
30], and hence plays a vital role in shaping forest structure and species composition [
31,
32]. The aims of our present study were to produce a climate-sensitive transition-matrix forest growth model (CM) by integrating climate variables for
Larix gmelinii natural forests. With the new CM, we simulated forest dynamics under varying climate change scenarios. Based on the results of the simulations, we propose some silvicultural practices that could be implemented to mitigate the negative effects of climate change.
4. Discussion
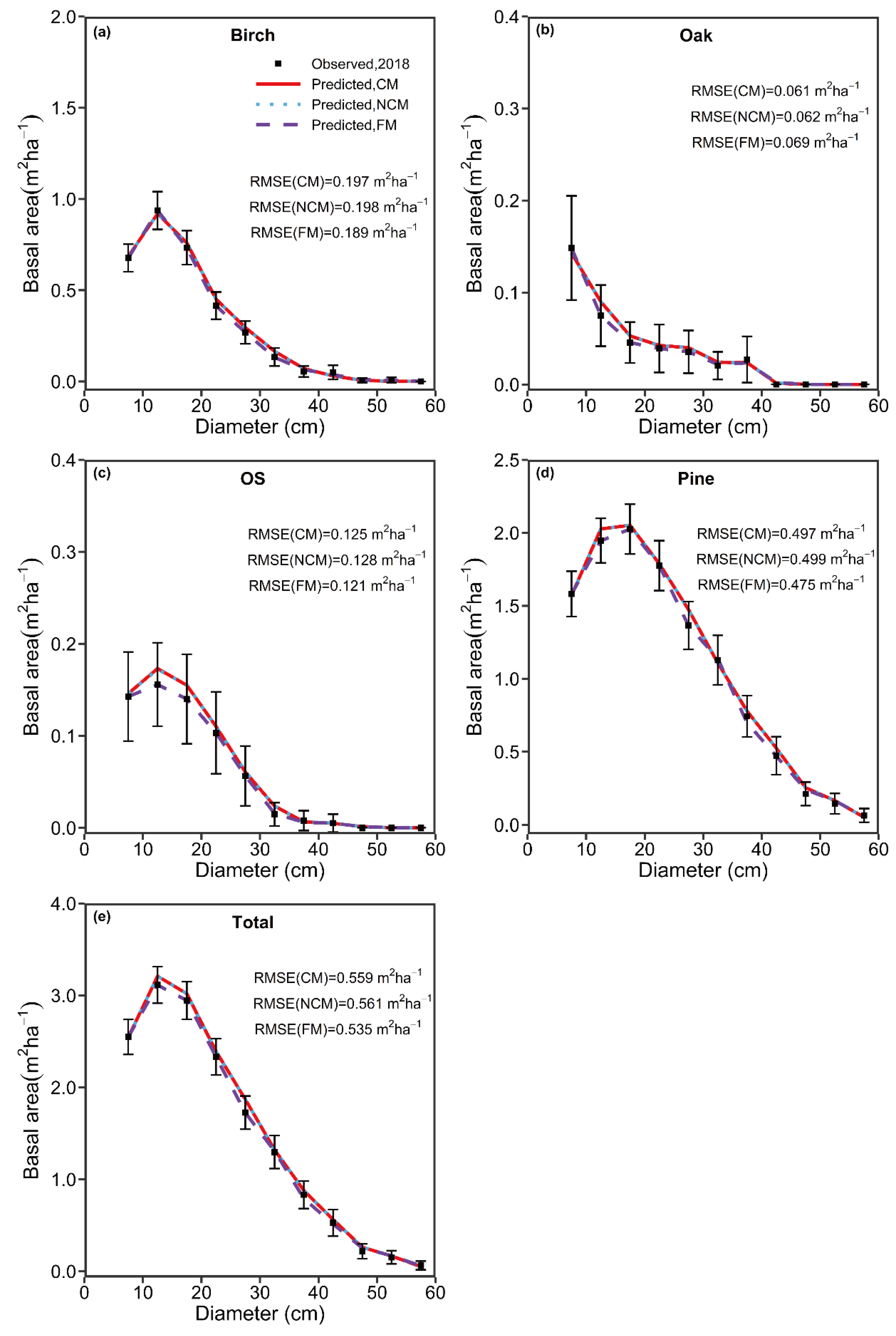
In the study, we produced a climate-sensitive, transition-matrix model for unevenly aged, Larix gmelinii mixed-species forests. Our transition-matrix model contained three sub-models, i.e., a tree diameter increment model, mortality model, and recruitment model, all of which were statistically robust, indicating the capability of projecting forest dynamics for these complex mixed-species forests.
We observed that tree species diversity (H
1) had a positive relationship with recruitment (birch, softwood, and pine) and tree growth (birch and pine), indicating that high species diversity could improve tree growth and recruitment. Similar results have been reported in other studies [
47,
48,
49,
50]. Sapijanskas, Paquette, Potvin, Kunert and Loreau [
50] attributed the positive effects of species diversity to the enhancement of light capture through crown plasticity and spatial and temporal niche differences in mixed-species forests. Tree species diversity (H
1) had a negative correlation with tree mortality, especially for the birch group. Similar results have been documented by Hisano et al. [
51], who found species-rich boreal forests suffered less mortality than species-poor forests under the environmental change of the past half-century and argued that improving tree diversity could help reduce the climate and environmental change vulnerability of boreal forests.
Tree size diversity (H
2) also had a positive relationship on tree diameter increment (pine and birch group) and tree recruitment (birch and softwood group), indicating that enhancing tree size diversity could also promote tree growth and recruitment. A similar result has been documented in other studies [
52,
53]. For example, Dănescu, Albrecht and Bauhus [
53] suggested that structural and species diversity acted as direct and independent drivers of stand productivity, with structural diversity (tree size diversity) being a slightly better predictor. The positive effects might be explained by niche complementarity theory. The higher tree size diversity indicates that trees differing in size could more efficiently occupy the growth space, which results in a complex spatial and vertical structure that could facilitate the utilization of natural resources, such as soil nutrients, water, and sunlight [
54,
55]. The effective allocation of natural resources can thus lead to high productivity, carbon sequestration, and tree recruitment [
52,
54]. Forest managers should implement silvicultural practices to increase tree size diversity and thus enhance the carbon sequestration capacity. The efficacy of this silvicultural practice has been demonstrated by several researchers. For example, Ruan, et al. [
56] showed that the carbon sequestration ability was significantly increased after a pure monoculture was transformed into an uneven-aged mixed-species forest with high size diversity.
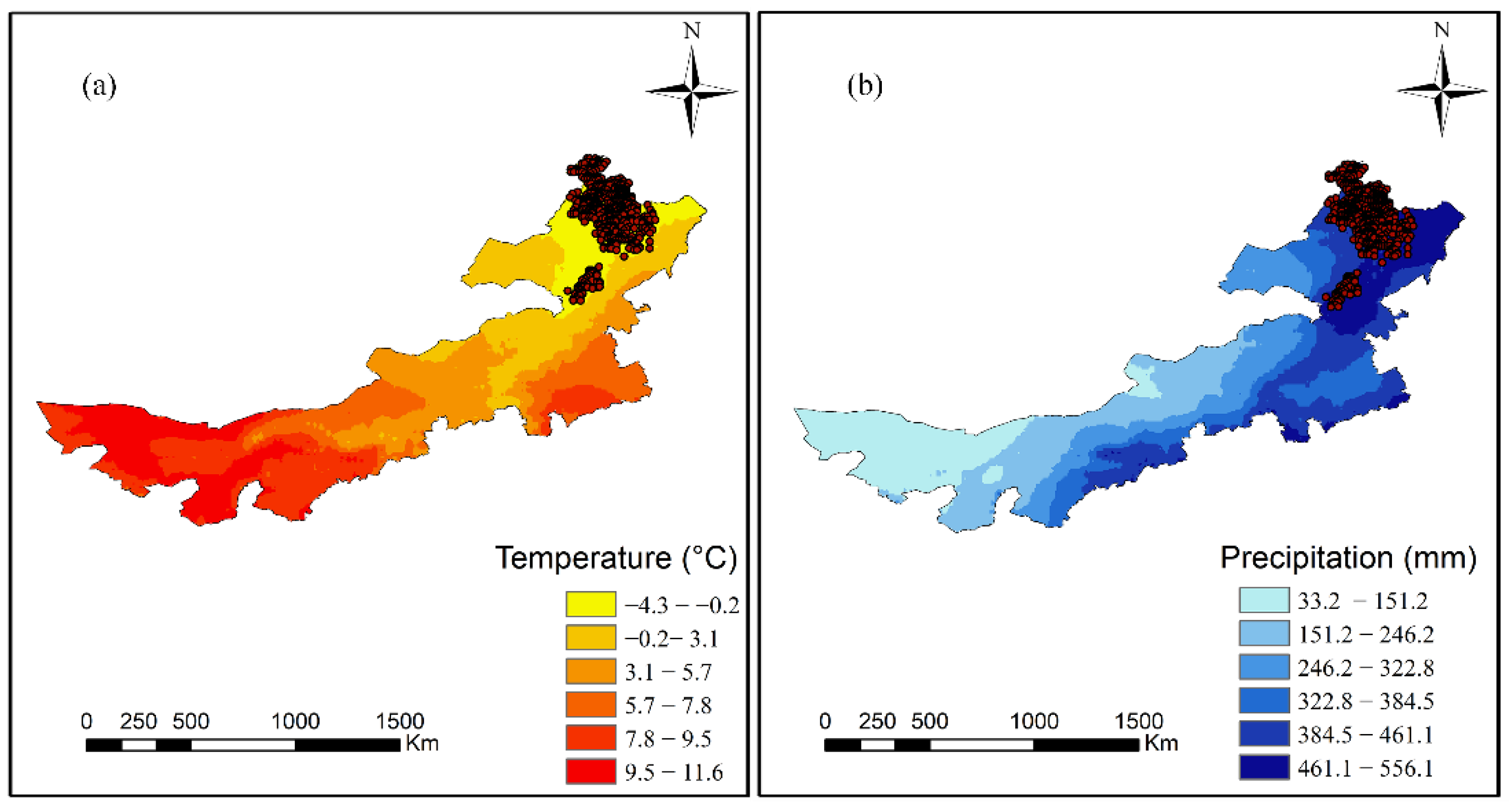
MAT showed a significant positive effect on tree diameter increment for all tree species groups except softwood. Similar results have been found in other studies [
57,
58,
59,
60]. For example, Raich, Russell, Kitayama, Parton and Vitousek [
57] found that tree growth and below-ground carbon allocation increased with MAT in evergreen broad-leaved tropical forests. The significant positive effect can be explained by the fact that our study was conducted in a cold region, and temperature is a factor limiting forest growth. Similar findings have been observed in other cold regions. For example, in the Alaska boreal region, Liang et al. [
42] and Mann et al. [
61] found that MAP and mean annual growing season temperature (GST) can positively affect tree diameter growth. Furthermore, we also observed that MAT had positive effects on tree mortality. These significant positive effects have been extensively explained by temperature-induced drought stress [
14,
62,
63]. Although Williams, Allen, Macalady, Griffin, Woodhouse, Meko, Swetnam, Rauscher, Seager and Grissino-Mayer [
63] argued that temperature was a potent driver of regional forest drought stress and tree mortality on the assumption of future decreases in water availability (no change or a decrease in precipitation) and increases in temperature. Because water is not a limiting factor in our study area, the positive effects of MAT on tree mortality might be attributed to an indirect relationship, wherein increases in MAT result in higher stand density; competition for light, water, and nutrients; and thus, increased mortality.
MAP had a positive relationship with tree diameter increment for the softwood and pine groups; such a positive correlation has been documented before. By contrast, a negative correlation between diameter increment and MAP was detected for birch, suggesting that precipitation was not a limiting factor for this species group. We observed that mortality had a positive correlation with MAP for birch and pine; the same pattern was reported by Du, Chen, Zeng and Meng [
14]. The positive effect of MAP on tree mortality could also be explained by an indirect relationship, wherein stand density increases due to greater diameter increments driven by increasing MAP results in more intense competition and thus higher mortality.
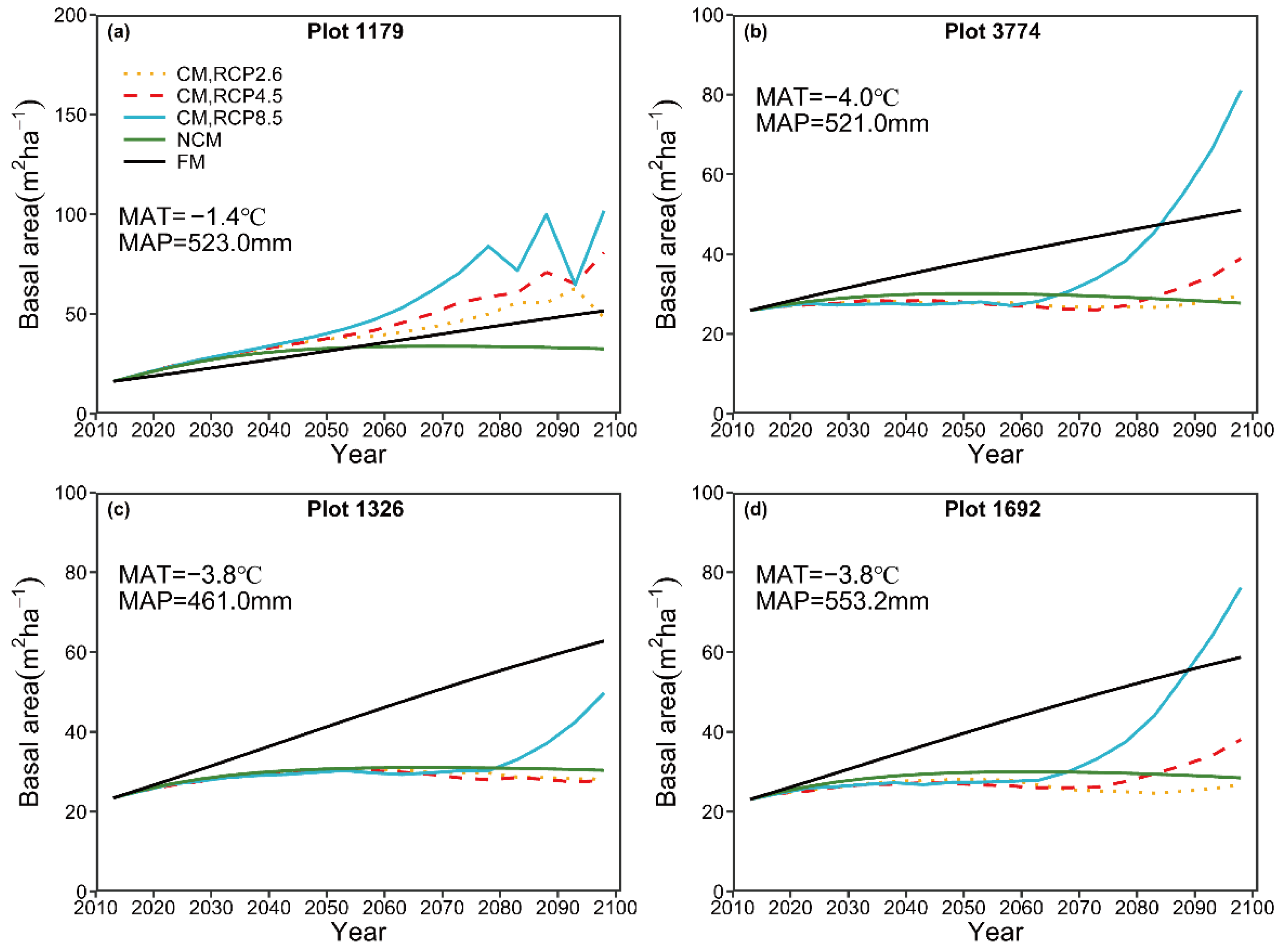
For prediction of 5-year intervals, the three models, i.e., FM, NCM, and CM, exhibited no pronounced differences in predictive performance for the focal species groups. We, therefore, recommended the FM, which has a simple structure and is easier to develop, to conduct short-term predictions. However, we found that the predicted long-term tree density and basal area by the FM showed a simple linear pattern, which may not be the true case (
Figure 6 and
Figure 7). Although NCM could generate more robust long-term predictions compared with FM, it might not be effective for long-term prediction because it neglected the long-term effects of climate variables on forest growth [
58,
64,
65], recruitment [
66,
67,
68], and mortality [
64,
69,
70].
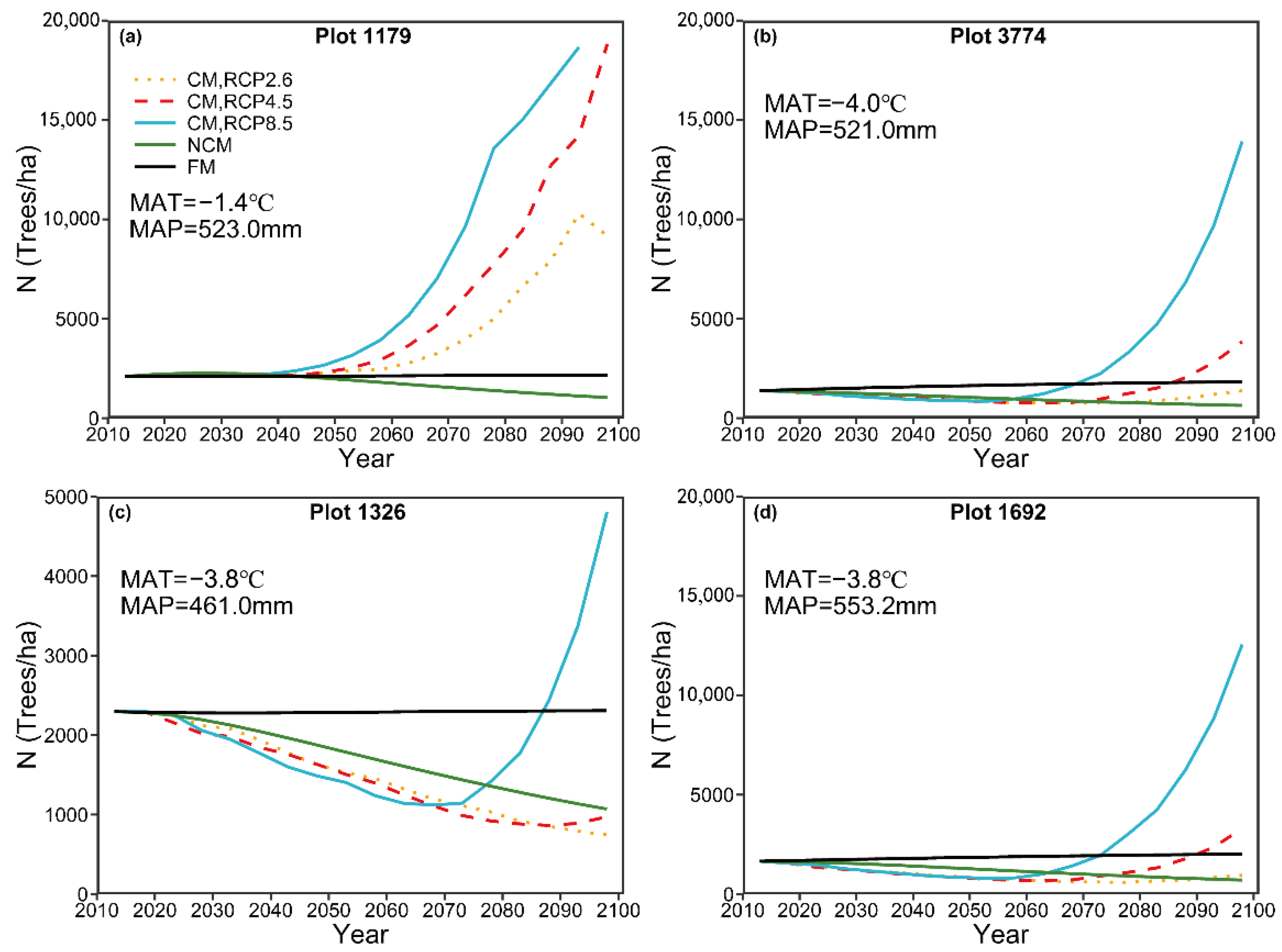
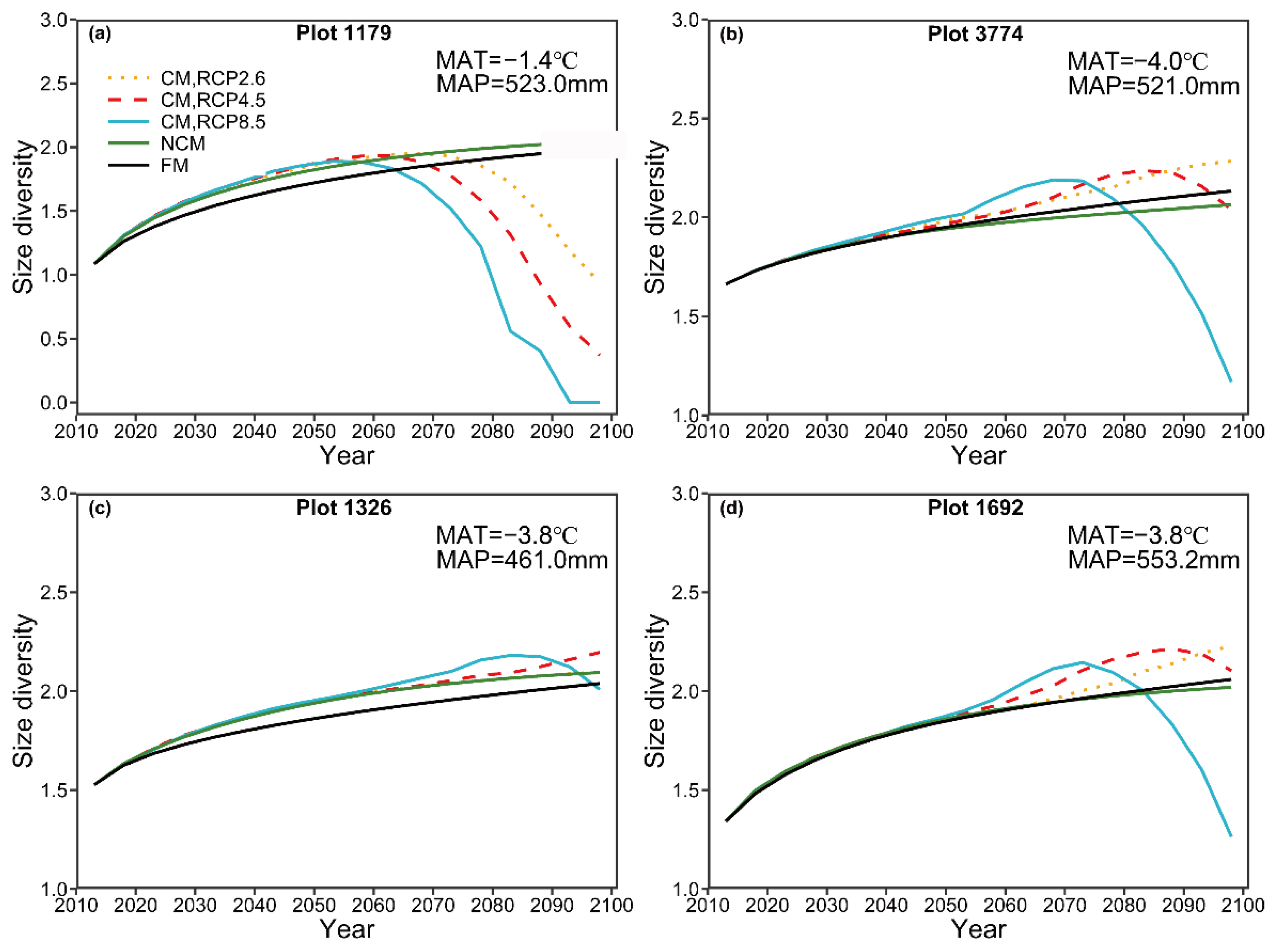
For long-term prediction, the predicted number of trees by the NCM suggests a decreasing trend, which could be explained by competition-induced self-thinning. The predicted basal area by the NCM showed a slightly increasing pattern and then reached a steady condition. The predicted patterns for the number of trees and basal area are in general consistent with the pattern in natural forest succession reported by Liang [
44]. By contrast, the predicted number of trees by the CM under three different climate change scenarios, in general, indicated a pronounced increasing pattern, though plot 1326 showed a decreasing pattern before 2070 (
Figure 6). A similar increasing pattern was also detected for the basal area under the three different RCPs (
Figure 7). Similar results have been documented by many authors [
14,
71]. For example, Ruiz-Benito, Madrigal-Gonzalez, Ratcliffe, Coomes, Kändler, Lehtonen, Wirth and Zavala [
71] reported that climatic warming caused an increase in stand average basal area, though this increase was offset by water availability. The predicted change in the steady state of natural stands suggests that increases in emissions can result in significant increases in stand density and basal area, which could preclude a steady state. Increases in stand density might reduce the economic value of forests and result in natural disasters, such as snow break, windthrow, and forest canopy fires. For example, high stand density can result in a large slenderness coefficient, which might increase the probability of windthrow or snow break [
72,
73]. Therefore, silvicultural practices should be implemented to mitigate the deleterious effects of climate change. For example, intermediate thinning intervals could be reduced or thinning intensity might be increased to manipulate stand density so that forests would be resistant to windthrow, snow break, or canopy fire.
Fast-growing short-rotation trees accumulate more carbon in the leaves, stems, and roots and thus have higher net annual carbon sequestration rates than slow-growing trees. However, forests with slow-growing long-rotation trees have larger carbon stocks over the long term. Kaul, et al. [
74] estimated the carbon sequestration potential of slow-growing sal (
Shorea robusta Gaertn. f.), fast-growing Eucalyptus (
Eucalyptus tereticornis Sm.), fast-growing poplar (
Populus deltoides Marsh), and moderate-growing teak (
Tectona grandis Linn. f.) forests in India and found that the living biomass of slow-growing long-rotation sal forests had the largest carbon stock; the opposite pattern was observed for net annual carbon sequestration rates. Sugden [
75] showed that slow-growing trees sequester more carbon and argued that higher levels of carbon accumulation are achieved in communities of slow-growing species, indicating that slow-growing trees should be used for enrichment planting. The effects of increasing atmospheric CO
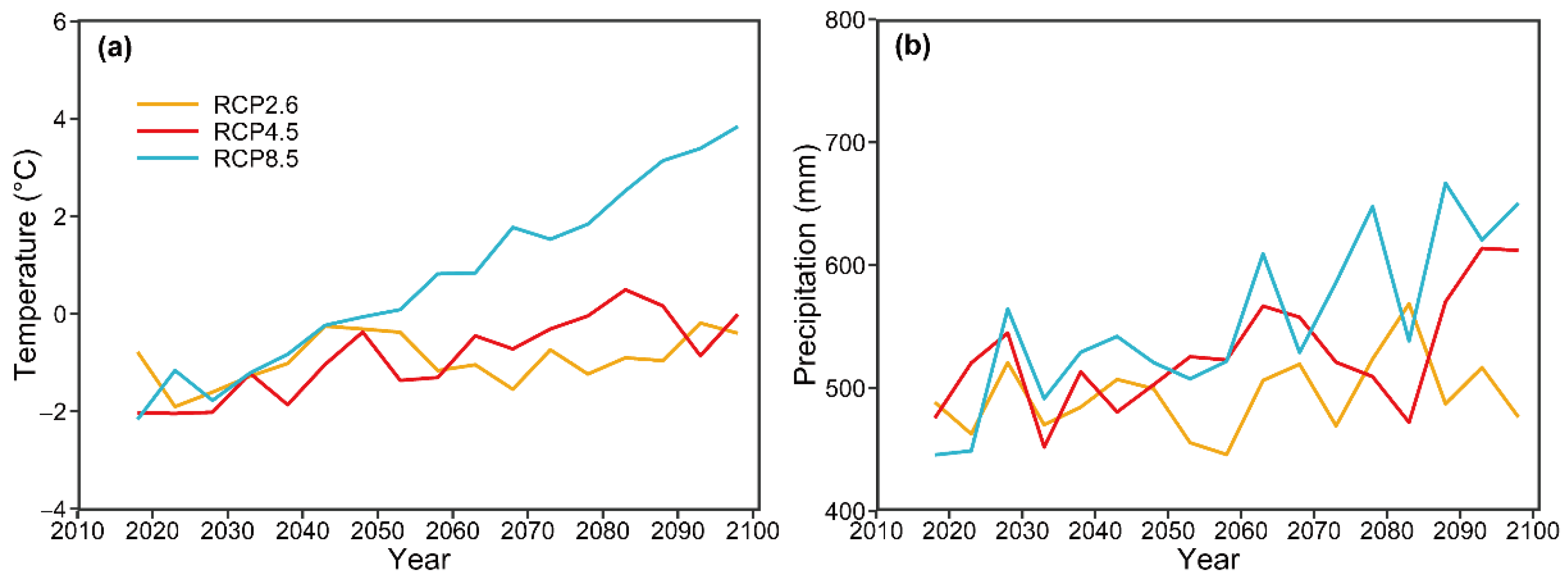
2 concentrations between the different GCMs (RCP2.6, RCP4.5, and RCP8.5) on MAT and MAP were similar to the patterns in emission intensity; for example, MAT under PRC 8.5 (high emission intensity) exhibited the sharpest increase, followed by RCP4.5 (intermediate emission intensity) and RCP2.6 (low peak-and-decay emission intensity) (
Figure 3). This finding suggests that atmospheric CO
2 concentrations are positively correlated with MAT and MAP. The pattern of variation in the predicted number of trees, basal area, species diversity, and size diversity was similar to that in emissions, suggesting that emissions contributed to changes in these variables. For example, the sharpest increase in the predicted number of trees was observed under RCP8.5, followed by RCP4.5 and RCP2.6 (
Figure 6).
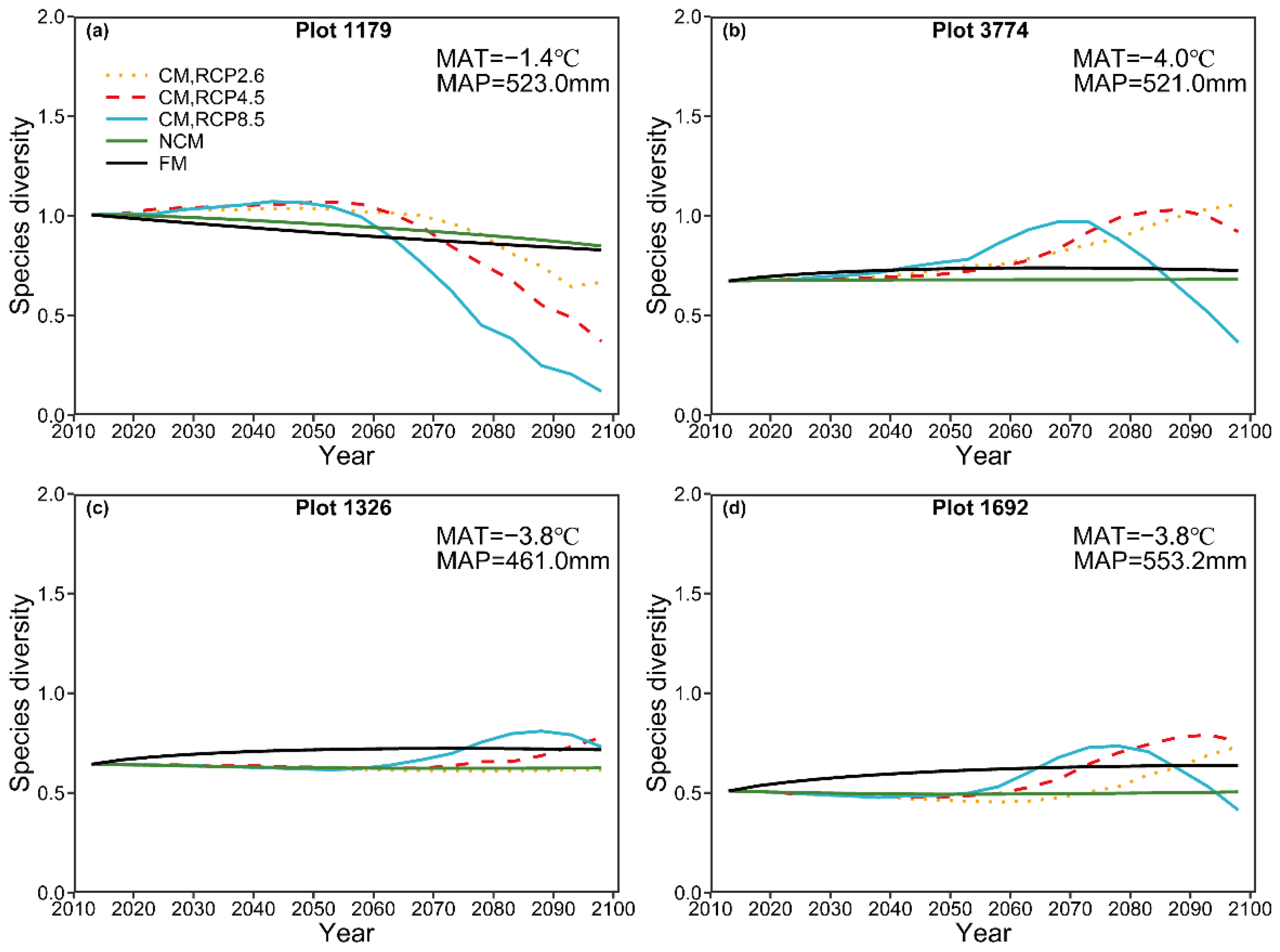
For long-term prediction, the predicted species diversity by the NCM, in general, showed almost no variation in time series, though a slightly decreasing trend was detected for plot 1179. By contrast, the species diversity predicted by the CM under these three climate change scenarios first shows an increasing pattern and then decreases, suggesting that species diversity could be negatively influenced by climate change. Many authors [
76,
77,
78,
79] have reported the results. For instance, Thuiller, et al. [
80] projected late 21st-century distributions for 1350 European plants species under seven climate change scenarios and found that more than half of European plant species could be vulnerable or threatened by 2080. Moreover, we observed that the predicted patterns of species diversity shared similar tendencies as emissions. The long-term predicted size diversity by the CM under three RCPs shared the same trend with the predicted species diversity, indicating climate change (increasing CO
2) can reduce tree size diversity and the decreasing trend is dependent on emissions.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








