
Climate Change Effects on Height–Diameter Allometric Relationship Vary with Tree Species and Size for Larch Plantations in Northern and Northeastern China


Abstract
:1. Introduction
2. Materials and Methods
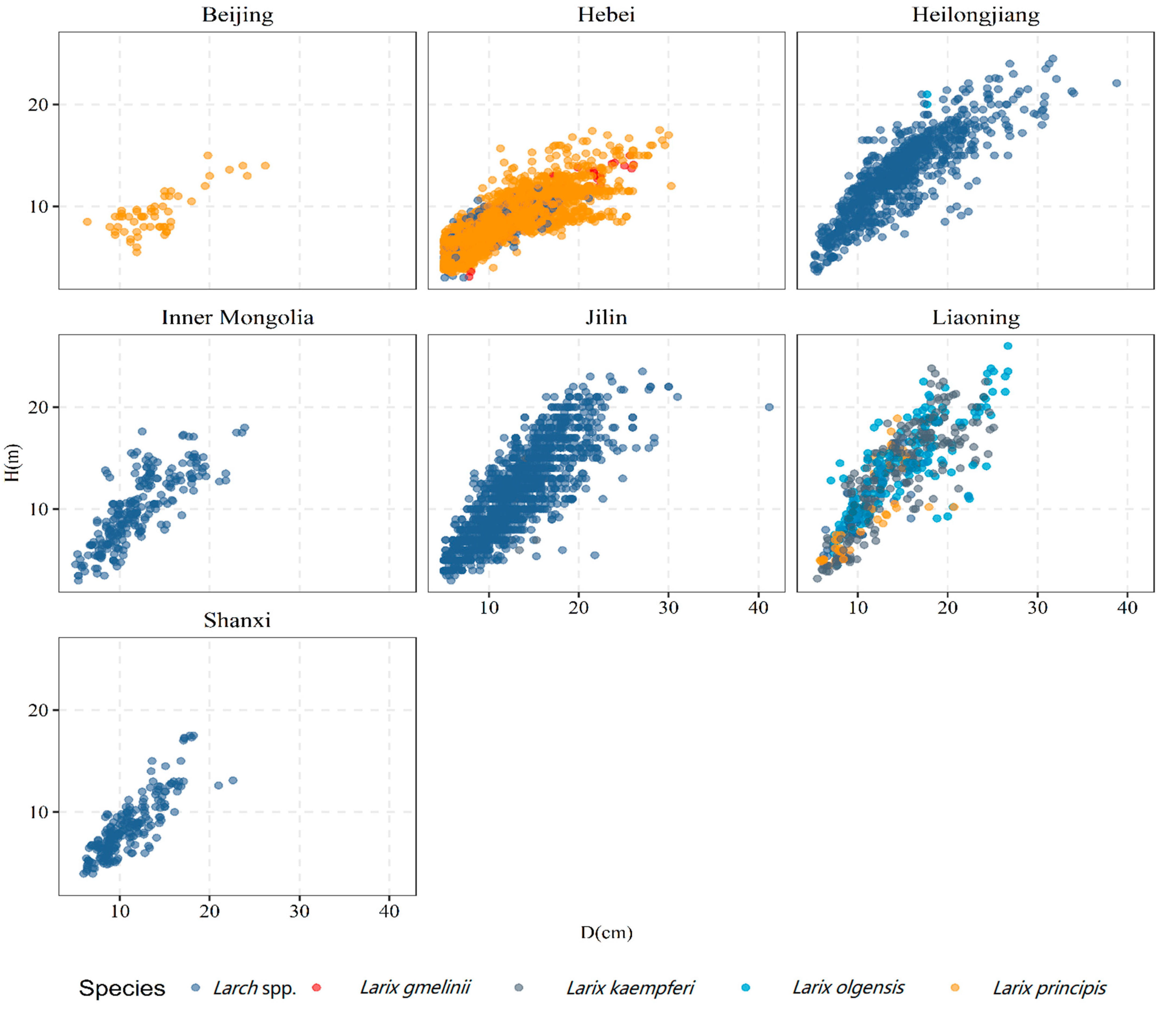
2.1. Tree Height–Diameter Data
2.2. Climate Data
2.3. Selection of Climate Variables
2.4. Basic H–D Models
2.5. Nonlinear Mixed-Effect Climate-Sensitive H–D Model
2.6. Model Evaluation and Validation
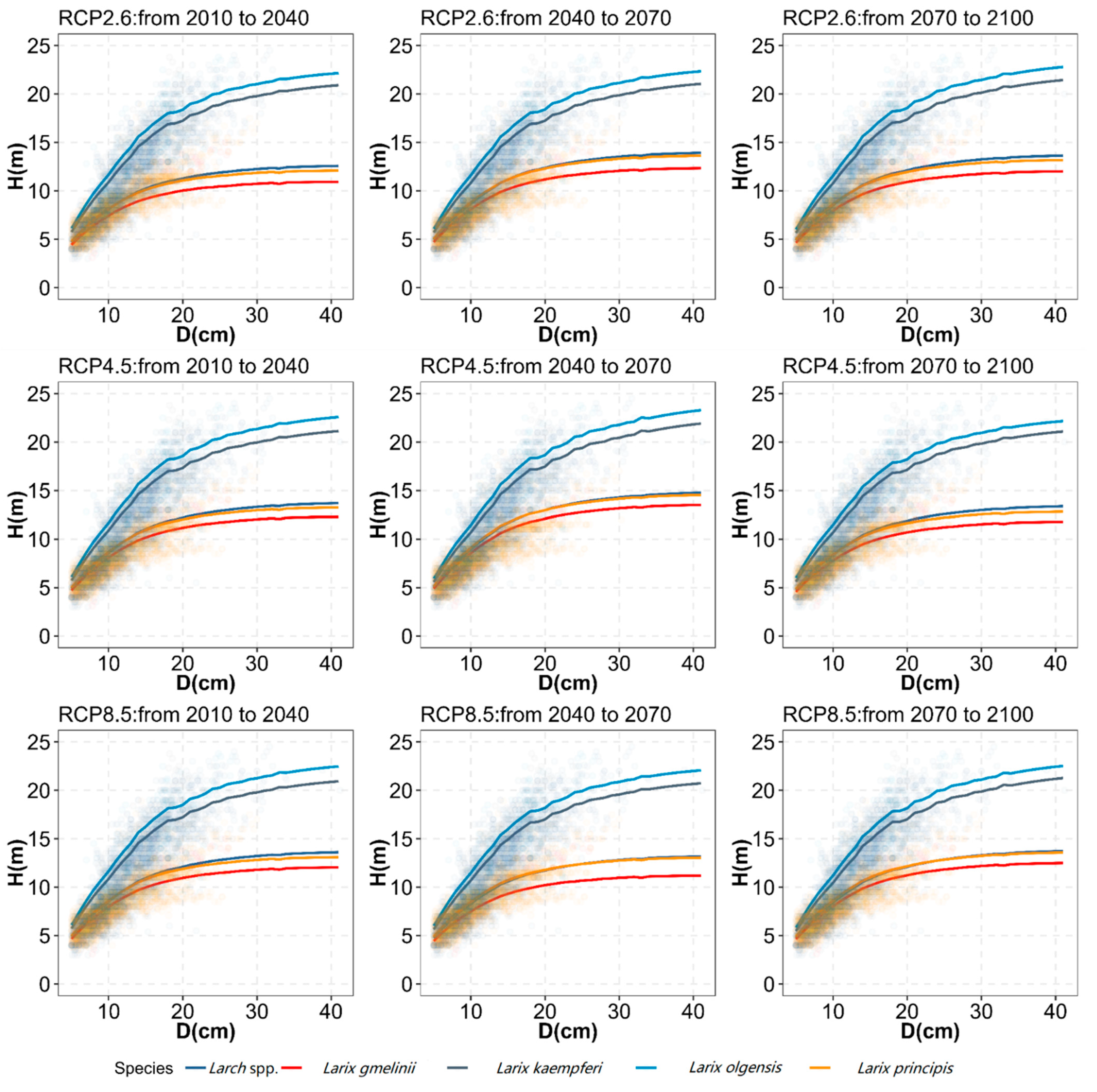
2.7. Comparisons of H–D Relationships among Larch Species under Future Climate Change
3. Results
3.1. Selected Climate Variables
3.2. Final NLME H–D Model with Climatic Variables
3.3. Model Comparison and Evaluation
3.4. H–D Relationships among Larch Tree Species and Tree Sizes under Future Climate Change
4. Discussion
4.1. Climate-Sensitive H–D Model
4.2. The Impact of Climate Change on H–D Relationship by Larch Species and Tree Size
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zell, J. SwissStandSim: A Climate Sensitive Single TREE Stand Simulator for Switzerland: Schlussbericht im Forschungsprogramm Waldund Klimawandel; Swiss Federal Institute of Forest, Snow and Landscape Research: Birmensdorf, Switzerland, 2016. [Google Scholar]
- Fang, Z.; Bailey, R. Height–diameter models for tropical forests on Hainan Island in southern China. For. Ecol. Manag. 1998, 110, 315–327. [Google Scholar] [CrossRef]
- Huang, S.; Price, D.; Titus, S.J. Development of ecoregion-based height–diameter models for white spruce in boreal forests. For. Ecol. Manag. 2000, 129, 125–141. [Google Scholar] [CrossRef]
- Jayaraman, K.; Zakrzewski, W. Practical approaches to calibrating height–diameter relationships for natural sugar maple stands in Ontario. For. Ecol. Manag. 2001, 148, 169–177. [Google Scholar] [CrossRef]
- Calama, R.; Montero, G. Interregional nonlinear height diameter model with random coefficients for stone pine in Spain. Can. J. For. Res. 2004, 34, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Shu, Y.Z. Height–diameter models using stand characteristics for Pinus banksiana and Picea mariana. Scand. J. For. Res. 2004, 19, 442–451. [Google Scholar] [CrossRef]
- Sharma, M.; Parton, J. Height–diameter equations for boreal tree species in Ontario using a mixed-effects modeling approach. For. Ecol. Manag. 2007, 249, 187–198. [Google Scholar] [CrossRef]
- Kroon, J.; Andersson, B.; Mullin, T.J. Genetic variation in the diameter–height relationship in Scots pine (Pinus sylvestris). Can. J. For. Res. 2008, 38, 1493–1503. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Swenson, N.G.; Weiser, M.D. Tree height–diameter allometry across the United States. Ecol. Evol. 2015, 5, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- Zang, H.; Lei, X.; Zeng, W. Height–diameter equations for larch plantations in northern and northeastern China: A comparison of the mixed-effects, quantile regression and generalized additive models. For. Int. J. For. Res. 2016, 89, 434–445. [Google Scholar] [CrossRef]
- Zhang, X.; Chhin, S.; Fu, L.; Lu, L.; Duan, A.; Zhang, J. Climate-sensitive tree height–diameter allometry for Chinese fir in southern China. For. Int. J. For. Res. 2019, 92, 167–176. [Google Scholar] [CrossRef]
- Bronisz, K.; Mehtätalo, L. Mixed-effects generalized height–diameter model for young silver birch stands on post-agricultural lands. For. Ecol. Manag. 2020, 460, 117901. [Google Scholar] [CrossRef]
- Ciceu, A.; Garcia-Duro, J.; Seceleanu, I.; Badea, O. A generalized nonlinear mixed-effects height–diameter model for Norway spruce in mixed-uneven aged stands. For. Ecol. Manag. 2020, 477, 118507. [Google Scholar] [CrossRef]
- Santiago-García, W.; Jacinto-Salinas, A.H.; Rodríguez-Ortiz, G.; Nava-Nava, A.; Santiago-García, E.; Ángeles-Pérez, G.; Enríquez-del Valle, J.R. Generalized height-diameter models for five pine species at Southern Mexico. For. Sci. Technol. 2020, 16, 49–55. [Google Scholar] [CrossRef]
- Zhang, B.; Sajjad, S.; Chen, K.; Zhou, L.; Zhang, Y.; Yong, K.K.; Sun, Y. Predicting Tree Height-Diameter Relationship from Relative Competition Levels Using Quantile Regression Models for Chinese Fir (Cunninghamia lanceolata) in Fujian Province, China. Forests 2020, 11, 183. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C.A.L.; Varela, J.G.; Dorado, F.C.; Alboreca, A.R.; Soalleiro, R.R.; González, J.G.Á.; Rodríguez, F.S. A height-diameter model for Pinus radiata D. Don in Galicia (Northwest Spain). Ann. For. Sci. 2003, 60, 237–245. [Google Scholar] [CrossRef]
- Krisnawati, H.; Wang, Y.; Ades, P.K. Generalized height-diameter models for Acacia mangium Willd. plantations in south Sumatra. Indones. J. For. Res. 2010, 7, 1–19. [Google Scholar]
- Saunders, M.R.; Wagner, R.G. Height-diameter models with random coefficients and site variables for tree species of Central Maine. Ann. For. Sci. 2008, 65, 203. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.B.; Amateis, R.L.; Burkhart, H.E. Implementing regional locale and thinning response in the loblolly pine height-diameter relationship. South. J. Appl. For. 2010, 34, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fang, J.; Tang, Z.; Zhu, B. Climatic control of primary forest structure and DBH–height allometry in Northeast China. For. Ecol. Manag. 2006, 234, 264–274. [Google Scholar] [CrossRef]
- Fortin, M.; Van Couwenberghe, R.; Perez, V.; Piedallu, C. Evidence of climate effects on the height-diameter relationships of tree species. Ann. For. Sci. 2019, 76, 1. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, M.U. Forest growth and species distribution in a changing climate. Tree Physiol. 2000, 20, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Watanabe, M.; Li, F.; Zhang, J.; Zhang, W.; Zhai, J. Factors affecting forest growth and possible effects of climate change in the Taihang Mountains, northern China. For. Int. J. For. Res. 2006, 79, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Hartl-Meier, C.; Dittmar, C.; Zang, C.; Rothe, A. Mountain forest growth response to climate change in the Northern Limestone Alps. Trees 2014, 28, 819–829. [Google Scholar] [CrossRef]
- Charney, N.D.; Babst, F.; Poulter, B.; Record, S.; Trouet, V.M.; Frank, D.; Enquist, B.J.; Evans, M.E. Observed forest sensitivity to climate implies large changes in 21st century North American forest growth. Ecol. Lett. 2016, 19, 1119–1128. [Google Scholar] [CrossRef]
- Albert, M.; Schmidt, M. Climate-sensitive modelling of site-productivity relationships for Norway spruce (Picea abies (L.) Karst.) and common beech (Fagus sylvatica L.). For. Ecol. Manag. 2010, 259, 739–749. [Google Scholar] [CrossRef]
- Ng’andwe, P.; Chungu, D.; Tailoka, F. Stand characteristics and climate modulate height to diameter relationship in Pinus merkusii and P. michoacana in Zambia. Agric. For. Meteorol. 2021, 307, 108510. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Banin, L.; Phillips, O.L.; Baker, T.R.; Lewis, S.L.; Quesada, C.A.; Affum-Baffoe, K.; Arets, E.J.; Berry, N.J.; Bird, M. Height-diameter allometry of tropical forest trees. Biogeosciences 2011, 8, 1081–1106. [Google Scholar] [CrossRef] [Green Version]
- State Forestry Administration, The People’s Republic of China. National Forest Resources Statistics (2009–2013); State Forestry Administration: Beijing, China, 2014; p. 233.
- Shen, C.; Lei, X.; Liu, H.; Wang, L.; Liang, W. Potential impacts of regional climate change on site productivity of Larix olgensis plantations in northeast China. ifor.-Biogeosci. For. 2015, 8, 642. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wang, H.; Lei, X. Application of the 3-PG model to predict growth of Larix olgensis plantations in northeastern China. For. Ecol. Manag. 2017, 406, 208–218. [Google Scholar] [CrossRef]
- Xie, Y.; Lei, X.; Shi, J. Impacts of climate change on biological rotation of Larix olgensis plantations for timber production and carbon storage in northeast China using the 3-PGmix model. Ecol. Model. 2020, 435, 109267. [Google Scholar] [CrossRef]
- Lei, X.; Yu, L.; Hong, L. Climate-sensitive integrated stand growth model (CS-ISGM) of Changbai larch (Larix olgensis) plantations. For. Ecol. Manag. 2016, 376, 265–275. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Innes, J.L.; Seely, B.; Chen, B. ClimateAP: An application for dynamic local downscaling of historical and future climate data in Asia Pacific. Front. Agric. Sci. Eng. 2017, 4, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5. [Google Scholar] [CrossRef]
- Voldoire, A.; Sanchez-Gomez, E.; y Mélia, D.S.; Decharme, B.; Cassou, C.; Sénési, S.; Valcke, S.; Beau, I.; Alias, A.; Chevallier, M. The CNRM-CM5.1 global climate model: Description and basic evaluation. Clim. Dyn. 2013, 40, 2091–2121. [Google Scholar] [CrossRef] [Green Version]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Scolforo, J.R.S.; Maestri, R.; Ferraz Filho, A.C.; de Mello, J.M.; de Oliveira, A.D.; de Assis, A.L. Dominant height model for site classification of Eucalyptus grandis incorporating climatic variables. Int. J. For. Res. 2013, 2013, 139236. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: Linear and nonlinear mixed effects models. R Package Version 2013, 3, 111. [Google Scholar]
- Sharma, R.; Vacek, Z.; Vacek, S. Nonlinear mixed effect height-diameter model for mixed species forests in the central part of the Czech Republic. J. For. Sci. 2016, 62, 470–484. [Google Scholar]
- Vizcaíno-Palomar, N.; Ibáñez, I.; Benito-Garzón, M.; González-Martínez, S.C.; Zavala, M.A.; Alía, R. Climate and population origin shape pine tree height-diameter allometry. New For. 2017, 48, 363–379. [Google Scholar] [CrossRef]
- Kilpeläinen, A.; Peltola, H.; Rouvinen, I.; Kellomäki, S. Dynamics of daily height growth in Scots pine trees at elevated temperature and CO2. Trees 2006, 20, 16–27. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Wang, G.; Wang, C.; Bai, T.; Lu, H.; Lv, H.; Chen, C.; Yuan, J.; Xu, Z.; et al. Larch Forest in China; China Forestry Publishing House: Beijing, China, 1992. [Google Scholar]
- Zhou, Y.; Lei, Z.; Zhou, F.; Han, Y.; Yu, D.; Zhang, Y. Impact of climate factors on height growth of Pinus sylvestris var. mongolica. PLoS ONE 2019, 14, e0213509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, Z.; Sebastian-Azcona, J.; Hamann, A.; Menzel, A.; Hacke, U. Adaptive limitations of white spruce populations to drought imply vulnerability to climate change in its western range. Evol. Appl. 2019, 12, 1850–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, H.; Chhin, S.; Zhang, J. Effects of competition, age and climate on tree slenderness of Chinese fir plantations in southern China. For. Ecol. Manag. 2020, 458, 117815. [Google Scholar] [CrossRef]
- Ryan, M.G.; Yoder, B. Hydraulic limits to tree height and tree growth. Bioscience 1997, 47, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Aiba, S.-i.; Kitayama, K. Structure, composition and species diversity in an altitude-substrate matrix of rain forest tree communities on Mount Kinabalu, Borneo. Plant Ecol. 1999, 140, 139–157. [Google Scholar] [CrossRef]
- Thornley, J.H. Modelling stem height and diameter growth in plants. Ann. Bot. 1999, 84, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Schelhaas, M. The wind stability of different silvicultural systems for Douglas-fir in the Netherlands: A model-based approach. Forestry 2008, 81, 399–414. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef]
- Campbell, E.M.; Magnussen, S.; Antos, J.A.; Parish, R. Size-, species-, and site-specific tree growth responses to climate variability in old-growth subalpine forests. Ecosphere 2021, 12, e03529. [Google Scholar] [CrossRef]
- Cannell, M.; Rothery, P.; Ford, E. Competition within stands of Picea sitchensis and Pinus contorta. Ann. Bot. 1984, 53, 349–362. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Province | Number of Plots | Number of Tree Observations | D (cm) | H (m) | AGE (a) | N (Trees × ha−1) | BA (m2 × ha−1) |
|---|---|---|---|---|---|---|---|---|
| Calibration | Beijing | 7 | 37 | 14.7 (4.6) | 9.3 (2.3) | 32.9 (10.6) | 561.9 (418.5) | 9.1 (9.9) |
| Hebei | 72 | 3326 | 10.7 (4.5) | 7.9 (2.2) | 21.7 (6.7) | 1072.6 (540.9) | 9.3 (7.3) | |
| Heilongjiang | 96 | 706 | 14.5 (5.3) | 13 (3.9) | 27.9 (9.7) | 653.9 (520.5) | 4.6 (4.6) | |
| Jilin | 132 | 1058 | 12.9 (4.7) | 11.4 (4.4) | 25.2 (9.9) | 1032.2 (565.3) | 9.0 (5.6) | |
| Liaoning | 52 | 406 | 14.4 (4.8) | 13.5 (4.5) | 23.6 (10.2) | 1292.3 (631.7) | 13.5 (8.6) | |
| Inner Mongolia | 35 | 188 | 12.2 (3.7) | 10.1 (3.2) | 25.8 (7.1) | 855.1 (617.7) | 8.0 (6.4) | |
| Shanxi | 34 | 195 | 11.1 (3.1) | 8.7 (2.8) | 26.5 (9.3) | 1297.8 (627.3) | 9.4 (7.7) | |
| total | 428 | 5916 | 11.9 (4.8) | 9.6 (3.8) | 23.6 (8.5) | 977.2 (610.6) | 8.5 (6.9) | |
| Validation | Beijing | 3 | 18 | 12.6 (1.7) | 9.2 (1.6) | 25.8 (3.1) | 816.8 (469.2) | 11.5 (10.7) |
| Hebei | 12 | 620 | 11.4 (3.8) | 8.9 (2.3) | 24.8 (7.6) | 994.9 (607.2) | 8.5 (5.4) | |
| Heilongjiang | 27 | 193 | 15.2 (5.0) | 13.1 (3.8) | 29.5 (9.8) | 559.3 (548.7) | 6.0 (6.3) | |
| Jilin | 38 | 351 | 13.5 (4.4) | 11.9 (4.0) | 28.4 (11.1) | 904.2 (532.9) | 8.2 (4.9) | |
| Liaoning | 12 | 97 | 12.2 (4.5) | 11.3 (5.4) | 23.2 (9.3) | 1207.8 (651.1) | 17.7 (7.0) | |
| Inner Mongolia | 11 | 53 | 11.1 (4.4) | 9.1 (4.1) | 28 (9.9) | 1135.5 (835.4) | 7.3 (6.4) | |
| Shanxi | 4 | 21 | 9 (1.8) | 6.7 (2.0) | 17.9 (4.2) | 848.6 (622.8) | 7.3 (6.3) | |
| total | 107 | 1353 | 12.5 (4.4) | 10.4 (3.8) | 26.3 (9.4) | 902.6 (626.8) | 8.6 (6.8) |
| Variable | Description |
|---|---|
| AHM | Annual heat:moisture index |
| CMD | Hargreaves climatic moisture deficit |
| DD_0 | Degree-days below 0 °C |
| DD_18 | Degree-days below 18 °C |
| DD18 | Degree-days above 18 °C |
| DD5 | Degree-days above 5 °C |
| EMT/°C | Extreme minimum temperature over a 30-year period |
| EXT/°C | Extreme maximum temperature over a 30-year period |
| EREF | Hargreaves reference evaporation |
| MAP/mm | Mean annual precipitation |
| MAT/°C | Mean annual temperature |
| MCMT/°C | Mean coldest month temperature |
| MWMT/°C | Mean warmest month temperature |
| NFFD | The number of frost-free days |
| PAS/mm | Precipitation as snow between August in previous year and July in current year |
| TD/°C | Temperature difference between MWMT and MCMT, or continentality |
| Comp1 | Comp2 | Comp3 | |
|---|---|---|---|
| MAT | 0.331 | 0.000 | 0.000 |
| MWMT | 0.265 | 0.248 | −0.229 |
| MCMT | 0.282 | −0.172 | 0.272 |
| TD | −0.114 | 0.341 | −0.43 |
| MAP | 0.000 | 0.394 | 0.338 |
| AHM | 0.147 | −0.387 | −0.262 |
| DD_0 | −0.302 | 0.104 | −0.238 |
| DD5 | 0.301 | 0.185 | −0.14 |
| DD_18 | −0.329 | 0.000 | −0.104 |
| DD18 | 0.269 | 0.243 | −0.183 |
| NFFD | 0.311 | 0.150 | 0.000 |
| PAS | −0.172 | 0.265 | 0.388 |
| EMT | 0.287 | −0.128 | 0.21 |
| EXT | 0.250 | 0.171 | −0.357 |
| Eref | 0.249 | −0.215 | 0.000 |
| CMD | 0.000 | −0.444 | −0.238 |
| Accumulated variance | 56.050 | 81.680 | 95.270 |
| Variables | CMD | TD | PAS | MAP | DD_18 | MAT | H |
|---|---|---|---|---|---|---|---|
| CMD | 1.000 | - | - | - | - | - | - |
| TD | −0.424 *** | 1.000 | - | - | - | - | - |
| PAS | −0.673 *** | 0.193 *** | 1.000 | - | - | - | - |
| MAP | −0.841 *** | 0.141 *** | 0.566 *** | 1.000 | - | - | - |
| DD_18 | −0.041 *** | 0.435 *** | 0.421 *** | −0.358 *** | 1.000 | - | - |
| MAT | −0.004 | −0.347 *** | −0.412 *** | 0.390 *** | −0.995 *** | 1.000 | - |
| H | −0.503 *** | 0.404 *** | 0.329 *** | 0.414 *** | 0.070 *** | −0.029 *** | 1.000 |
| MAT | Period: 2010–2040 | Period: 2040–2070 | Period: 2070–2100 |
| RCP2.6 | 3.95 (2.40) | 4.41 (2.41) | 4.53 (2.43) |
| RCP4.5 | 3.92 (2.43) | 4.87 (2.44) | 5.70 (2.42) |
| RCP8.5 | 4.14 (2.42) | 5.69 (2.42) | 7.44 (2.36) |
| CMD | Period: 2010–2040 | Period: 2040–2070 | Period: 2070–2100 |
| RCP2.6 | 180.96 (88.22) | 164.38 (75.68) | 164.34 (87.09) |
| RCP4.5 | 158.82 (82.30) | 142.16 (77.02) | 180.26 (83.53) |
| RCP8.5 | 160.62 (83.16) | 186.03 (82.88) | 187.04 (82.78) |
| Parameter | Parameter Definition | Equation (13) | Equation (14) | Equation (15) | |
|---|---|---|---|---|---|
| Fixed-effects parameters | a0 | 21.382 (0.000) | 21.460 (0.000) | 19.772 (0.000) | |
| b0 | 0.078 (0.000) | 0.088 (0.000) | 0.106 (0.000) | ||
| c0 | 1.616 (0.000) | 1.545 (0.000) | 2.083 (0.000) | ||
| a1 | BAL | −0.137 (0.000) | −0.085 (0.000) | −0.111 (0.000) | |
| a2 | MAT | 1.322 (0.000) | 0.259 (0.0381) | ||
| a3 | CMD | −0.021 (0.000) | −0.030 (0.000) | ||
| b1 | BAL | 0.001 (0.000) | 0.001 (0.000) | 0.005 (0.000) | |
| b2 | MAT | −0.005 (0.000) | 0.003 (0.0024) | ||
| b3 | CMD | 0.000 (0.07) | 0.000 (0.000) | ||
| g1 | L. gmelinii | −5.137 (0.000) | −3.790 (0.000) | 0.216 (0.898) | |
| g2 | L. olgensis | 3.234 (0.000) | 3.317 (0.000) | 1.879 (0.013) | |
| g3 | L. kaempferi | −4.944 (0.000) | −2.951 (0.000) | 0.865 (0.050) | |
| g4 | L. principis | 2.557 (0.000) | 2.319 (0.000) | 0.226 (0.735) | |
| Variance components | 1.349 | ||||
| 2.700 | |||||
| Model performance | |||||
| 0.674 | |||||
| AIC | 23,007.83 | 22,368 | 17,911.4 | ||
| 0.77 | 0.79 | 0.92 | |||
| 1.37 | 1.28 | 0.76 | |||
| 1.82 | 1.72 | 1.06 | |||
| 1.5 | 1.44 | 1.38 | |||
| 2.03 | 1.93 | 1.8 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.; Lei, X.; Zang, H.; Zeng, W. Climate Change Effects on Height–Diameter Allometric Relationship Vary with Tree Species and Size for Larch Plantations in Northern and Northeastern China. Forests 2022, 13, 468. https://doi.org/10.3390/f13030468
Xu Q, Lei X, Zang H, Zeng W. Climate Change Effects on Height–Diameter Allometric Relationship Vary with Tree Species and Size for Larch Plantations in Northern and Northeastern China. Forests. 2022; 13(3):468. https://doi.org/10.3390/f13030468
Chicago/Turabian StyleXu, Qigang, Xiangdong Lei, Hao Zang, and Weisheng Zeng. 2022. "Climate Change Effects on Height–Diameter Allometric Relationship Vary with Tree Species and Size for Larch Plantations in Northern and Northeastern China" Forests 13, no. 3: 468. https://doi.org/10.3390/f13030468