
Global Warming Favors the Development of a Rich and Heterogeneous Mycobiota on Alien Vines in a Boreal City under Continental Climate

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
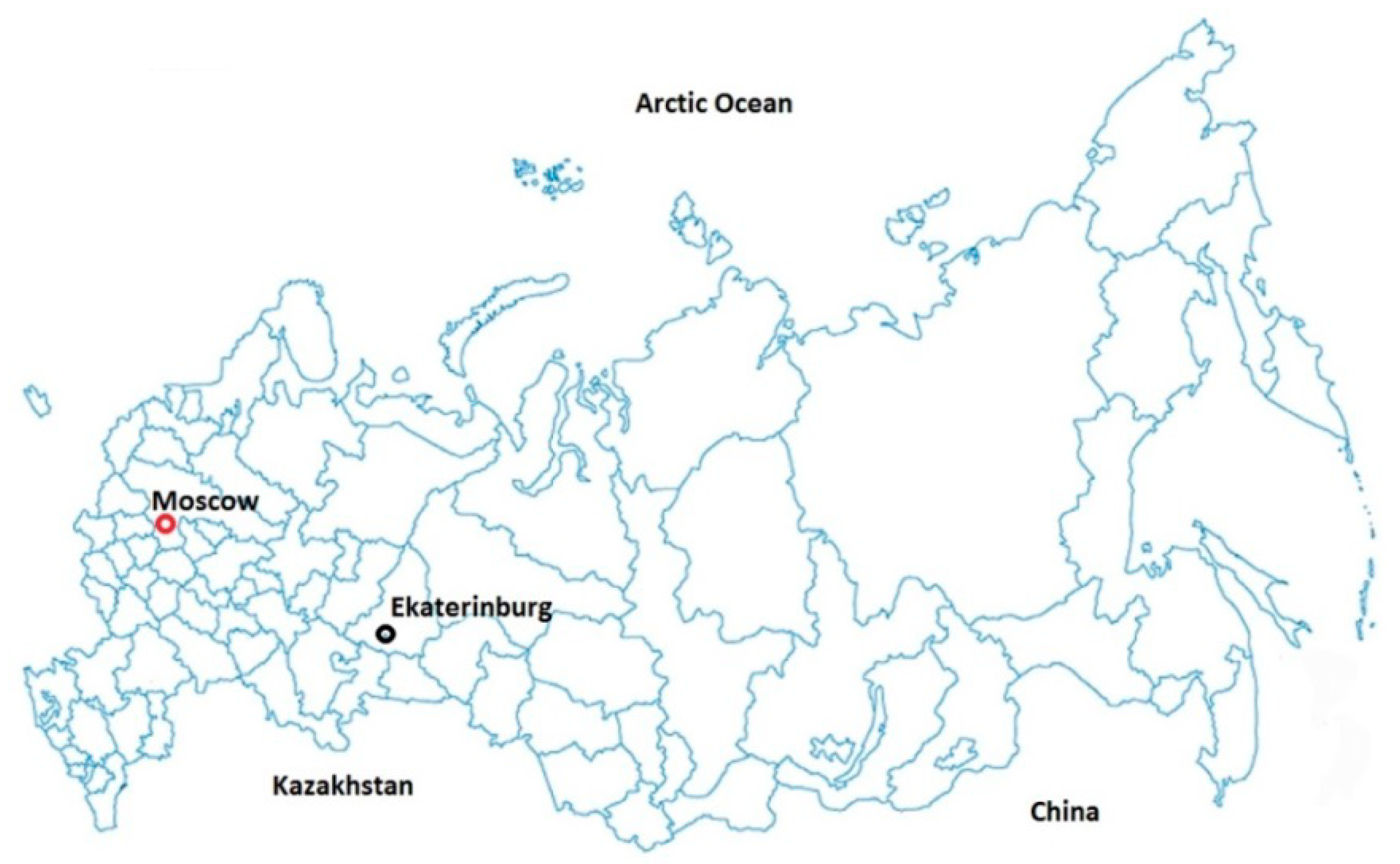
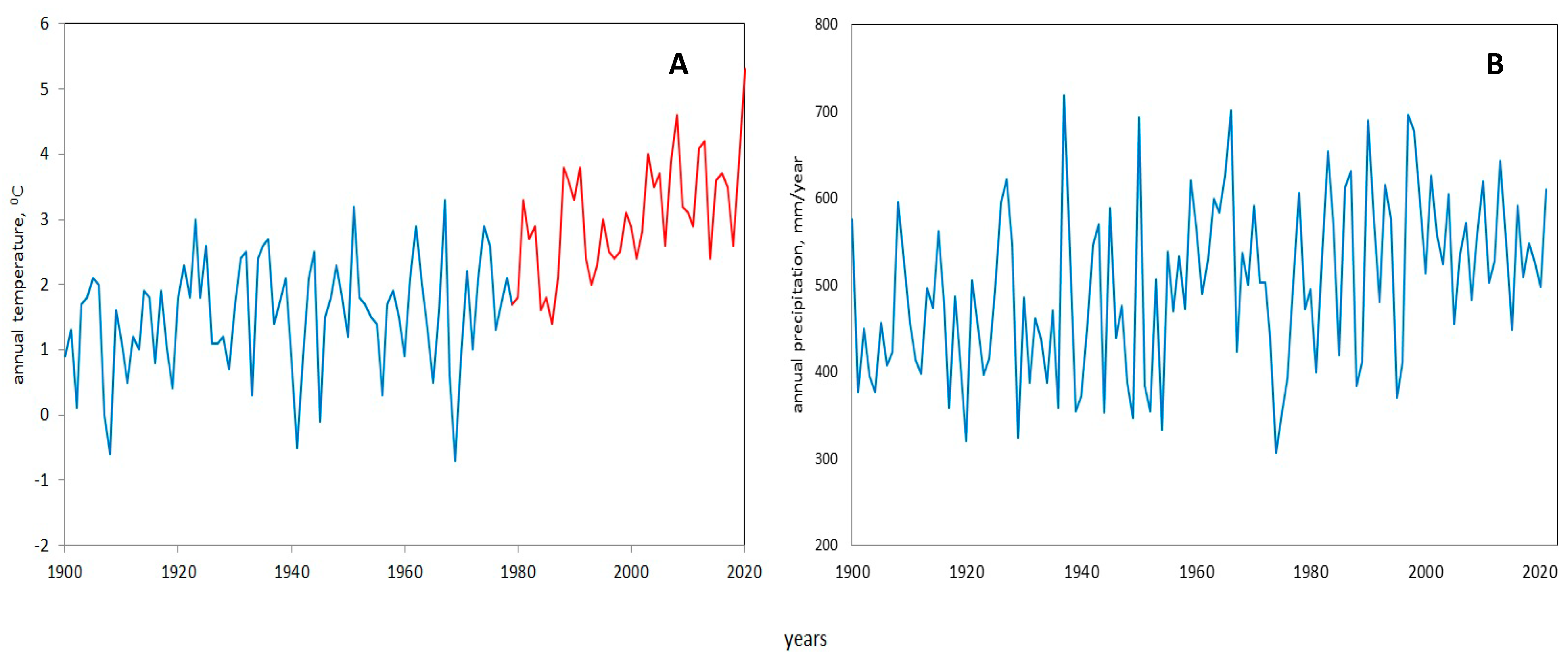
2.1. Climate of Ekaterinburg City
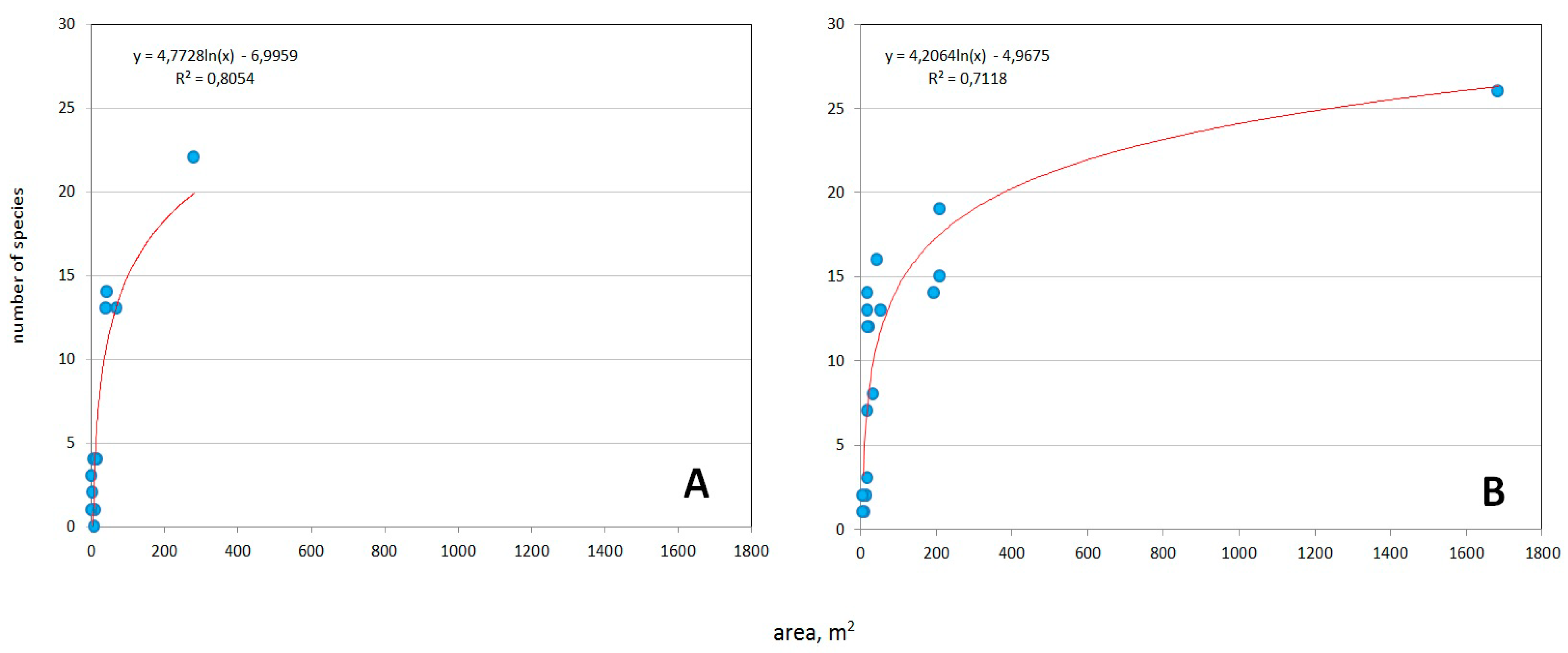
2.2. Systematic Review of Yield Dynamics and Assessment of Vines Covered Areas
2.3. Fungal Sampling
2.4. Data Analysis
3. Results
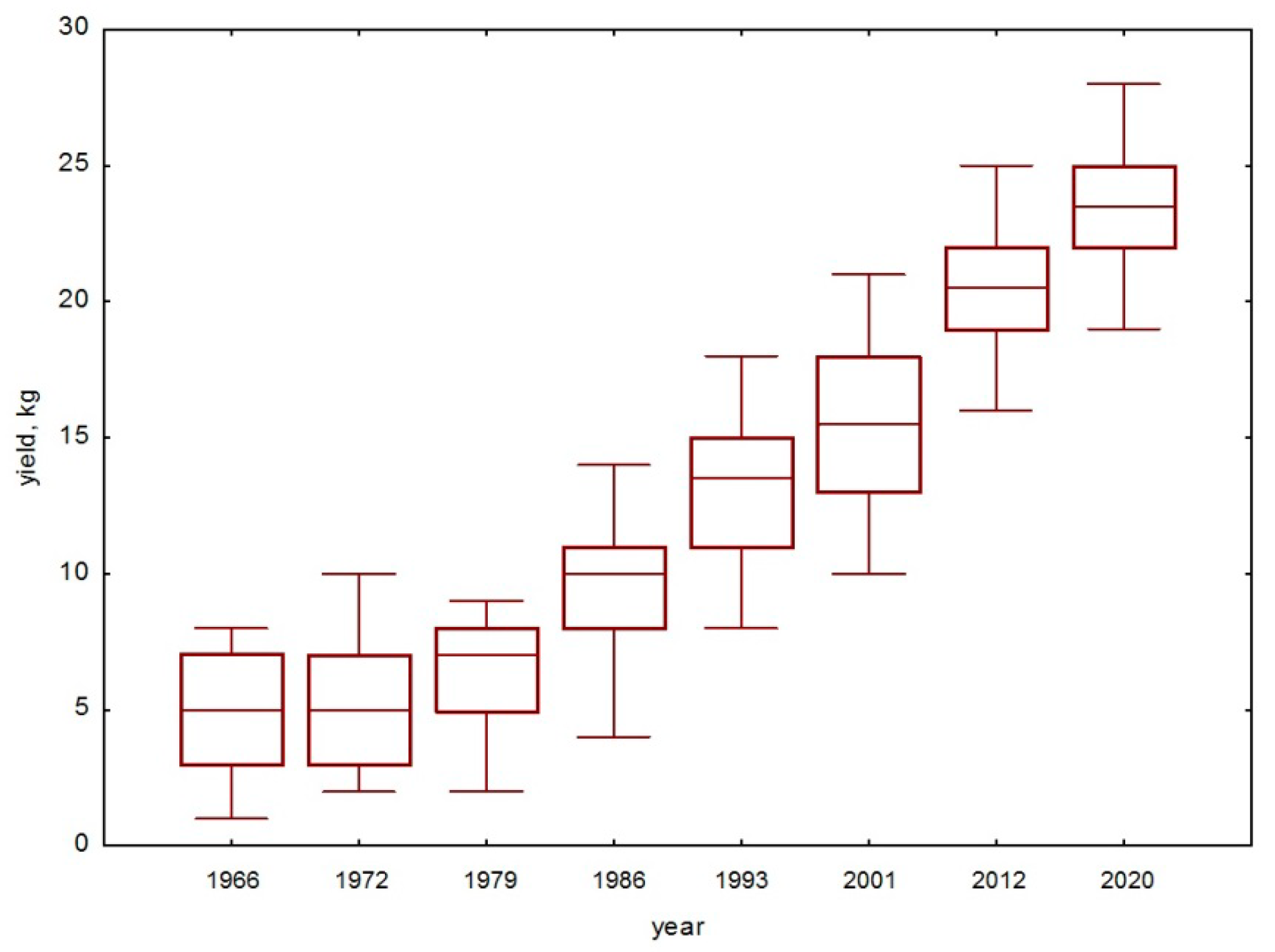
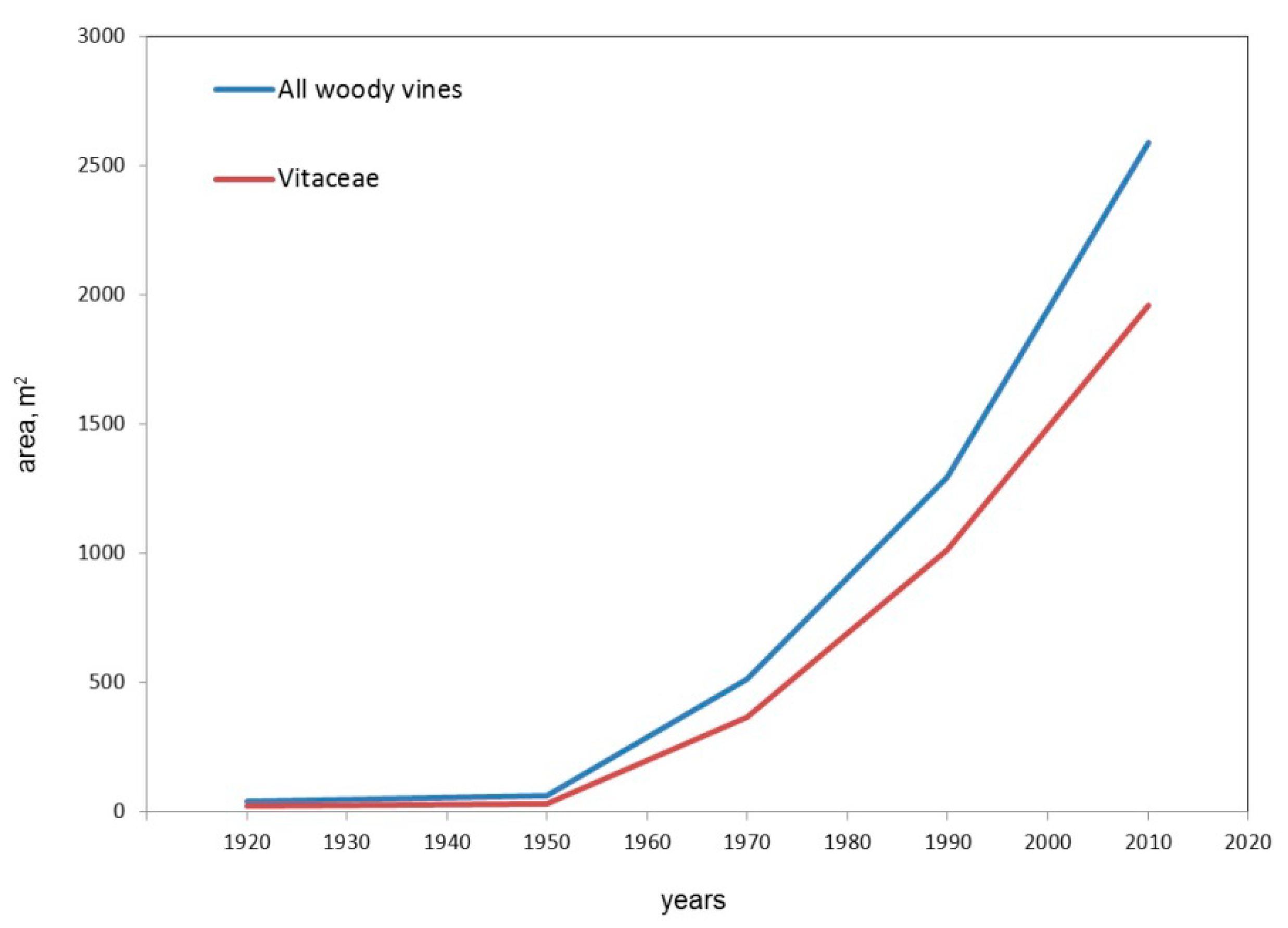
3.1. Dynamics of Grape Yield and Woody Vines Areas in Ekaterinburg City
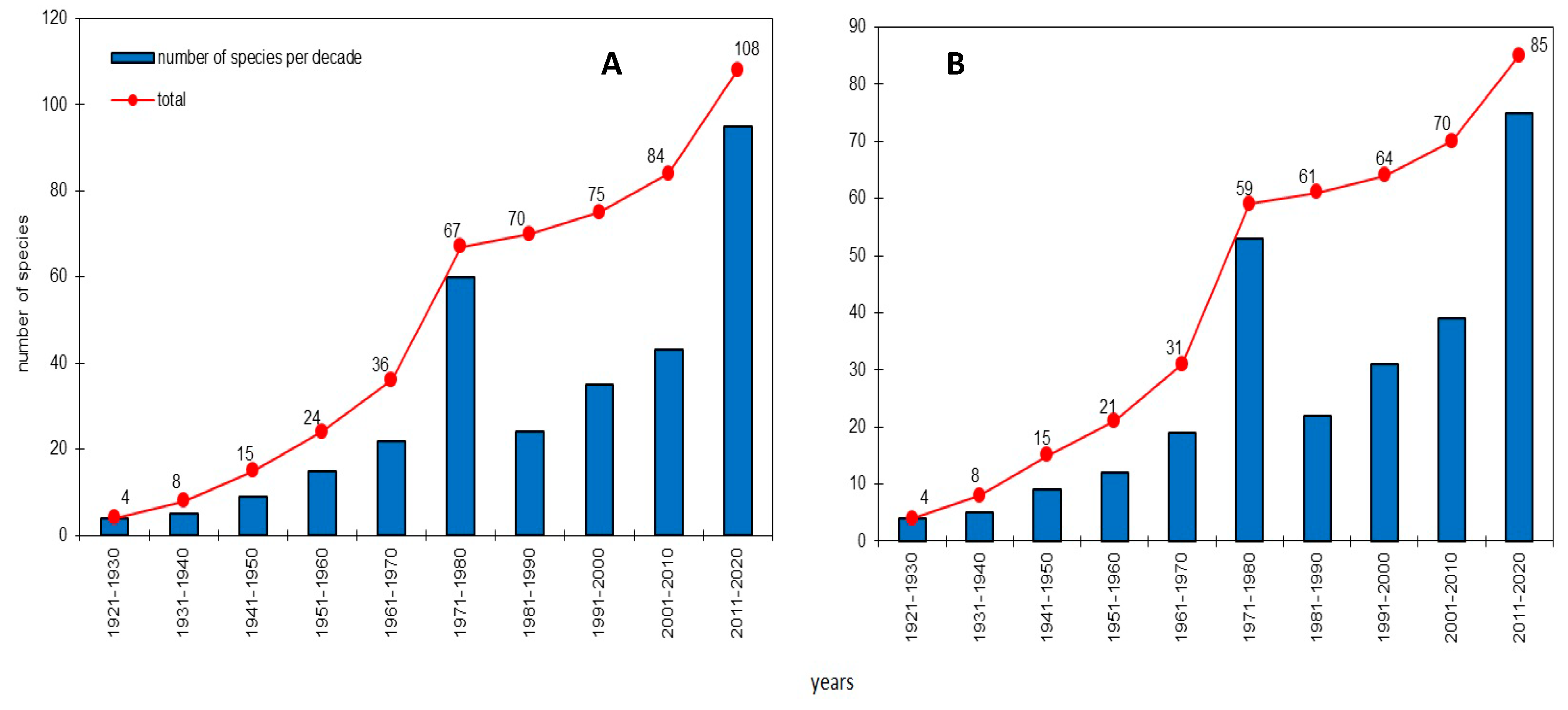
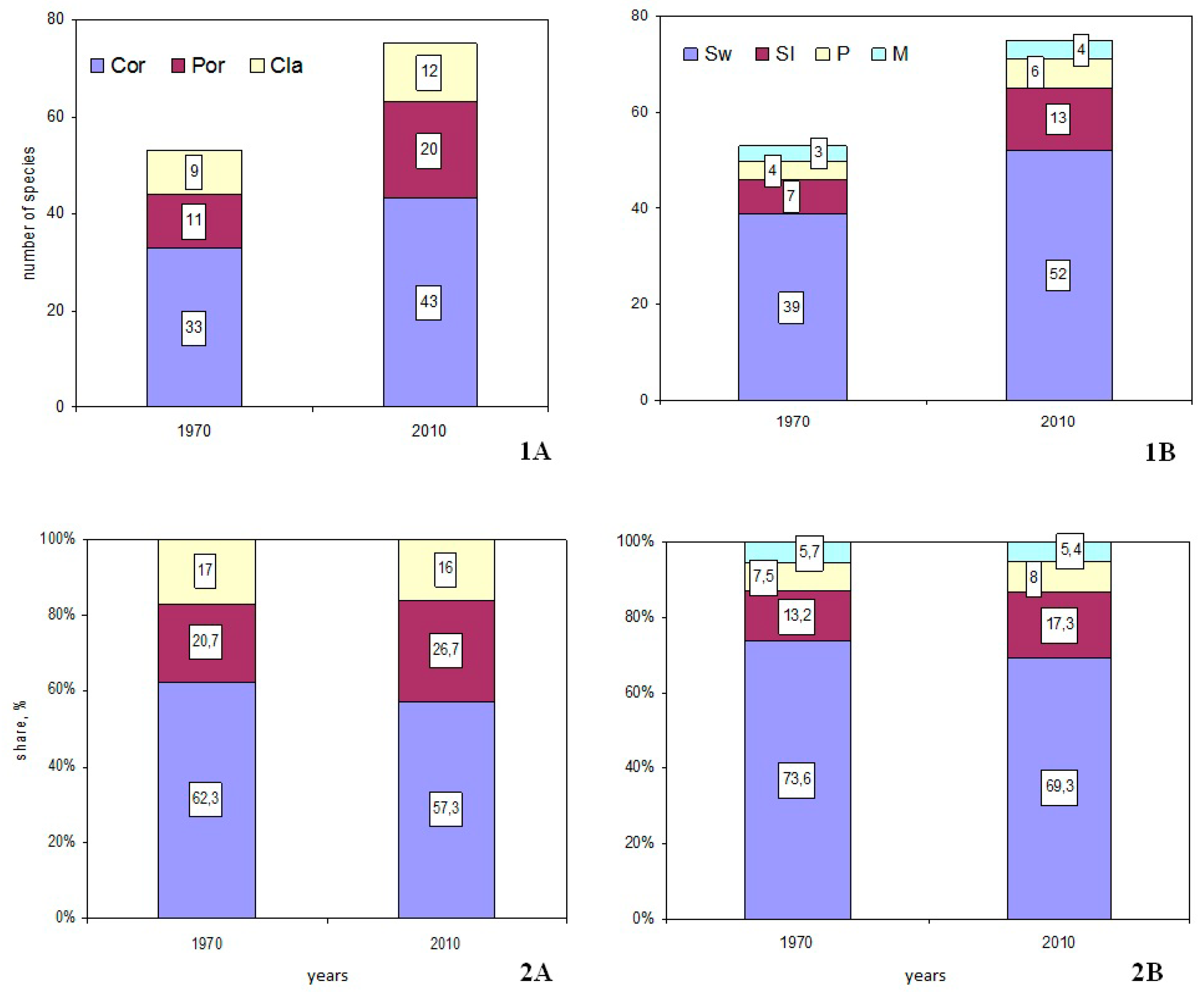
3.2. Systematic Review of the Fungal Species Richness Revealed during the Last 100 Years
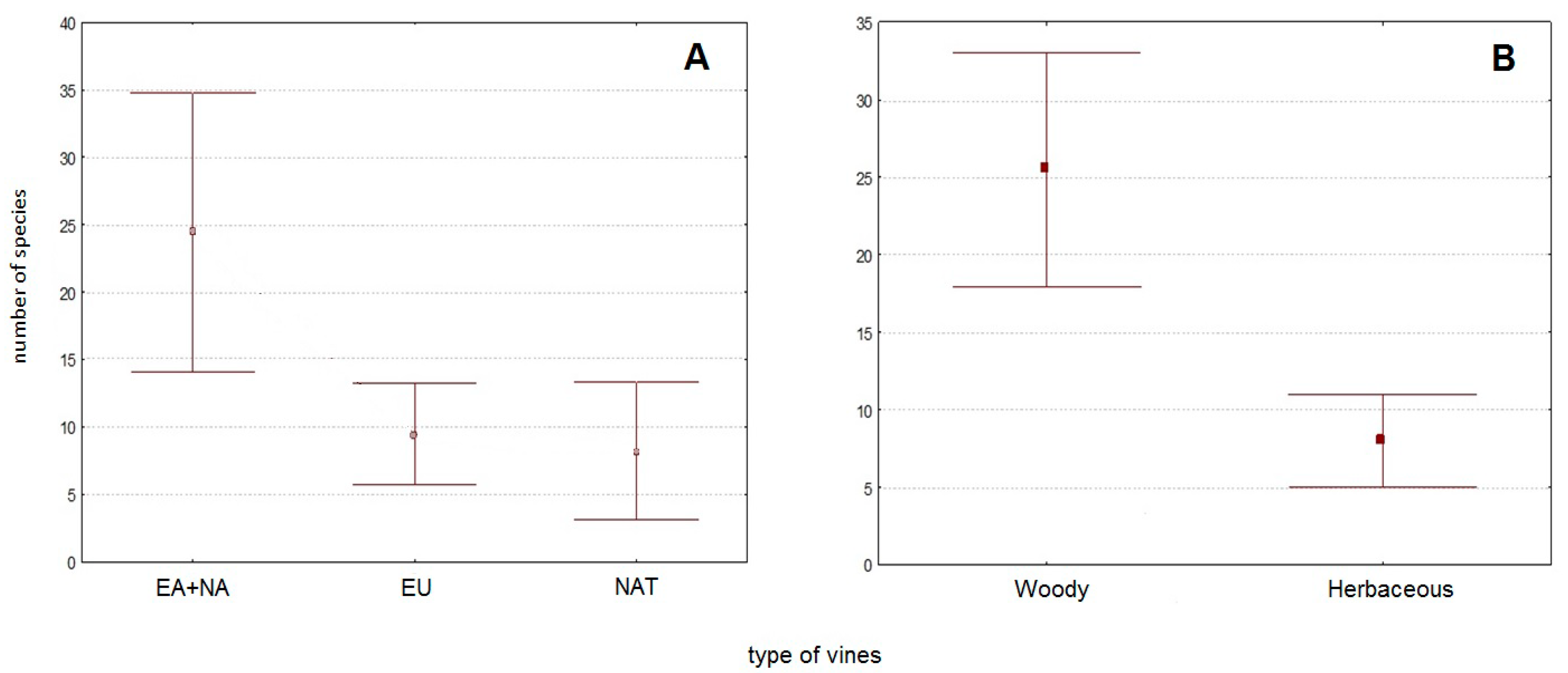
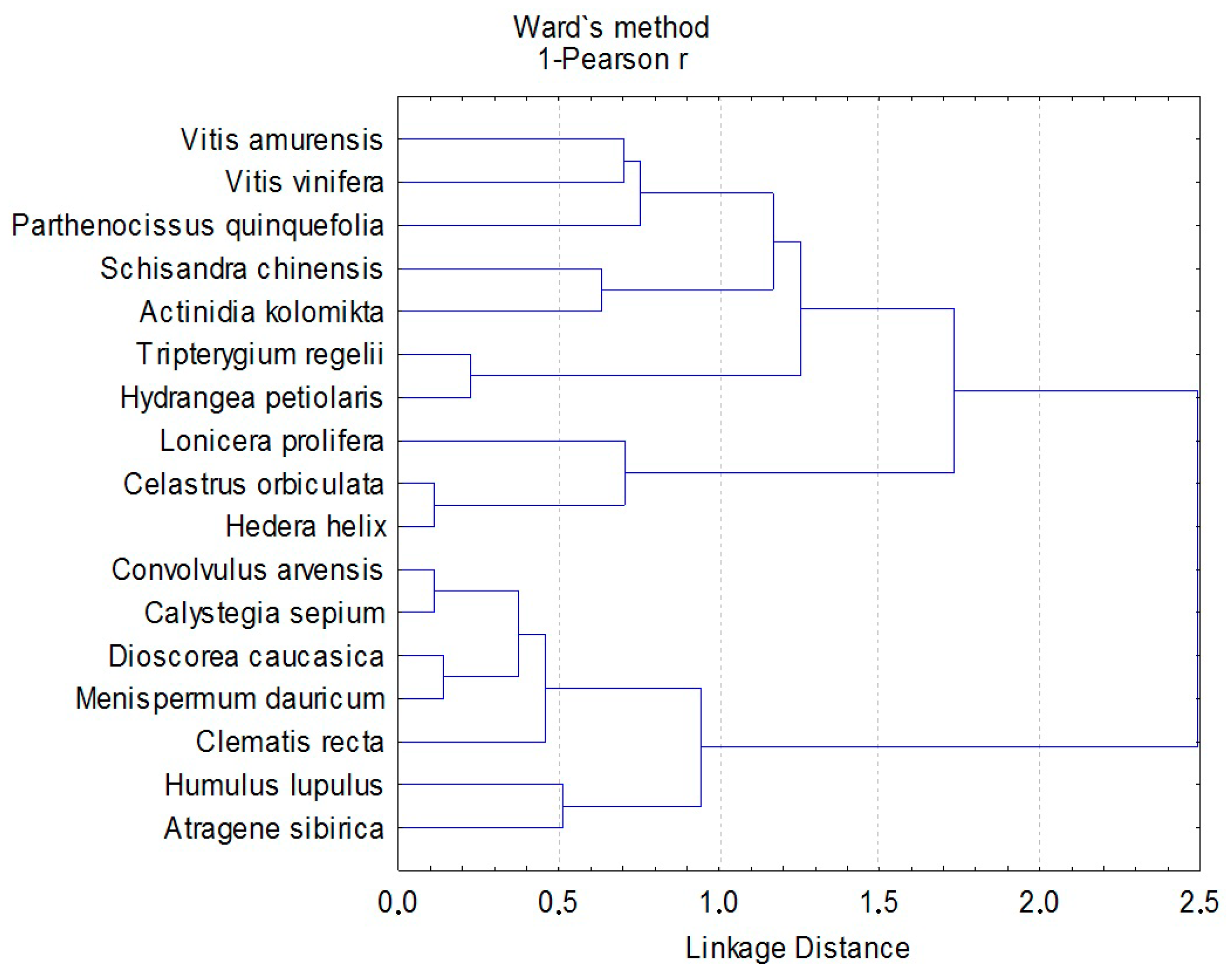
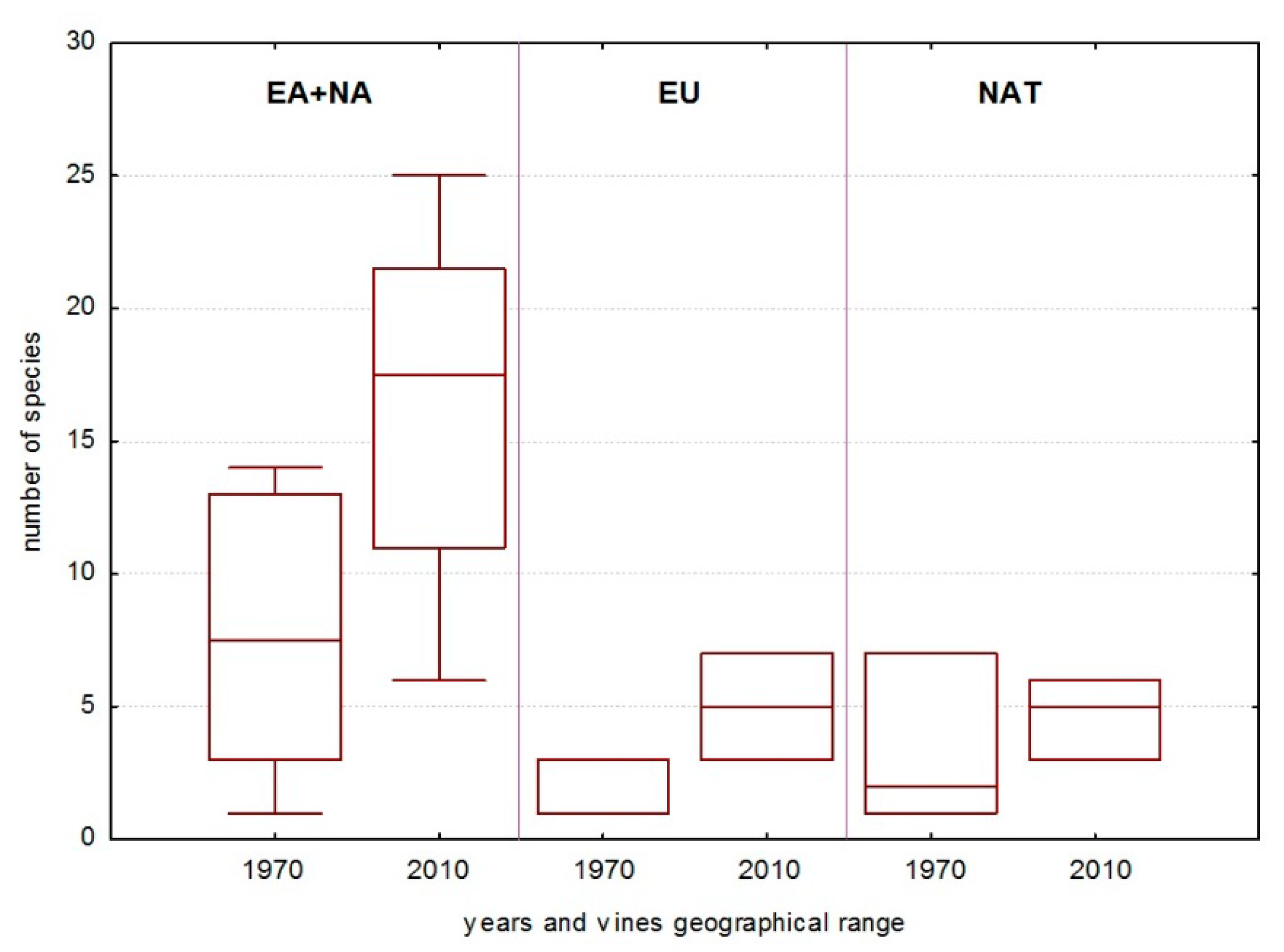
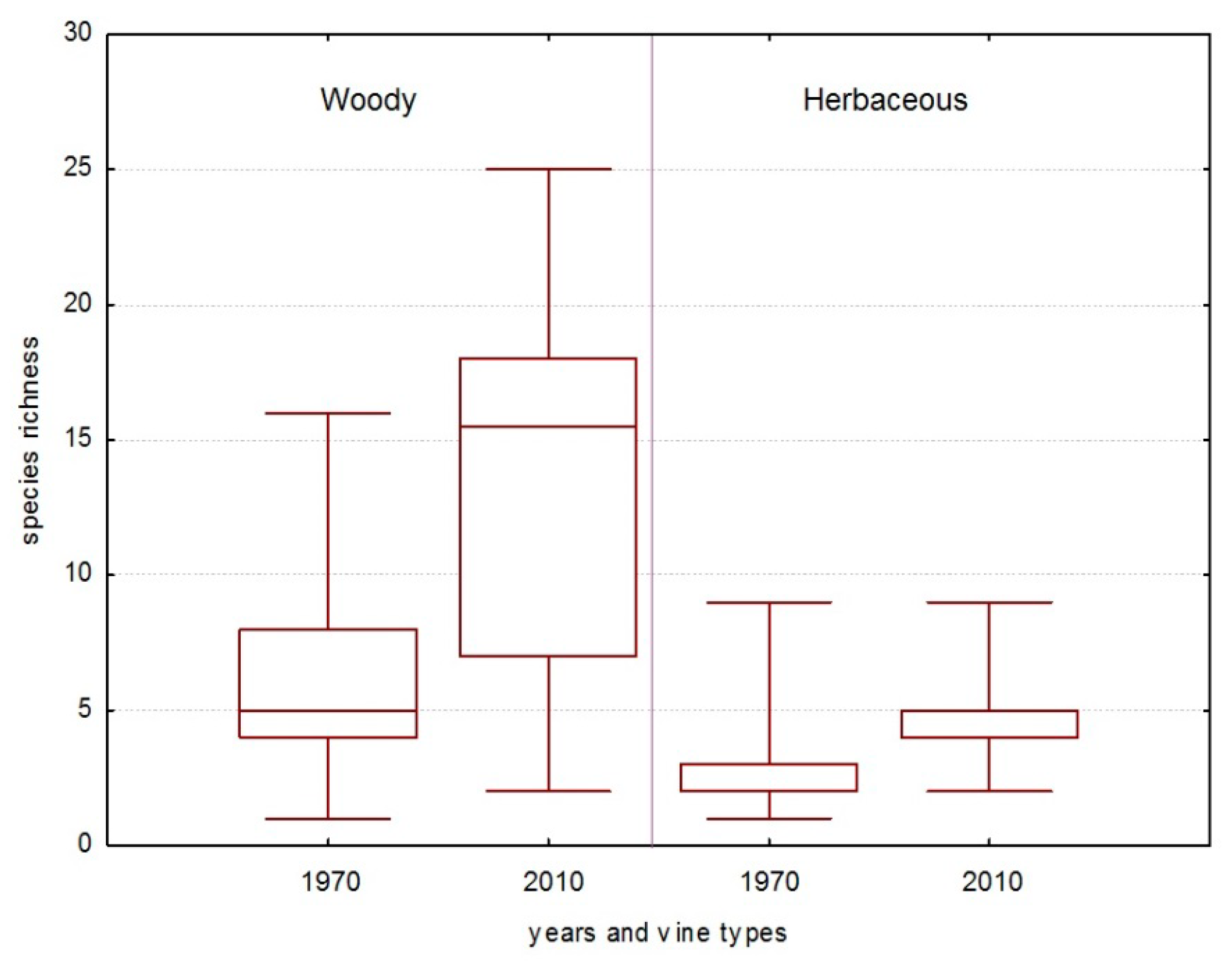
3.3. Fungal Groups’ Tendencies during the Last 40 Years
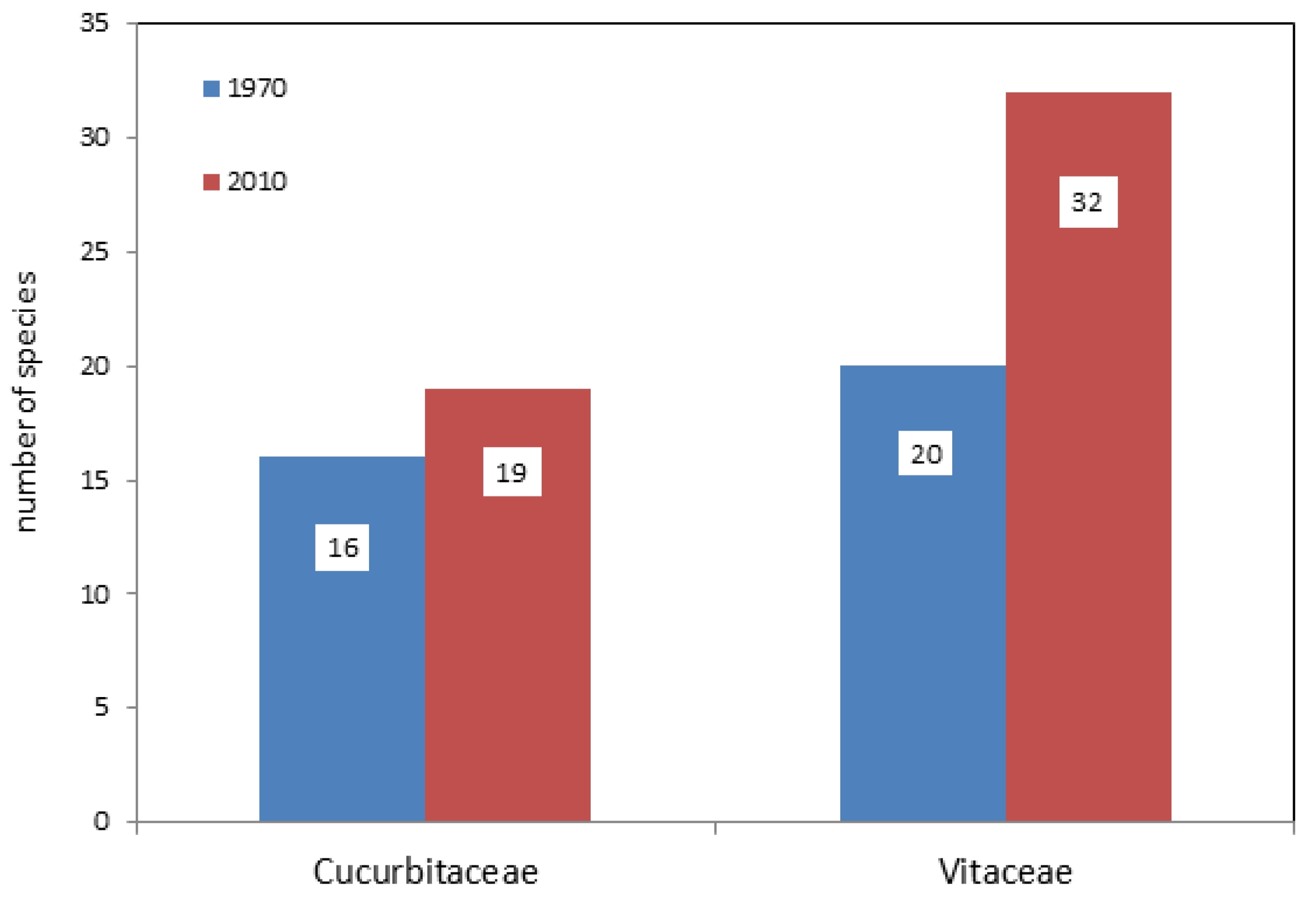
3.4. Phytopathogenic Fungi on Edible Vines
4. Discussion
4.1. General Results of 100-Year-Long Monitoring
4.2. Dynamics of the Model Groups during the Last 40 Years
4.3. Pathogenic Fungi on Alien Edible Vines
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dorofeyeva, L.M.; Mamaev, S.A. Introduction of the Far Eastern woody lianas in the Middle Ural and the prospects of their use. In Plants in a Monsoon Climate; Materials of the International Conference Dedicated to the 50th Anniversary of the Botanical Garden; Institute of the Far Eastern Branch of the Russian Academy of Sciences: Vladivostok, Russian, 1998; pp. 171–172. (In Russian) [Google Scholar]
- Mamaev, S.A. 70th Anniversary of the Botanical Garden of the Ural Branch, Russian Academy of Sciences; Ekaterinburg Publishing: Ekaterinburg, Russia, 2006. (In Russian) [Google Scholar]
- Vlasenko, V.E.; Dorofeyeva, L.M.; Yakovleva, S.V.; Semkina, L.A. Green plantations in the dendroparks of Ekaterinburg City. Bull. Samara Sci. Cent. RAS 2010, 10, 1376–1378. (In Russian) [Google Scholar]
- Dorofeyeva, L.M. Lianas collection from the Botanical Garden and its science-practice utilization. Northern Asia Plant Diversity 2021. BIO Web Conf. 2021, 38, 25. [Google Scholar] [CrossRef]
- Nemytov, A.Y. Shatilov’s Grape; Chelyabinsk Printing House: Chelyabinsk, Russia, 2016. (In Russian) [Google Scholar]
- Govorukhin, V.S. Flora of the Urals. Key Book of Plants Grow on the Urals and Hills from the Shores of KARA Sea till the Southern Treeline; Sredural Press: Sverdlovsk, Russia, 1937. (In Russian) [Google Scholar]
- Tretyakova, A.S.; Kulikov, P.V. Adventitious component of the flora of Sverdlovsk region: Dynamics of the species. Bull Udmurt. Univ. Biol. Earth Sci. 2013, 4, 184–188. [Google Scholar]
- Shiryaev, A.G.; Zmitrovich, I.V.; Shiryaeva, O.S. Species richness of Agaricomycetes on hedge vines in Ekaterinburg City (Russia). Mycol. Phytopathol. 2021, 55, 340–352. [Google Scholar] [CrossRef]
- Troshin, L.P. Native Grapes in Russia; Kuban University: Krasnodar, Russia, 2012. [Google Scholar]
- Bulgakov, T.S.; Shiryaev, A.G. New finds of phyllotrophic plant pathogenic microfungi in Ekaterinburg and its suburbs. Mycol. Phytopathol. 2021, 55, 405–410. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Clim. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- RIHMI-WDC. Federal Service for Hydrometeorology and Environmental Monitoring; RIHMI-WDC: Obninsk, Russia, 2021; Available online: https://www.meteo.ru (accessed on 17 August 2021).
- Leskinen, P.; Lindner, M.; Verkerk, P.J.; Nabuurs, G.-J.; Van Brusselen, J.; Kulikova, E.; Hassegawa, M.; Lerink, B. (Eds.) Russian Forests and Climate Change; What Science Can Tell Us 11; European Forest Institute: Joensuu, Finland, 2020. [Google Scholar] [CrossRef]
- Demidova, Z.A. Diseases overview of cultivated and wild plants in the Ural region. Bull. Uraloblzu. 1925, 10, 9–20. (In Russian) [Google Scholar]
- Stepanova, N.T.; Sirko, A.V. Flora of acomycetous and imperfect fungi of the Ural. Spore plants of the Ural. Proc. Inst. Plant Anim. Ecol. 1970, 70, 3–52. (In Russian) [Google Scholar]
- Blagoveshchenskaya, E.Y. Phytopathogenic Micromycetes: Educational Keybook; URSS Press: Moscow, Russia, 2015. (In Russian) [Google Scholar]
- Bondartsev, A.S.; Singer, R.A. A guide to collecting higher basidiomycetes for scientific study. Proc. Bot. Inst. AS USSR 1950, 2, 499–572. (In Russian) [Google Scholar]
- Index Fungorum, CABI, Kew. 2021. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 29 July 2021).
- Shiryaev, A.; Kotiranta, H.; Mukhin, V.A.; Stavishenko, I. Aphyllophoroid Fungi of Sverdlovsk Region, Russia. Biodiversity, Distribution, Ecology and the IUCN Threat Categories; Goshchitskiy Publisher: Ekaterinburg, Russia, 2010. [Google Scholar]
- Stepanova-Kartavenko, N.T. Aphyllophoroid Fungi of the Ural; UFAS USSR: Sverdlovsk, Russia, 1967. (In Russian) [Google Scholar]
- Stepanova, N.T. Ecogeographical Analysis of Aphyllophoroid Fungi of the Ural. Ph.D. Thesis, Institute UFAS USSR, Sverdlovsk, Russian, 1971. (In Russian). [Google Scholar]
- Shiryaeva, O.S. New records of Agaricoid fungi from Sverdlovsk region, Russia. Botanica 2018, 24, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Shiryaev, A.G.; Mukhin, V.A.; Kotiranta, H.; Stavishenko, I.V.; Arefyev, S.P.; Safonov, M.A.; Kodolapov, D.A. Biodiversity of aphyllophoroid fungi of the Ural. In Biological Diversity of the Urals’s Plant World and Adjacent Territories; Mukhin, V.A., Ed.; Institute of Plant and Animal Ecology: Ekaterinburg, Russia, 2012; pp. 311–313. (In Russian) [Google Scholar]
- Bondartseva, M.A. Keybook to Fungi of Russia. Aphyllophorales; Nauka: St. Pteresburg, Russia, 1998; Volume 2. (In Russian) [Google Scholar]
- Ryvarden, L.; Melo, I. Poroid Fungi of Europe; Fungiflora: Oslo, Norway, 2014. [Google Scholar]
- Bernicchia, A. A checklist of corticioid, polyporoid and clavarioid fungi (Basidiomycotina) from the Emilia-Romagna region, Italy. Sydowia 2001, 53, 1–33. [Google Scholar]
- Bernicchia, A.; Gorjon, S.P. Fungi Europaei: Corticiaceae s.l.; Edizioni Candusso: Origgio, Italy, 2010; Volume 12. [Google Scholar]
- Fischer, M.; Gonzalez, V. An annotated checklist of European basidiomycetes related to white rot of grapevine (Vitis vinifera). Phytopathol. Mediterr. 2015, 54, 281–298. [Google Scholar] [CrossRef]
- Karadelev, M.; Rusevska, K.; Kost, G.W.; Kopanja, D.M. Checklist of macrofungal species from the phylum Basidiomycota of the Republic of Macedonia. Acta Musei Maced. Sci. Nat. 1995, 21, 23–112. [Google Scholar]
- Isikov, V.P. Systematic Catalogue of Fungi on Woody Plants in Crimea; Arial Press: Simferopol, Russia, 2019. [Google Scholar]
- Bernicchia, A.; Marchisio, V.F.; Padovan, F.; Perini, C.; Ripa, C. Checklist of Italian Fungi. Basidiomycota; Onofri, S., Ed.; Commission on Fungi of the Organization for the Phyto-Taxonomic Investigation of the Mediterranean Area: Sassari, Italy, 2005. [Google Scholar]
- Tura, D.; Zmitrovich, I.V.; Wasser, S.P.; Nevo, E. Checklist of Hymenomycetes (Aphyllophorales s.l.) and Heterobasidiomycetes in Israel. Mycobiology 2010, 38, 256–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkina, I.S.; Mironova, L.P. Annotated list of basidiomycetous and ascomycetous macrofungi in the Karadag Nature Reserve. Sci. Note Nat. Res. Martyan Penins. 2015, 6, 297–327. [Google Scholar]
- Ablakatova, A.A. Mycoflora and Basic Fungal Disease of Fruit and Berry Crops at South of the Russian Far East; Nauka: Moscow, Russia, 1965. (In Russian) [Google Scholar]
- Azbukina, Z.M. (Ed.) Flora, Vegetation and Mycobiota of Ussury Nature Reserve; Dalnauka: Vladivostok, Russia, 2006. (In Russian) [Google Scholar]
- Egorova, L.N. (Ed.) Flora, Mycobiota and Vegetation of Lazovsky Nature Reserve; Russkiy Ostrov Publishing: Vladivostok, Russia, 2002. (In Russian) [Google Scholar]
- Pavlyuk, N.A. Plant’s Mycobiota of the Botanical Garden Far East Branch RAS. Ph.D. Thesis, Far East Branch RAS, Vladivostok, Russia, 2009. (In Russian). [Google Scholar]
- Golan, J.J.; Pringle, A. Long-distance dispersal of fungi. Microbiol. Spectr. 2017, 5, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingold, C.T. Dispersal in Fungi; Clarendon Press: Oxford, UK, 1953. [Google Scholar]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.; Liu, M.; Zhao, W.; Hyde, K.D.; Liu, J.; Yan, J. Biodiversity of fungi on Vitis vinifera L. revealed by traditional and high-resolution culture-independent approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef] [Green Version]
- Popushoy, I.S.; Marzhina, L.A. Fungal Diseases of Grapevine; Shtiintsa: Kishinev, Moldova, 1989. (In Russian) [Google Scholar]
- Khokhryakov, M.K.; Dobrozrakova, T.L.; Stepanov, K.M.; Letova, M.F. Guidebook of Plant Diseases, 3rd ed.; Lan Press: Saint-Petersburg, Russia, 2003. (In Russian) [Google Scholar]
- Gafforov, Y.S. A preliminary checklist of Ascomycetous microfungi from southern Uzbekistan. Mycosphere 2017, 8, 660–696. [Google Scholar] [CrossRef]
- Gaponenko, N.I. Survey of the Fungi of Bukhara; Akademy of Science of Uzbek SSR Publ.: Tashkent, Uzbekistan, 1965. (In Russian) [Google Scholar]
- Burdinskaya, V.F.; Poymanov, V.E.; Tolokova, R.P. Diseases and Pests of Grapevine and Their Control; Potapenko, Y.I., Ed.; VNIIViV Press: Novocherkassk, Russia, 2009. (In Russian) [Google Scholar]
- Collado, E.; Bonet, J.A.; Camarero, J.J.; Egli, S.; Peter, M.; Salo, K.; Martínez-Peña, F.; Ohenoja, E.; Martín-Pinto, P.; Primicia, I.; et al. Mushroom productivity trends in relation to tree growth and climate across different European forest biomes. Sci. Total Environ. 2019, 689, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Nnadi, N.E.; Carter, D.A. Climate change and the emergence of fungal pathogens. PLoS Pathog. 2021, 17, e1009503. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cucurbitaceae | Vitaceae |
|---|---|
| Macrofungi | |
| Ceratobasidium cornigerum | Armillaria borealis |
| Rhizoctonia solani | Athelia rolfsii |
| Typhula micans | Ceratobasidium cornigerum |
| Typhula culmigena | Fomitiporia punctata |
| Ganoderma applanatum | |
| Irpex lacteus | |
| Inonotus hispidus | |
| Phanerochaete velutina | |
| Phellinopsis conchata | |
| Pholiota limonella | |
| Pleurotus pulmonarius | |
| Schizophyllum commune | |
| Stereum hirsutum | |
| Microfungi | |
| Alternaria alternata | Alternaria alternata |
| Alternaria cucumerina | Aureobasidium pullulans |
| Botrytis cinerea | Botrytis cinerea |
| Cladosporium cucumerinum | Colletotrichum parthenocissicola |
| Colletotrichum orbiculare | Colletotrichum quinquefoliae |
| Fusarium oxysporum | Coniella diplodiella |
| Golovinomyces tabaci | Diplodia seriata |
| Podosphaera xanthi | Elsinoe ampelina |
| Pseudoperonospora cubensis | Erysiphe necator |
| Pythium aphanidermatum | Eutypa scabrosa |
| Septoria cucurbitacearum | Fusarium oxysporum |
| Sclerotinia sclerotiorum | Globisporangium debaryanum |
| Stagonosporopsis cucurbitacearum | Nectria cinnabarina |
| Verticillium alboatrum | Phaeomoniella chlamydospora |
| Verticillium dahliae | Phyllosticta ampelicida |
| Plasmopara viticola | |
| Pseudocercospora vitis | |
| Ramularia vitis | |
| Verticillium glaucum | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiryaev, A.G.; Zmitrovich, I.V.; Bulgakov, T.S.; Shiryaeva, O.S.; Dorofeyeva, L.M. Global Warming Favors the Development of a Rich and Heterogeneous Mycobiota on Alien Vines in a Boreal City under Continental Climate. Forests 2022, 13, 323. https://doi.org/10.3390/f13020323
Shiryaev AG, Zmitrovich IV, Bulgakov TS, Shiryaeva OS, Dorofeyeva LM. Global Warming Favors the Development of a Rich and Heterogeneous Mycobiota on Alien Vines in a Boreal City under Continental Climate. Forests. 2022; 13(2):323. https://doi.org/10.3390/f13020323
Chicago/Turabian StyleShiryaev, Anton G., Ivan V. Zmitrovich, Timur S. Bulgakov, Olga S. Shiryaeva, and Lyudmila M. Dorofeyeva. 2022. "Global Warming Favors the Development of a Rich and Heterogeneous Mycobiota on Alien Vines in a Boreal City under Continental Climate" Forests 13, no. 2: 323. https://doi.org/10.3390/f13020323