
Inter- and Intraxylary Phloem in Vascular Plants: A Review of Subtypes, Occurrences, and Development
 ,
,
Abstract
:1. Introduction
2. List of Species Investigated and Sample Processing
3. Inter- and Intraxylary Phloem in Vascular Plants
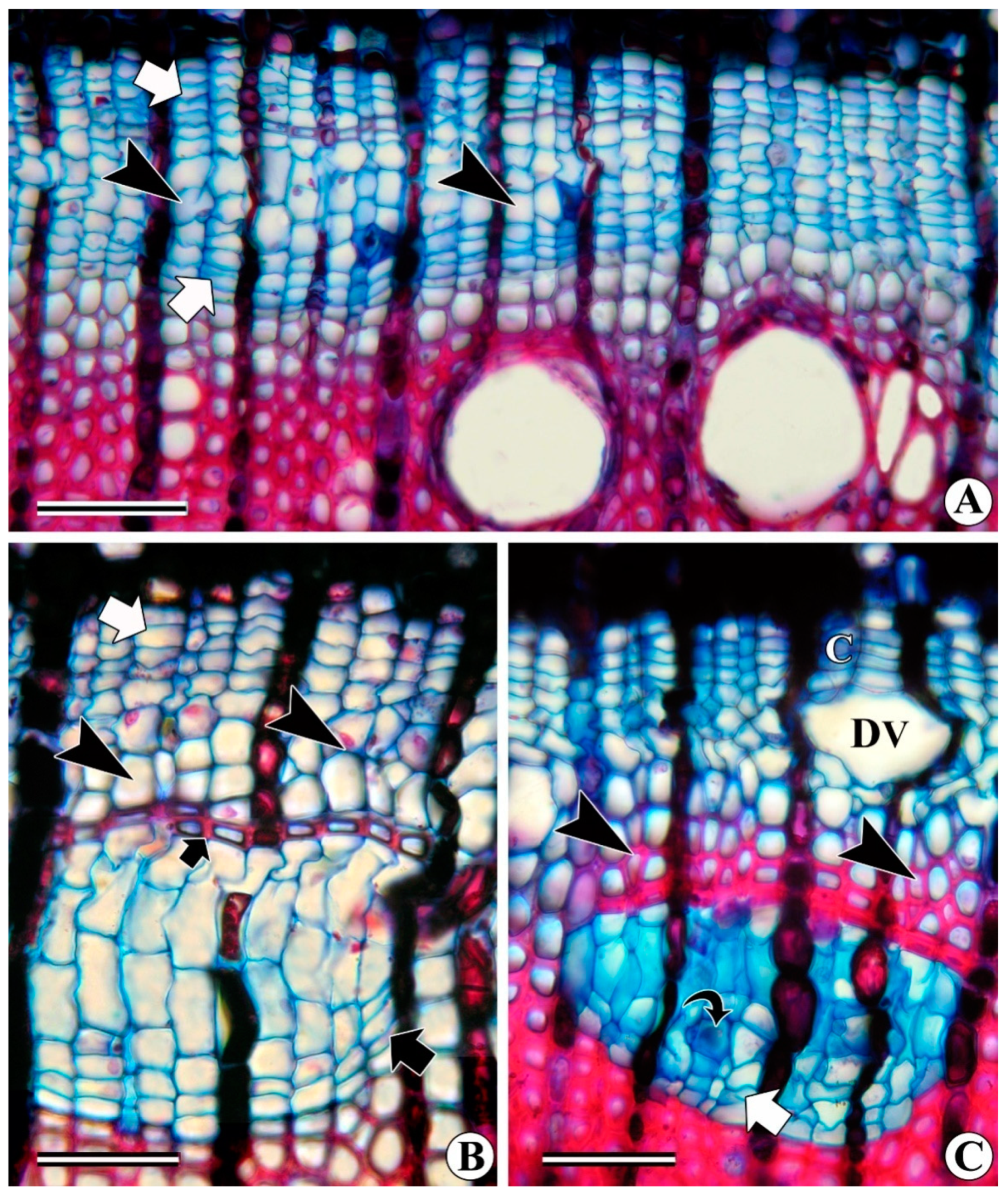
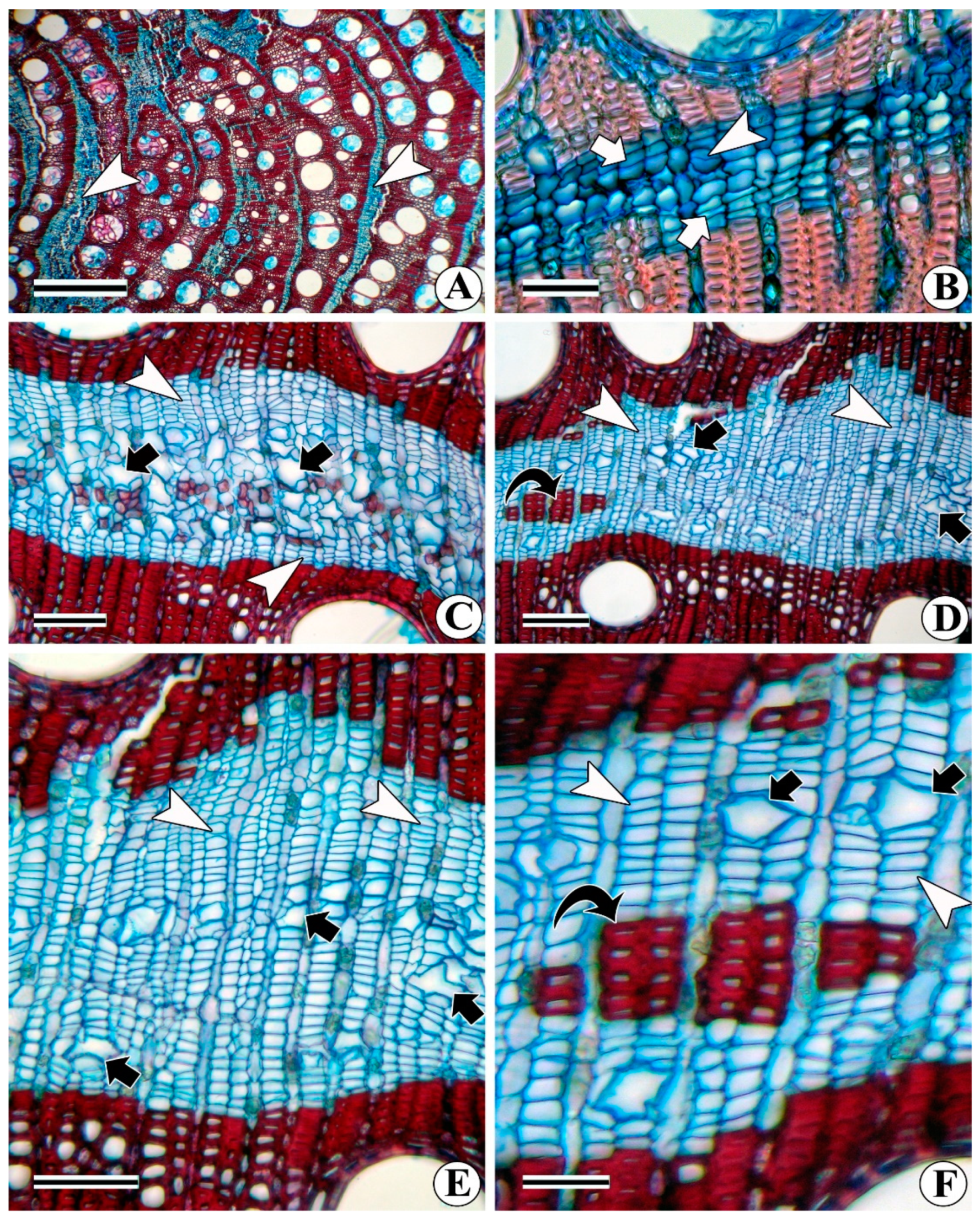
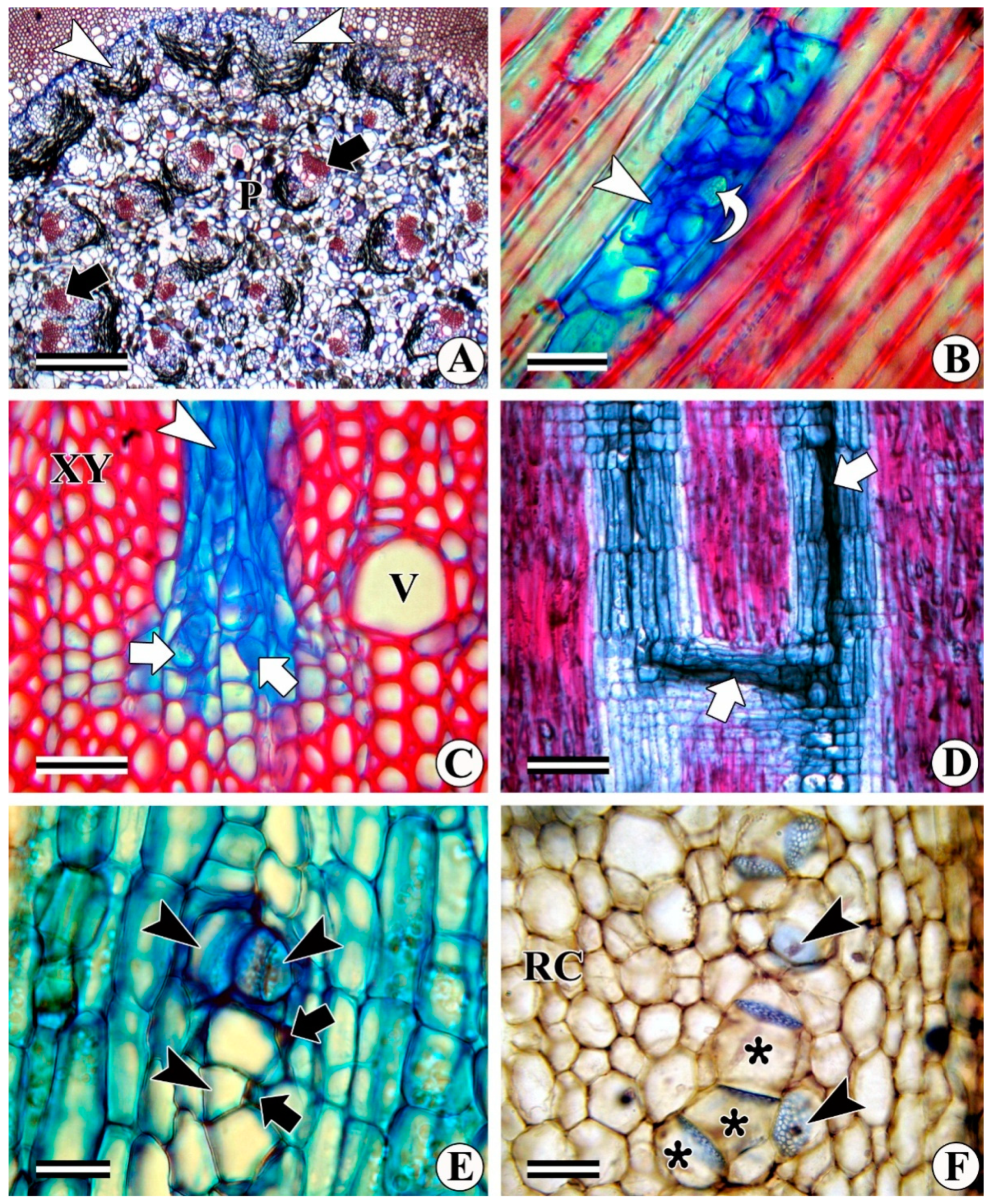
3.1. Interxylary (Included) Phloem
3.1.1. Combretum Subtype
3.1.2. Strychnos Subtype
3.1.3. Azima Subtype
3.1.4. Calycopteris Subtype
3.2. Intraxylary (Internal/Medullary) Phloem
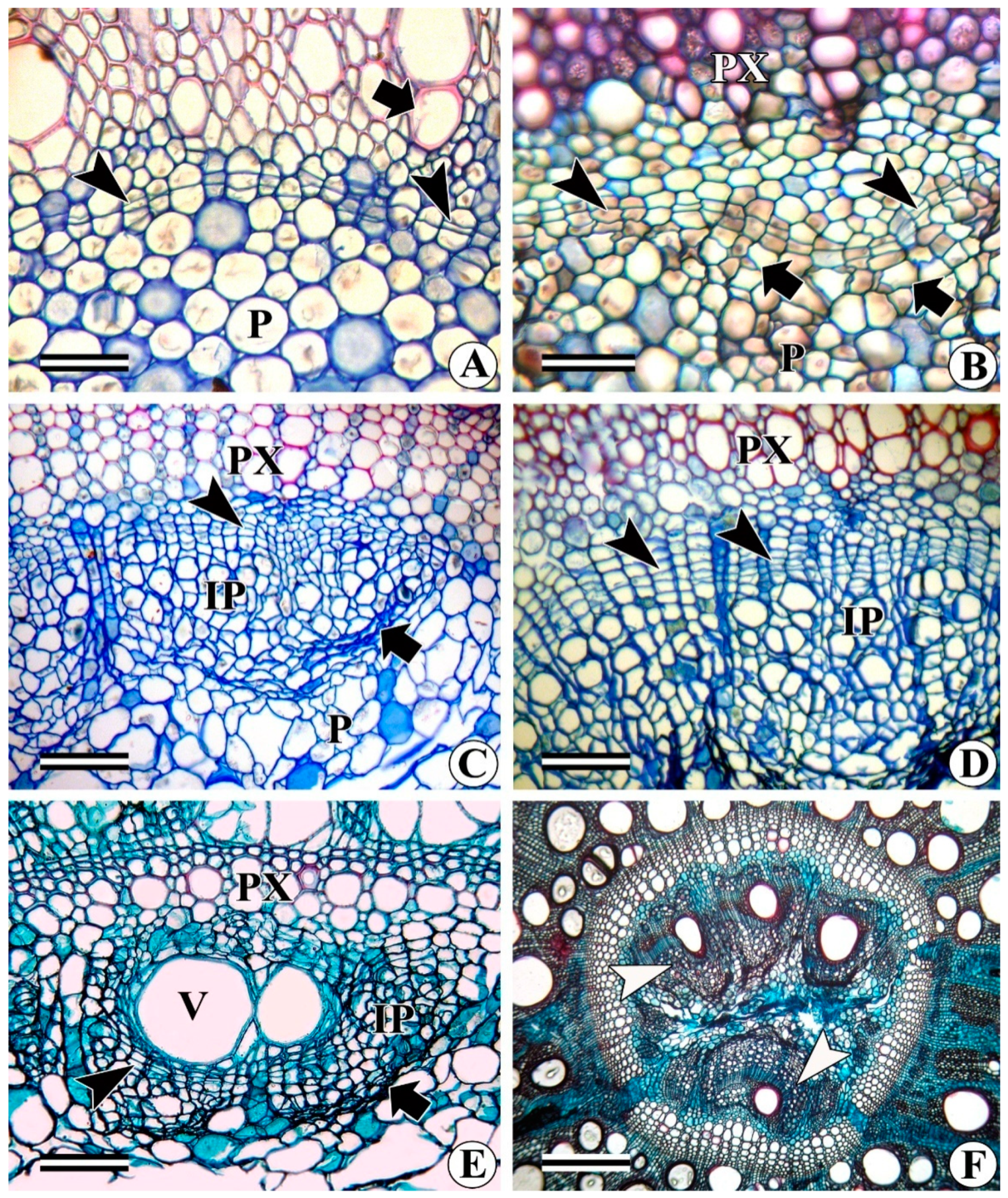
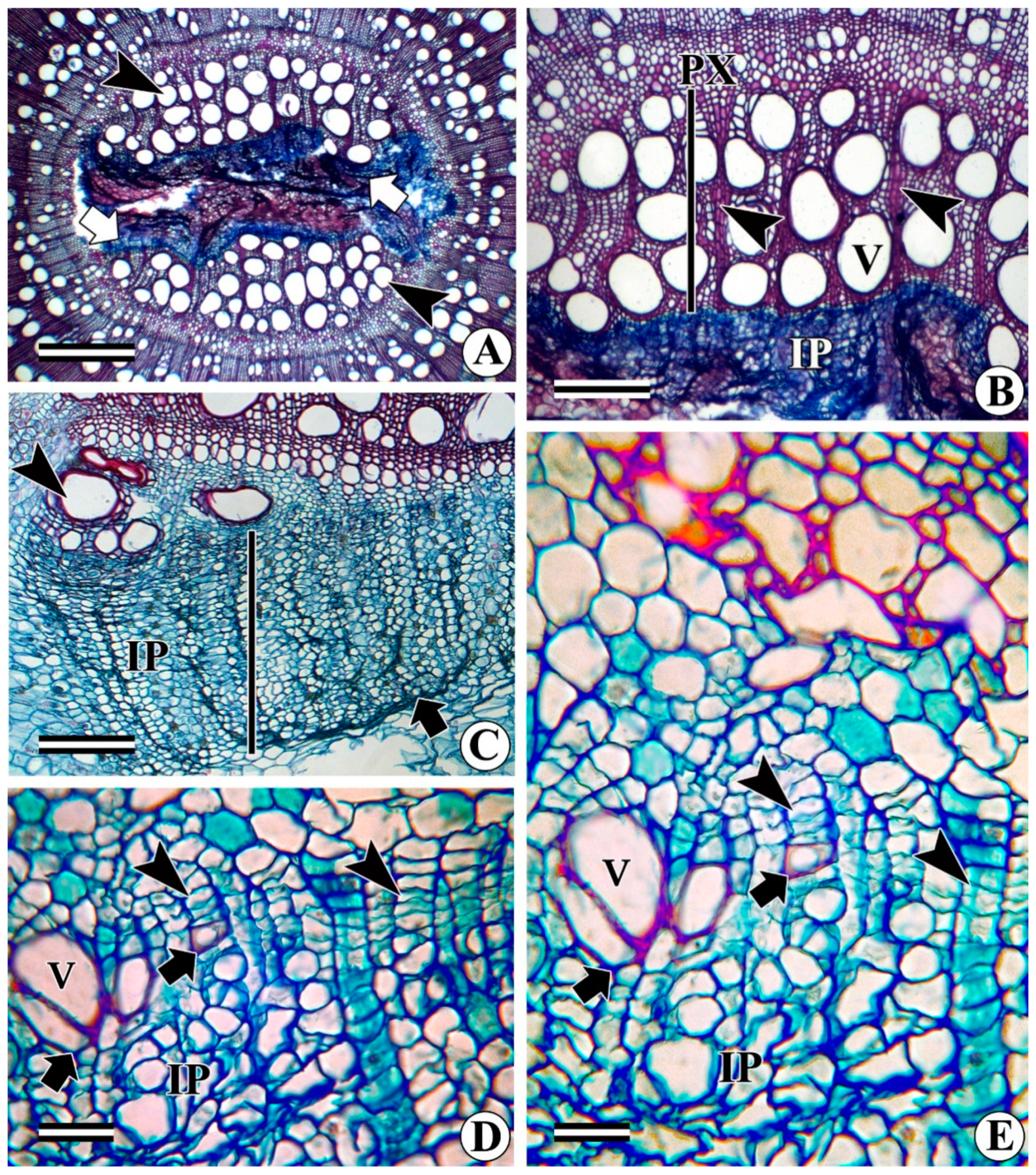
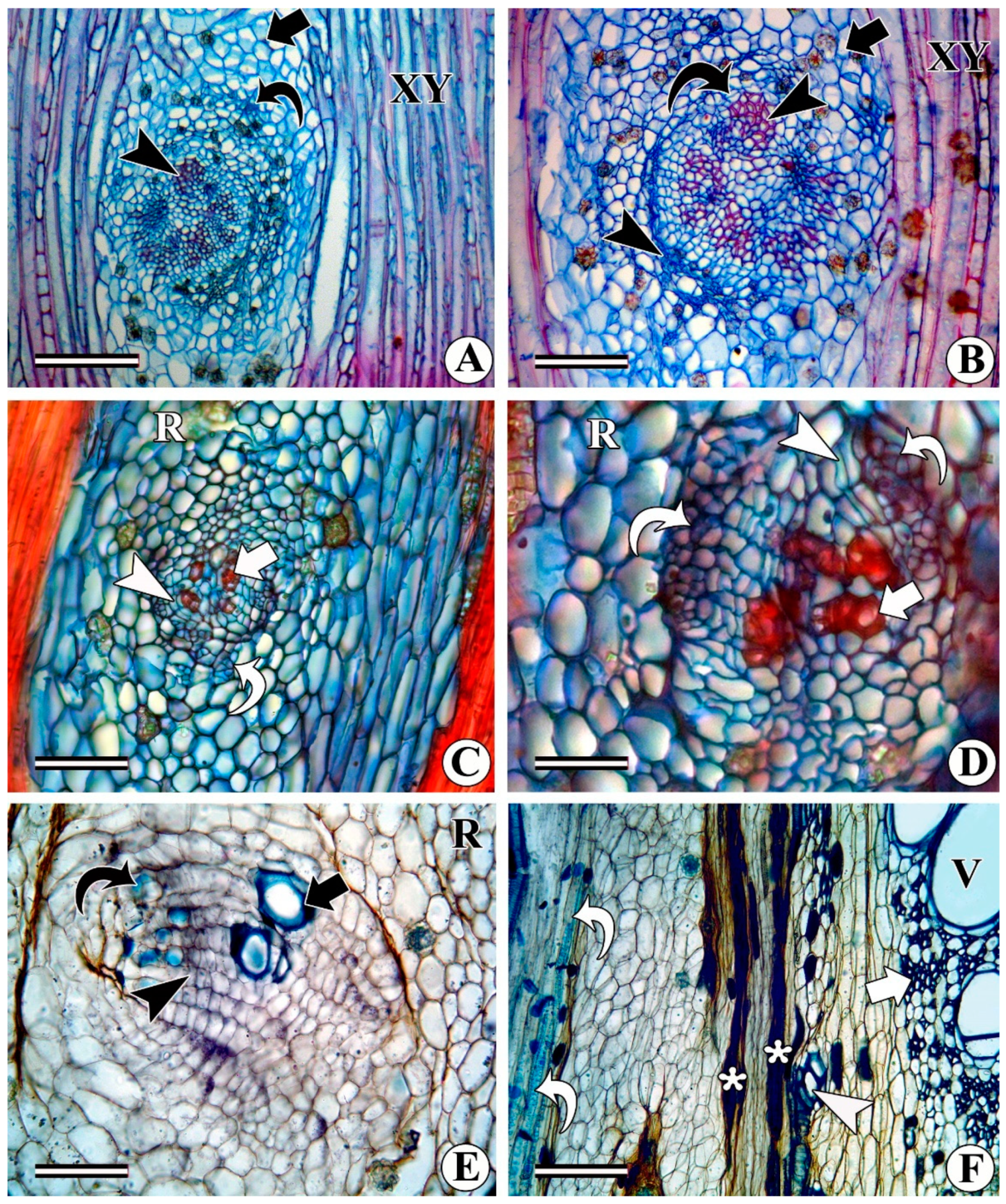
- Development of intraxylary phloem cambium (medullary cambium): In relatively thick stems (as per our phenological observations when the plant enters the reproductive phase), the marginal pith cells become meristematic and divide periclinally to form radial files of cells (Figure 10A–D) that share all the characteristics of the vascular cambium. This is variously referred to as internal cambium, intraxylary phloem cambium, and medullary cambium/peri-medullary cambium. the cell division activity of the intraxylary phloem cambium differs from species to species and sometimes it becomes species-specific. In most species, it deposits only phloem derivatives (e.g., Argyreia elliptica, A. nervosa, Dicranostylis ampla, Solanum pseudocapsicum) even in fully grown thick stems (Figure 10C,D). In contrast, in species such as Campsis radicans, Hewettia malabarica, I. hederifolia, I. muricata, Turbina corymbosa, Vallaris solanacea, and several other members of the Convolvulaceae family, this cambium is functionally bidirectional and produces both secondary xylem centrifugally and secondary phloem centripetally (Figure 10C–F, 11A–C). Depending on the species, the ratio of the xylem to phloem production from the intraxylary phloem cambium differs from species to species. Some species such as Campsis radicans (Figure 11A,B) and Hewettia malabarica (Figure 11C) show the deposition of more xylem than phloem. Products of this cambium consist of wide and narrow (fibriform) vessels, fibres, and rays in the xylem (Figure 11B), whereas the phloem is composed of sieve elements, companion cells, and associated axial and ray parenchyma cells (Figure 10C). Some samples of Ipomoea turbinata show a unique behaviour of the intraxylary phloem cambium, where it is functionally unidirectional and produces both secondary xylem and phloem centripetally, i.e., in the same direction (Figure 11D,E). Some of the samples of the same species (i.e., I. turbinata) also show the presence of two intraxylary cambia on either side of the lignified tissue, in which the outer (adjacent to protoxylem) cambium deposits secondary phloem externally and xylem internally, whereas the inner cambium produces phloem centripetally and xylem centrifugally (Figure 12A). Structurally, the phloem remains similar to the regular external phloem and can possess simple or compound sieve plates that are oriented transversely or obliquely to the main axis (Figure 12B).
3.3. Formation of Ray (Radial) Sieve Elements and Vascular Bundles in Rays
3.4. Formation of Phloem Wedges
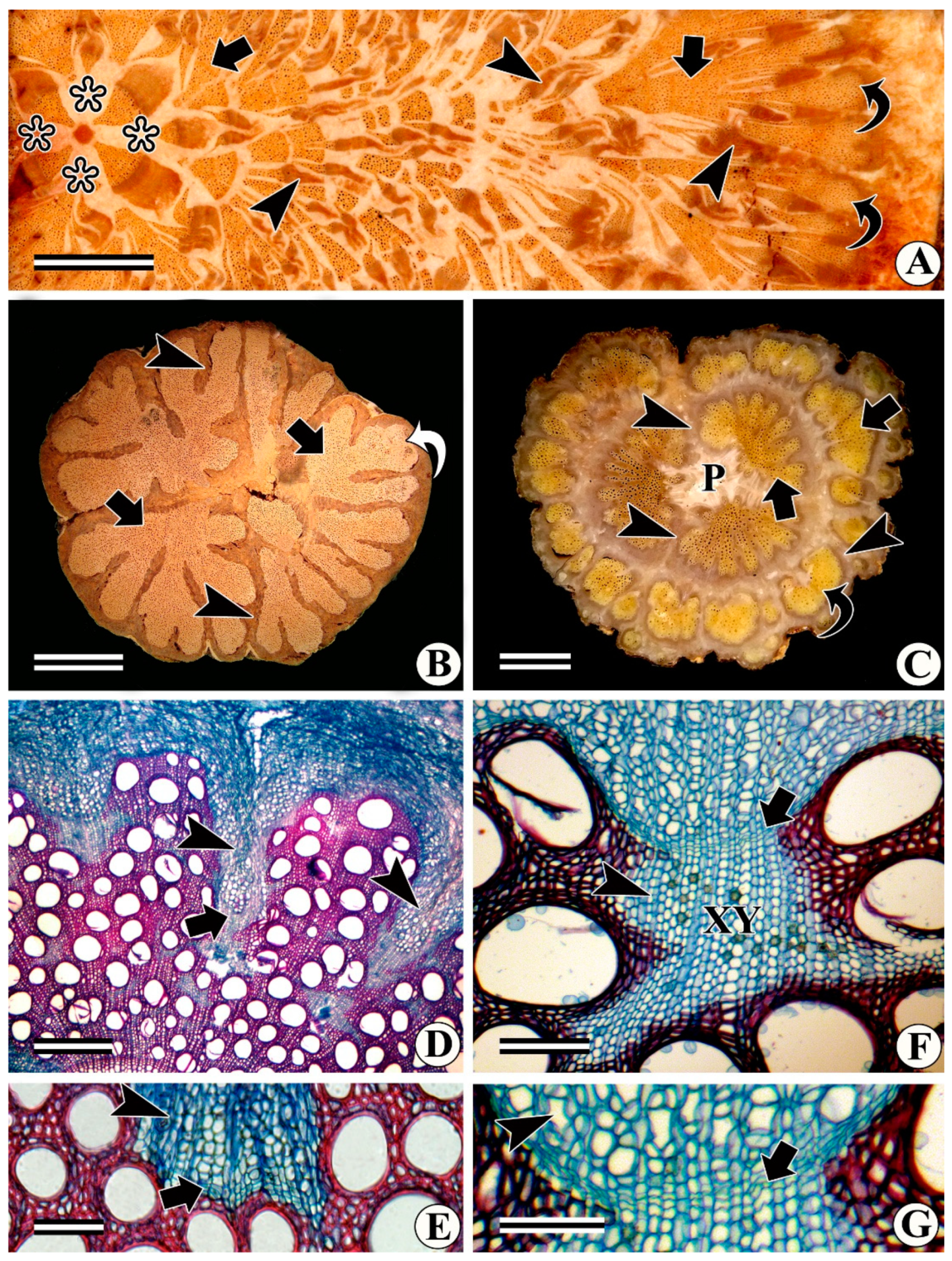
4. Interxylary Phloem in Some Families of Caryophyllales
Fate of Previously Formed Phloem Islands
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez-Villalon, A. Wiring a plant: Genetic networks for phloem formation in Arabidopsis thaliana roots. New Phytol. 2016, 210, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Evert, R.F. Esau’s plant anatomy. In Meristems, Cells and Tissues of Plant Body: Their Structure, Function and Development, 3rd ed.; John Wiley and Son Inc: Hoboken, NJ, USA, 2006; p. 567. [Google Scholar]
- Taylor, E.L. Phloem evolution: An appraisal based on fossil record. In Sieve Elements Comparative Structure, Induction, and Development; Behnke, H.-D., Sjolund, R.D., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany; New York, NY, USA, 1990; pp. 285–294. [Google Scholar]
- Scott, D.H.; Brebner, G. On the anatomy and histology of Strychnos. Ann. Bot. 1889, 3, 275–304. [Google Scholar] [CrossRef]
- Scott, D.H.; Brebner, G. On internal phloem in the root and stem of dicotyledons. Ann. Bot. 1891, 5, 259–300. [Google Scholar] [CrossRef] [Green Version]
- Hallier, H.J.G. Versuch Einer Naturlichen Gliederung der Convolvulaceen auf morphologischer und anatomischergr undlage. Bot. Jahrb. Für Syst. Pflanzen. Pflanzengeogr. 1893, 16, 453–591. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons; Clarendon Press: Oxford, UK, 1950. [Google Scholar]
- Fukuda, Y. Anatomical study of the internal phloem in the stems of dicotyledons, with special reference to its histogenesis. J. Fac. Sci. Univ. Tokyo Sect III 1967, 9, 313–375. [Google Scholar]
- Carlquist, S. Comparative Wood Anatomy: Systematic, Ecological, and Evolutionary Aspect of Dicotyledon Wood; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2001; p. 411. [Google Scholar]
- Carlquist, S.; Hanson, M.A. Wood and stem anatomy of Convolvulaceae: A survey. Aliso 1991, 13, 51–94. [Google Scholar] [CrossRef] [Green Version]
- Evert, R.F. The cambium and seasonal development of the phloem in Pyrus malus. Am. J. Bot. 1963, 50, 149–159. [Google Scholar] [CrossRef]
- Evert, R.F. Seasonal development of the secondary phloem in Rhus glabra L. Bot. Soc. Am. Misc. Series Publ. 1978, 156, 25. [Google Scholar]
- Liphschitz, N.; Waisel, Y. Effects of environment on relations between extension and cambial growth of Populuseuphratica Oliv. New Phytol. 1970, 69, 1059–1064. [Google Scholar] [CrossRef]
- Ghouse, A.K.M.; Hashmi, S. Cambium periodicity in Polyalthialongifolia. Phytomorphology 1979, 29, 64–67. [Google Scholar]
- Savidge, R.A. Seasonal cambial activity in Larix laricina saplings in relation to endogenous Indol-3-Acetic Acid, Sucrose, and Coniferin. Forest Sci. 1991, 37, 953–958. [Google Scholar]
- Rajput, K.S.; Rao, K.S. Seasonal anatomy of secondary phloem of teak (Tectona grandis L. Verbenaceae) growing in dry and moist deciduous forests. Phyton 1998, 38, 251–258. [Google Scholar]
- Esau, K. Development and structure of the phloem tissue. Bot. Rev. 1939, 7, 373–432. [Google Scholar] [CrossRef]
- Hartig, T. Vergleichende Untersuchungeniuber die Organi- sation des Stammes der einheimischen Waldbaume. Jahresber. Fortschr. Forstwissensch. Forstl. Naturkunde 1837, 1, 125–168. [Google Scholar]
- Von Mohl, H. Einige Andeutungeniuber den Bau des Bastes. Bot. Zeitung 1855, 13, 873–881; 889–897. [Google Scholar]
- Nägeli, C.W. Das Wachsthum des Stammes und der Wurzelbei den Gefisspflanzen und die Anordnung der Gefassstrangeim Stengel. Beitr. Zur Wiss. Bot. Heft 1858, 1, 1–156. [Google Scholar]
- Wilhelm, K. Beitragezur Kenntnis des Siebrhrenapparates dicotyler; Pflanzen: Berlin, Germany, 1880; p. 90. [Google Scholar]
- De Bary, A. Comparative Anatomy of the Vegetative Organs of the Phanerogams and Ferns; Bower, F.O.; Scott, D.H., Translators; The Clarendon Press: Oxford, UK, 1884; pp. 578–580. [Google Scholar]
- Lecomte, H. Contribution a l’etude du liber des Angiospermes. Ann. Sci. Nat. Bot. 1889, 10, 193–324. [Google Scholar]
- De Janczewski, E. Etudes compareessur les tubes cribreux. MWm. Soc. Nat. Sci. Nat. Math. Cherbg. 1881, 23, 209–350. [Google Scholar]
- Russow, E. Ueber den Bau und die Entwicklung der Siebr6hren und Bau und Entwicklung der secundaren Rinde der Dicotylen und Gymnospermen. Sitzber. Naturf. Ges. Univ. Dorpat 1882, 6, 257–327. [Google Scholar]
- Artschwager, E.F. Studies on the potato tuber. J. Agri. Res. 1924, 27, 809–835. [Google Scholar]
- Crafts, A.S. Phloem anatomy, exudation, and transport of organic nutrients in cucurbits. Plant Physiol. 1932, 7, 183–225. [Google Scholar] [CrossRef] [PubMed]
- Esau, K. Ontogeny of phloem in the sugar beet (Beta vulgaris L.). Amer. J. Bot. 1934, 21, 632–644. [Google Scholar] [CrossRef]
- Esau, K. Initial localization, and subsequent spread of curly-top symptoms in the sugar beet. Hilgardia 1935, 9, 397–436. [Google Scholar] [CrossRef]
- Esau, K. Ontogeny and structure of the phloem of tobacco. Hilgardia 1938, 11, 343–424. [Google Scholar] [CrossRef] [Green Version]
- Abbe, L.B.; Crafts, A.S. The phloem of white pine and other coniferous species. Bot. Gaz. 1939, 100, 659–722. [Google Scholar] [CrossRef]
- Oparka, K.J.; Cruz, S.S. The great escape: Phloem transport and unloading of macromolecules. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 323–347. [Google Scholar] [CrossRef] [Green Version]
- Omid, A.; Keilin, T.; Glass, A.; Leshkowitz, D.; Wolf, S. Characterization of phloem-sap transcription profile in melon plants. J. Expt. Bot. 2007, 58, 3645–3656. [Google Scholar] [CrossRef] [Green Version]
- Haberland, G. Physiological Wood Anatomy, 4th ed.; Montagu Drummond, B.A., Translator; MacMillon and Company Ltd.: New York, NY, USA; London, UK, 1914. [Google Scholar]
- Drobnitch, S.T.; Jensen, K.H.; Prentice, P.; Pittermann, J. Convergent evolution of vascular optimization in kelp (Laminariales). Proc. Royal Soc. B. 2015, 282, 20151667. [Google Scholar] [CrossRef] [Green Version]
- Pace, M.R. Phloem: Cell Types, Structure, and Commercial Uses. In Plant Science-Structure, Anatomy and Physiology in Plants Cultured In Vivo and In Vitro; Gonzalez, A., Rodriguez, M., Sağlam, N.G., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.A. Vascular transport of auxins and cytokinin in Ricinus. Plant Growth Regul. 2000, 32, 157–160. [Google Scholar] [CrossRef]
- Kehr, J. Phloem sap proteins: Their identities and potential roles in the interaction between plants and phloem-feeding insects. J. Expt. Bot. 2006, 57, 767–774. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, R.; Wolf, S. Phloem transport: Cellular pathways and molecular trafficking. Ann. Rev. Plant Biol. 2009, 60, 207–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Tolstikov, V.; Turnbull, C.; Hicks, L.M.; Fiehn, O. Divergent metabolome and proteome suggest functional independence of dual phloem transport systems in cucurbits. Proc. Natl. Acad. Sci. USA 2010, 107, 13532–13537. [Google Scholar] [CrossRef] [PubMed]
- Azizpor, P.; Sullivan, L.; Lim, A.; Groover, A. Facile labeling of sieve element phloem protein bodies using the reciprocal oligosaccharide probe OGA488. Front. Plant Sci. 2022, 13, 809923. [Google Scholar] [CrossRef]
- Bonnemain, J.L. Transport du 14C assimile chez les Solanacees. Rev. Gén. Bot. 1968, 75, 579–610. [Google Scholar]
- Bonnemain, J.L. Transport chez la plantesuperieure. Physiol. Veg. 1972, 10, 697–729. [Google Scholar]
- Bonnemain, J.L. Microautoradiography as a tool for the recognition of phloem transport. Ber. Deut. Bot. Ges. Bd. 1980, 93, 99–107. [Google Scholar]
- Zamski, E.; Tsivion, Y. Translocation in plants possessing supernumerary phloem. I. 14C assimilates and auxin in the internal phloem of tobacco (Nicotiana tabacum L). J. Expt. Bot. 1977, 28, 117–126. [Google Scholar] [CrossRef]
- Fredon, J.J.; Bonnemain, J.L. Transport du 14C assimile chez le radis a divers stades de son developpement. C. r. hebd. Seanc. Acad. Sci. Paris D 1970, 270, 354–357. [Google Scholar]
- Turgeon, R.; Oparka, K. The secret phloem of pumpkins. Proc. Natl. Acad. Sci. USA 2010, 107, 13201–13202. [Google Scholar] [CrossRef] [Green Version]
- Rajput, K.S. Formation of unusual tissue complex in the stem of Adansonia digitata Linn. (Bombacaceae). Beitr. Biol. Pflanzen. 2004, 73, 331–342. [Google Scholar]
- Kapadane, K.K.; Lawand, P.R.; Shimpale, V.B.; Rajput, K.S. Development of the cambial variant and structure of the secondary xylem in Argyreia elliptica (Roth.) Choisy (Convolvulaceae). J. Plant Growth Regul. 2022; Published online. [Google Scholar] [CrossRef]
- Kapadane, K.K.; Shelke, A.R.; Gondaliya, A.D.; Rajput, K.S. Formation of medullary phloem in Argyreia nervosa (Burm. f.) Bojer. Plant Sci. Today 2019, 6, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Kapadane, K.K. Stem anatomy of some species of Argyreia (Convolvulaceae). Ph. D. Thesis, North Maharashtra University, Jalgaon, India, 2021. [Google Scholar]
- Gondaliya, A.D.; Lekhak, M.M.; Yadav, S.R.; Rajput, K.S. Structure of secondary xylem and development of intraxylary phloem in Beaumontia jerdoniana (Apocynaceae). Phyton 2018, 57. [Google Scholar]
- Rajput, K.S.; Rao, K.S. Cambial anatomy and absence of rays in the stem of Boerhaavia species (Nyctaginaceae). Ann. Bot. Fennici. 1998, 35, 131–135. [Google Scholar]
- Rajput, K.S.; Rao, K.S. Occurrence of sieve elements in phloem rays. IAWA J. 1997, 18, 197–201. [Google Scholar] [CrossRef]
- Rajput, K.S.; Patil, V.S.; Rao, K.S. Development of included phloem of Calycopteris floribunda Lamk. (Combretaceae). J. Torrey Bot. Soc. 2009, 136, 302–312. [Google Scholar] [CrossRef]
- Rajput, K.S.; Gondaliya, A.D.; Lekhak, M.M.; Yadav, S.R. Structure and ontogeny of intraxylary secondary xylem and phloem development by the internal vascular cambium in Campsis radicans (L.) Seem. (Bignoniaceae). J. Plant Growth Regul. 2018, 37, 755–767. [Google Scholar] [CrossRef]
- Rajput, K.S. Structure of cambium and its derivatives in the compressed stem of Canavalia ensiformis (L.) DC. (Fabaceae). Phyton 2003, 43, 135–146. [Google Scholar]
- Patil, V.S.; Marcati, C.R.; Rajput, K.S. Development of intra-and interxylary phloem in Coccinia indica (Cucurbitaceae). IAWA J. 2011, 32, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Moya, R.; Gondaliya, A.D.; Rajput, K.S. Formation of successive cambia and structure of secondary xylem in Dicranostyles ampla and Maripa nicaraguensis (Convolvulaceae). Phyton 2019, 119–129. [Google Scholar]
- Tamaio, N.; Braga, J.M.A.; Rajput, K.S. Development of successive cambia and structure of secondary xylem in the stems and roots of Distimake tuberosus (Convolvulaceae). Flora 2021, 279, 151814. [Google Scholar] [CrossRef]
- Rajput, K.S.; Romeiro, D.; Longui, E.L.; Marcati, C.R. Development of successive cambia and structure of wood in Gallesia integrifolia (Spreng.) Harms (Phytolaccaceae). Trees 2012, 26, 1943–1950. [Google Scholar] [CrossRef]
- Rajput, K.S.; Gondaliya, A.D.; Baijnath, H. Development of cambial variant and parenchyma proliferation in Hewittia malabarica (Convolvulaceae) from India and South Africa. IAWA J. 2021, 42, 50–63. [Google Scholar] [CrossRef]
- Rajput, K.S.; Raole, V.M.; Gandhi, D. Radial secondary growth, formation of successive cambia and their products in Ipomoea hederifolia L. (Convolvulaceae). Bot. J. Linn. Soc. 2008, 158, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Rajput, K.S.; Patil, V.S.; Rao, K.S. Multiple cambia and secondary xylem of Ipomoea pes-caprae (L.) R. Br. (Convolvulaceae). Acta Bot. Gall. 2014, 161, 13–19. [Google Scholar] [CrossRef]
- Rajput, K.S. Development of inverse cambia and structure of secondary xylem in Ipomoea turbinata (Convolvulaceae). Pol. Bot. J. 2017, 62, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Patil, V.S.; Rajput, K.S. Structure and development of inter- and intraxylary phloem in Leptadenia reticulata (Asclepiadaceae). Pol. Bot. J. 2008, 53, 5–15. [Google Scholar]
- Gondaliya, A.D.; Rajput, K.S. Stem Anatomy and development of intraxylary phloem in Vallaris solanacea (Roth) Kuntze (Apocynaceae). J. Indian bot. Soc. 2016, 95, 16–27. [Google Scholar]
- Gondaliya, A.D.; Rajput, K.S. Stem anatomy and development of inter- and intraxylary phloem in Leptadenia pyrotechnica (Forssk.) Decne. (Asclepiadaceae). Plant Biosystem. 2017, 151, 855–865. [Google Scholar] [CrossRef]
- Rajput, K.S.; Rao, K.S. Structural and developmental studies on cambial variant in Pupalia lappacea (Amaranthaceae). Ann. Bot. Fennici. 1999, 36, 137–141. [Google Scholar]
- Patil, V.S.; Koyani, R.D.; Sanghvi, G.V.; Rajput, K.S. Structure and development of internal phloem in Solanum pseudocapsicum (Solanaceae). IAWA J. 2014, 35, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rajput, K.S.; Gondaliya, A.D.; Moya, R. Structure of the secondary xylem and development of a cambial variant in Serjania mexicana (Sapindaceae). IAWA J. 2022, 43, 103–115. [Google Scholar] [CrossRef]
- Rajput, K.S.; Fiamengui, M.B.; Marcati, C.R. Stem anatomy of Strychnos bicolor Prog. (Loganiaceae) from Brazilian Cerrado. Phytomorphology 2010, 60, 49–57. [Google Scholar]
- Moya, R.; Gondaliya, A.D.; Rajput, K.S. Stem anatomy and development of interxylary phloem in Strychnos bredemeyeri (Loganiaceae). Ann. Biol. 2017, 39, 75–87. [Google Scholar] [CrossRef]
- Rajput, K.S. Occurrence of radial sieve elements in the phloem rays of some tropical species. Israel J. Plant Sci. 2004, 52, 109–114. [Google Scholar] [CrossRef]
- Lekhak, M.M.; Gondaliya, A.D.; Yadav, S.R.; Rajput, K.S. The stem anatomy at various developmental stages of secondary growth in Turbina corymbosa (L.) Raf. (Convolvulaceae). Plant Ecol. Evol. 2018, 151, 219–230. [Google Scholar] [CrossRef]
- Shelke, R.A.; Kapadane, K.K.; Ramoliya, D.G.; Gondaliya, A.D.; Rajput, K.S. Development of successive cambia and formation of secondary xylem in Suaeda nudiflora and S. fruticosa (Amaranthaceae s.l.). Flora. 2019, 256, 43–51. [Google Scholar] [CrossRef]
- Schenck, H. Beiträgezurbiologie und Anatomie der Lianen im besonderen der in Brasilien einheimischen Arten. In Botanische Mittheilungenaus den Tropen; Schimper,Gustav Fisher, Jena. 1893, 5, 1–271. [Google Scholar]
- Pfeiffer, H. Das abnorme. Dickenwachstum. In Hand buch der Pflanzenatomie.; Linsbauer, K., Ed.; Verlag von gebruder Borntraeger: Berlin, Germany, 1926; Volume IX. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons, Vol. II, 2nd ed.; Clarendon Press: Oxford, UK, 1983. [Google Scholar]
- Obaton, M. Les lianesligneuses à structure anormale des forêts denses d’Afrique Occidentale. Ann. Sci. Natur. Bot. Biol. Végé. 1960, 12, 1–20. [Google Scholar]
- Carlquist, S. Comparative Wood Anatomy: Systematic, Ecological and Evolutionary Aspect of Dicotyledon Wood; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1988; p. 385. [Google Scholar]
- Putz, H.A.; Mooney, F.E. The Biology of Vines; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1991. [Google Scholar]
- Rowe, N.P.; Speck, T. Biomechanical characteristics of the ontogeny and growth habit of the tropical liana Condylocarponguianense (Apocynaceae). Intl. J. Plant Sci. 1996, 157, 406–417. [Google Scholar] [CrossRef]
- Rowe, N.P.; Speck, T. Biomechanics of plant growth forms: The trouble with fossil plants. Rev. Plaeobot. Palynol. 1998, 102, 43–62. [Google Scholar] [CrossRef]
- Rowe, N.; Speck, T. Plant growth forms: An ecological and evolutionary perspective. New Phytol. 2005, 166, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Rowe, N.P.; Isnard, S.; Speck, T. Diversity of mechanical architecture in climbing plants: An evolutionary perspective. J. Plant Growth Regul. 2004, 23, 108–128. [Google Scholar] [CrossRef]
- Isnard, S.; Silk, W.K. Moving with climbing plants from Charles Darwin’s time into the 21st century. Amer. J. Bot. 2009, 96, 1205–1221. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A.; Bongers, F.; Burnham, R.J.; Putz, F.E. (Eds.) , Ecology of Lianas; John Wiley and Sons Ltd.: Oxford, UK, 2015; p. 512. [Google Scholar]
- Dillenburg, L.R.; Whigham, D.F.; Teramura, A.H.; Forseth, I.N. Effects of below- and aboveground competition from the vines Lonicera japonica and Parthenocissus quinquefolia on the growth of the tree host Liquidambar styraciflua. Oecologia 1993, 93, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Schnitzer, S.A.; Kuzee, M.E.; Bongers, F. Disentangling above-and below-ground competition between lianas and trees in a tropical forest. J. Ecol. 2005, 93, 1115–1125. [Google Scholar] [CrossRef] [Green Version]
- De Mirbel, M. M’emoire Sur lorigine, le development et I’organisattion du bios. Mémoire Du Mus. Hist. Nat. 1828, 16, 9–36. [Google Scholar]
- Gaudichaud, B.C. Observations surqulques points de physiologie et d’anatomie cpmparées des végétaux et specialement sur l’accroissement des tiges. Arch. Bot. 1833, 2, 481–500. [Google Scholar]
- De Jussieu, A. Monographie de la famille des Malpighiacées. Arch. Du Muséum D’histoire Nat. Paris 1843, 3, 5–151. [Google Scholar]
- Vesque, J. Mémoire sur l’anatomie comparé de l’ écorce. Ann. Des Sci. Natu. Bot. VI. 1875, 2, 82–198. [Google Scholar]
- Darwin, C. On the Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859; p. 440. [Google Scholar]
- Angyalossy, A.; Angeles, G.; Pace, M.R.; Lima, A.C.; Dias-Leme, C.L.; Lohmann, L.G.; Madero-Vega, C. An overview of the anatomy, development and evolution of the vascular system of lianas. Plant Ecol. Divers. 2012, 5, 167–182. [Google Scholar] [CrossRef]
- Angyalossy, V.; Pace, M.R.; Lima, A.C. Liana anatomy: A broad perspective on structural evolution of the vascular system. In Ecology of Lianas; Schnitzer, S.A., Bongers, F., Burnham, R.J., Putz, F.E., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 253–287. [Google Scholar]
- Chery, J.G.; Pace, M.R.; Acevedo-Rodríguez, P.; Specht, C.D.; Rothfels, C.J. Modifications during early plant development promote the evolution of nature’s most complex woods. Curr. Biol. 2020, 30, 237–244. [Google Scholar] [CrossRef]
- Caballé, G. Liana structure, function and selection: A comparative study of xylem cylinders of tropical rainforest species in Africa and America. Bot. J. Linn. Soc. 1993, 113, 41–60. [Google Scholar] [CrossRef]
- Pace, M.R.; Lohmann, L.G.; Angyalossy, V. The rise and evolution of the cambial variant in Bignonieae (Bignoniaceae). Evol. Dev. 2009, 11, 465–479. [Google Scholar] [CrossRef]
- Jacques, F.M.B.; De Franceschi, D. Menispermaceae wood anatomy and cambial variant. IAWA J. 2007, 28, 139–172. [Google Scholar] [CrossRef]
- Radlkofer, L. Monographie der Sapindaceen-GattungSerjania. In Verlag der Königl. Bayer; Akademie: Munich, Germany, 1875. [Google Scholar]
- Tamaio, N.; Somner, G.V. Development of corded vascular cylinder in Thinouia restingae Ferruci and Somner (Sapindaceae: Paullinieae). J. Torrey Bot. Soc. 2010, 137, 319–326. [Google Scholar] [CrossRef]
- Cunha Neto, I.L.; Martins, F.M.; Genise, V.; Somner, G.V.; Tamaio, N. Successive cambia in liana stems of Paullinieae and their evolutionary significance in Sapindaceae. Bot. J. Linn. Soc. 2018, 186, 66–88. [Google Scholar] [CrossRef]
- Dobbins, D.R. Studies on the anomalous cambial activity in Doxantha unguis-cati (Bignoniaceae). II. A case of differential production of secondary tissues. Amer. J. Bot. 1971, 58, 697–705. [Google Scholar] [CrossRef]
- Isnard, S.; Feild, T.S. The Evolution of Angiosperm Lianescence: A Perspective from Xylem Structure-Function; Ecology of Lianas. Wiley–Blackwell: Oxford, UK, 2015; pp. 221–250. [Google Scholar]
- Chalk, L.; Chattaway, M.M. Identification of woods with included phloem. Trop. Woods 1937, 50, 1–31. [Google Scholar]
- Cockrell, R.A. A comparative study of the wood structure of several South American species of Strychnos. Am. J. Bot. 1941, 28, 32–41. [Google Scholar] [CrossRef]
- Carlquist, S. Interxylary phloem: Diversity and functions. Brittonia 2013, 65, 477–495. [Google Scholar] [CrossRef]
- Bonnemain, J.L. Le phloème interne et le phloèmeinclus des dicotylédones: Leurhistogénèse et leurphysiologie. Rev. Gén. Bot. 1969, 76, 5–36. [Google Scholar]
- Philipson, W.R. Anomalous cambia. In The Vascular Cambium; Iqbal, M., Ed.; Research Studies Press: Taunton, UK, 1990; pp. 201–212. [Google Scholar]
- Philipson, W.R.; Butterfield, B.G.; Ward, J.M. The Vascular Cambium: Its Development and Activity; Chapman and Hall: London, UK, 1971. [Google Scholar]
- Den Outer, R.W.; Van Veenendaal, W.L.H. Development of included phloem in the stem of Combretum nigricans (Combretaceae). IAWA J. 1995, 16, 155–158. [Google Scholar] [CrossRef]
- Solereder, H. Systematic Anatomy of the Dicotyledons (Vol I); Boodle, L.A.; Fritsch, F.E.; Scott, D.H., Translators; Clarendon Press: Oxford, UK, 1908; pp. 562–575. [Google Scholar]
- Singh, B. Origin and distribution of inter-and intraxylary phloem in Leptadenia. Proc. Indian Acad. Sci. B 1943, 18, 14–19. [Google Scholar] [CrossRef]
- Luo, B.; Ou, Y.; Pan, B.; Qi, J.; Takao, I. The structure and development of interxylary and external phloem in Aquilaria sinensis. IAWA J. 2018, 39, 3–17. [Google Scholar] [CrossRef]
- Müller, F. Tiber das Holzeiniger um Desterro wachsenden Kletterpflanzen. Bot. Zeitung 1866, 68. Figure 10. [Google Scholar]
- Hérail, J. Recherches sur l’anatomie comparee de la tige des Dieotyledones. Ann. Sci. Nat. Bot. 1885, 2, 203–314. [Google Scholar]
- Van Veenendaal, W.L.H.; Den Outer, R.W. Development of included phloem and organization of the phloem network in the stem of Strychnos millepunctata (Loganiaceae). IAWA J. 1993, 14, 253–265. [Google Scholar] [CrossRef]
- Rajput, K.S.; Marcati, C.R. Stem anatomy and development of successive cambia in Hebanthe eriantha (Poir.) Pedersen: A neotropical climbing species of the Amaranthaceae. Plant Syst. Evol. 2013, 299, 1449–1459. [Google Scholar] [CrossRef]
- Cunha Neto, I.L.; Pace, M.; Hernandez-Gutierrez, R.; Angyalossy, A. Linking the evolution of development of stem vascular system in Nyctaginaceae and its correlation to habit and species diversification. EvoDevo 2022, 13, 4. [Google Scholar] [CrossRef]
- Carlquist, S. Successive cambia revisited: Ontogeny, histology, diversity and functional significance. J. Torrey Bot. Soc. 2007, 134, 301–332. [Google Scholar] [CrossRef]
- Rajput, K.S. Stem anatomy of Amaranthaceae: Rayless nature of xylem. Flora 2002, 197, 224–232. [Google Scholar] [CrossRef]
- Rajput, K.S.; Patil, V.S.; Rao, K.S. Wood anatomy and the development of interxylary phloem of Ipomoea hederifolia Linn. (Convolvulaceae). J. Plant Growth Regul. 2013, 32, 654–662. [Google Scholar] [CrossRef]
- Carlquist, S. Wood Anatomy of Onagraceae, with notes on alternative modes of photosynthate movement in dicotyledon woods. Ann. Missouri Bot. Garden 1975, 62, 386–424. [Google Scholar] [CrossRef]
- Mikesell, J.; Schroeder, A.C. Internal phloem development in Pharbitis nil Chois. (Convolvulaceae). Bot. Gaz. 1984, 145, 196–203. [Google Scholar] [CrossRef]
- Patil, V.S.; Rao, K.S.; Rajput, K.S. Development of intraxylary phloem and internal cambium in Ipomoea hederifolia (Convolvulaceae). J. Torrey Bot. Soc. 2009, 136, 423–432. [Google Scholar] [CrossRef]
- Rajput, K.S.; Gondaliya, A.D. Internal cambium and intraxylary phloem development in Ipomoea turbinata Lag. (Convolvulaceae). Flora 2017, 226, 47–54. [Google Scholar] [CrossRef]
- Terrazas, T.; Rodriguez, S.A.; Ojanguren, C.T. Development of successive cambia, cambial activity, and their relationship to physiological traits in Ipomoea arborescens (Convolvulaceae) seedlings. Am. J. Bot. 2011, 98, 765–774. [Google Scholar] [CrossRef]
- Rajput, K.S.; Lekhak, M.M.; Kapadane, K.K.; Yadav, S.R. Formation of tri-lobed stem and development of successive cambia in the stems of Argyreia hookeri C.B. Clarke (Convolvulaceae). Flora 2017, 233, 140–149. [Google Scholar] [CrossRef]
- Petersen, O.G. Ueber das Auftreten bicollateraler Gefassbiindel in verschiedenen Pflanzenfamilien und iiber den Werth derselben fiir die Systematik. Bot. Jahrb. 1882, 3, 359–402. [Google Scholar]
- Baranetzky, J. Recherches sur les faisceaux bicollateraux. Ann. Sci. Nat. Bot. 1900, 12, 261–332. [Google Scholar]
- Handa, T. Anatomical observations on the internal cambium of the stem in Marsdenia tomentose Morr. et Dence. Jap. J. Bot. 1937, 8, 59–64. [Google Scholar]
- Fischer, A. Untersuchungenuber das Siebrohren-System der Cucurbitaceen; Gebr. Borntraeger: Berlin, Germany, 1984. [Google Scholar]
- Schepper, V.; Swaef, T.; Bauweraerts, I.; Steppe, K. Phloem transport: A review of mechanisms and controls. J. Expt. Bot. 2013, 64, 4839–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoblauch, M.; Knoblauch, J.; Mullendore, D.L.; Savage, J.A.; Babst, B.A.; Beecher, S.D.; Dodgen, A.C.; Jensen, K.H.; Holbrook, N.M. Testing the Munch hypothesis of long-distance phloem transport in plants. eLife 2016, 5, e15341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esau, K. Plant Anatomy; John Wiley: New York, NY, USA, 1967. [Google Scholar]
- Beck, C.B. An Introduction to Plant Structure and Development, 2nd ed.; Cambridge University Press: New York, NY, USA, 2010; p. 433. [Google Scholar]
- Cunha Neto, I.L.; Pace, M.; Douglas, N.A.; Nee, M.H.; de Sá, C.F.C.; Moore, M.J.; Angyalossy, A. Diversity, distribution, development, and evolution of medullary bundles in Nyctaginaceae. Amer. J. Bot. 2020, 107, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Angyalossy, V.; Pace, M.R.; Evert, R.F.; Marcati, C.R.; Oskolski, A.A.; Terrazas, T.; Kotina, E.; Lens, F.P.; Mazzoni-Viveiros, S.C. IAWA list of Microscopic Bark Features. IAWA J. 2016, 37, 517–615. [Google Scholar] [CrossRef] [Green Version]
- Aloni, R. Phloem and Xylem differentiation. In Vascular Differentiation and Plant Hormones; Aloni, R., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2021; pp. 101–130. [Google Scholar]
- Aloni, R. Role of auxin and sucrose in the differentiation of sieve and tracheary elements in plant tissue cultures. Planta 1980, 150, 255–263. [Google Scholar] [CrossRef]
- Lev-Yadun, S.; Aloni, R. Polycentric vascular rays in Suaeda monoica and the control of ray initiation and spacing. Trees 1991, 5, 22–29. [Google Scholar] [CrossRef]
- Carlquist, S. Anatomy of vines and lianas: A review and synthesis. In Biology of Vines; Putz, F.E., Mooney, H.A., Eds.; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Bastos, C.L.; Tamaio, N.; Angyalossy, V. Unravelling roots of lianas: A case study in Sapindaceae. Ann. Bot. 2016, 118, 733–746. [Google Scholar] [CrossRef] [Green Version]
- Quintanar-Castilo, A.; Pace, M.R. Phloem wedges in Malpighiaceae: Origin, structure, diversification, and systematic relevance. EvoDevo 2022, 13, 11. [Google Scholar] [CrossRef]
- Pace, M.R.; Lohmann, L.G.; Angyalossy, V. Evolution of disparity between the regular and variant phloem in Bignonieae (Bignoniaceae). Am. J. Bot. 2011, 98, 602–618. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Name of the Taxon | Family | Habit | References for Further Information |
|---|---|---|---|---|
| 1 | Adansonia digitata L. | Malvaceae | Tree | [48] |
| 2 | Argyreia elliptica (Roth.) Choisy | Convolvulaceae | Woody climber | [49] |
| 3 | Argyreia nervosa (Burm f.) Bojer | Convolvulaceae | Woody climber | [50] |
| 4 | Argyreia osyrensis (Roth.) Choisy | Convolvulaceae | Woody climber | [51] |
| 5 | Argyreia sericea Dalzell and A. Gibson | Convolvulaceae | Woody climber | [51] |
| 6 | Argyreia splendense (Hornem) Sweet | Convolvulaceae | Woody climber | [51] |
| 7 | Arthrocnemum indicum (Willd.) Moq. | Amaranthaceae | Shrub | Present study |
| 8 | Barleria prionitis L. | Acanthaceae | Herb/shrub | Present study |
| 9 | Beaumontia jerdoniana Wight | Apocynaceae | Woody climber | [52] |
| 10 | Boerhavia diffusa L. | Nyctaginaceae | Diffuse herb | [53] |
| 11 | Bougainvillea sp. | Nyctaginaceae | Scandent shrub | Present study |
| 12 | Bombax ceiba L. | Malvaceae | Tree | [54] |
| 13 | Calycopteris floribunda (Roxb.) Lam. | Combretaceae | Woody lianas | [55] |
| 14 | Campsis radicans (L.) Bureau | Bignoniaceae | Woody climber | [56] |
| 15 | Camonea vitaefolia (Burm.f.) A.R Simoes and Staples | Convolvulaceae | Climber | Present study |
| 16 | Canavalia ensiformis (L.) DC. | Fabaceae | Annual climber | [57] |
| 17 | Canavalia gladiata (Jacq.) DC. | Fabaceae | Annual climber | Present study |
| 18 | Coccinia grandis (L.) Voigt | Cucurbitaceae | Climber | [58] |
| 19 | Dalechampia coriacea Klotzsch ex Müll.Arg. | Euphorbiaceae | Climber | Present study |
| 20 | Dicranostylis ampla Ducke | Convolvulaceae | Woody liana | [59] |
| 21 | Diplopterys carvalhoi W.R. Anderson and C.Davis | Malpighiaceae | Woody liana | Present study |
| 22 | Distimake tuberosus (L.) A.R. Simõesand Staples | Convolvulaceae | Woody liana | [60] |
| 23 | Dolichandra unguis-cati (L.) L.G. Lohmann | Bignoniaceae | Woody liana | Present study |
| 24 | Erythrina indica Lam. | Fabaceae | Tree | [54] |
| 25 | Gallesia integrifolia (Spreng.) Harms. | Phytolaccaceae | Tree | [61] |
| 26 | Heteropterys sp. | Malpighiaceae | Woody liana | Present study |
| 27 | Hewittia malabarica (L.) Suresh | Convolvulaceae | Woody climber | [62] |
| 28 | Ipomea hederifolia L. | Convolvulaceae | Annual climber | [63] |
| 29 | Ipomoea muricata (L.) Jacq. | Convolvulaceae | Climber | Present study |
| 30 | Ipomoea pes-caprae (L.) R.Br. | Convolvulaceae | Perennial climber | [64] |
| 31 | Ipomoea turbinata Lag. | Convolvulaceae | Annual climber | [65] |
| 32 | Jacquemontia pentantha (Jacq.) G.Don | Convolvulaceae | Perennial climber | Present study |
| 33 | Leptadenia reticulata (Retz.) Wight and Arn. | Apocynaceae (Asclepiadaceae) | Climber | [66] |
| 34 | Leptadenia reticulata (Retz.) Wight and Arn. and L. pyrotechnica (Forssk.) Decne. | Apocynaceae (Asclepiadaceae) | Climber and shrub respectively | [67] |
| 35 | Leptadenia pyrotechnica (Forssk.) Decne. | Apocynaceae (Asclepiadaceae) | Shrub | [68] |
| 36 | Mansoa alliacea (Lam.) A.H.Gentry (syn. Bignonia alliacea) | Bignoniaceae | Climber | Present study |
| 37 | Merremia hederacea (Burm.f.) Hallier f. | Convolvulaceae | Climber | Present study |
| 38 | Mucuna pruriens (L.) DC. | Fabaceae | Climber | Present study |
| 39 | Neuropeltis racemosa Wall. | Convolvulaceae | Woody liana | Present study |
| 40 | Phaseolus lunatus L. | Fabaceae | Annual climber | Present study |
| 41 | Pupalia lappacea (L.) Juss. | Amaranthaceae | Perennial shrub | [69] |
| 42 | Salvadora persica L. | Salvadoraceae | Tree | Present study |
| 43 | Salvadora oleoides Decne. | Salvadoraceae | Tree | Present study |
| 44 | Solanum pseudocapsicum L. | Solanaceae | Perennial shrub | [70] |
| 45 | Serjania mexicana (L.) Willd. | Sapindaceae | Woody climber | [71] |
| 46 | Strychnos bicolor Prog. | Loganiaceae | Straggling shrub | [72] |
| 47 | Strychnos bredemeyeri (Schult. and Schult. f.) Sprague and Sandwith | Loganiaceae | Straggling shrub | [73] |
| 48 | Strychnos potatorum L.f. | Loganiaceae | Tree | Present study |
| 49 | Sterculia urens Roxb. | Malvaceae (Sterculiaceae) | Tree | [74] |
| 50 | Strychnos potatorum L. | Loganiaceae | Tree | Unpublished data |
| 51 | Tectona grandis L.f. | Lamiaceae | Tree | [54] |
| 52 | Turbina corymbosa (L.) Raf. | Convolvulaceae | Perennial climber | [75] |
| 53 | Thunbergia grandiflora Roxb. | Acanthaceae | Climber | Present study |
| 54 | Suaeda fruticosa Forssk. ex J.F.Gmel. | Amaranthaceae | Small tree | [76] |
| 55 | Suaeda nudiflora Forssk. exJ.F.Gmel. | Amaranthaceae | Small tree | |
| 56 | Vallaris solanacea (Roth) Kuntze | Apocynaceae | Woody climber | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajput, K.S.; Kapadane, K.K.; Ramoliya, D.G.; Thacker, K.D.; Gondaliya, A.D. Inter- and Intraxylary Phloem in Vascular Plants: A Review of Subtypes, Occurrences, and Development. Forests 2022, 13, 2174. https://doi.org/10.3390/f13122174
Rajput KS, Kapadane KK, Ramoliya DG, Thacker KD, Gondaliya AD. Inter- and Intraxylary Phloem in Vascular Plants: A Review of Subtypes, Occurrences, and Development. Forests. 2022; 13(12):2174. https://doi.org/10.3390/f13122174
Chicago/Turabian StyleRajput, Kishore S., Kailas K. Kapadane, Dhara G. Ramoliya, Khyati D. Thacker, and Amit D. Gondaliya. 2022. "Inter- and Intraxylary Phloem in Vascular Plants: A Review of Subtypes, Occurrences, and Development" Forests 13, no. 12: 2174. https://doi.org/10.3390/f13122174