The Effects of Suillus luteus Inoculation on the Diversity of Fungal Communities and Their Structures in the Soil under Pinus massoniana Located in a Mining Area

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Soil Sampling
2.3. Physicochemical Soil Characterization
2.4. Determination of Enzyme Activity in Rhizospheric Soil
2.5. DNA Extraction and High-Throughput Sequencing
2.6. Statistical Analysis
3. Results
3.1. Heavy Metals and Enzymatic Activities
3.2. Rarefraction Measurement
3.3. Diversity and Richness
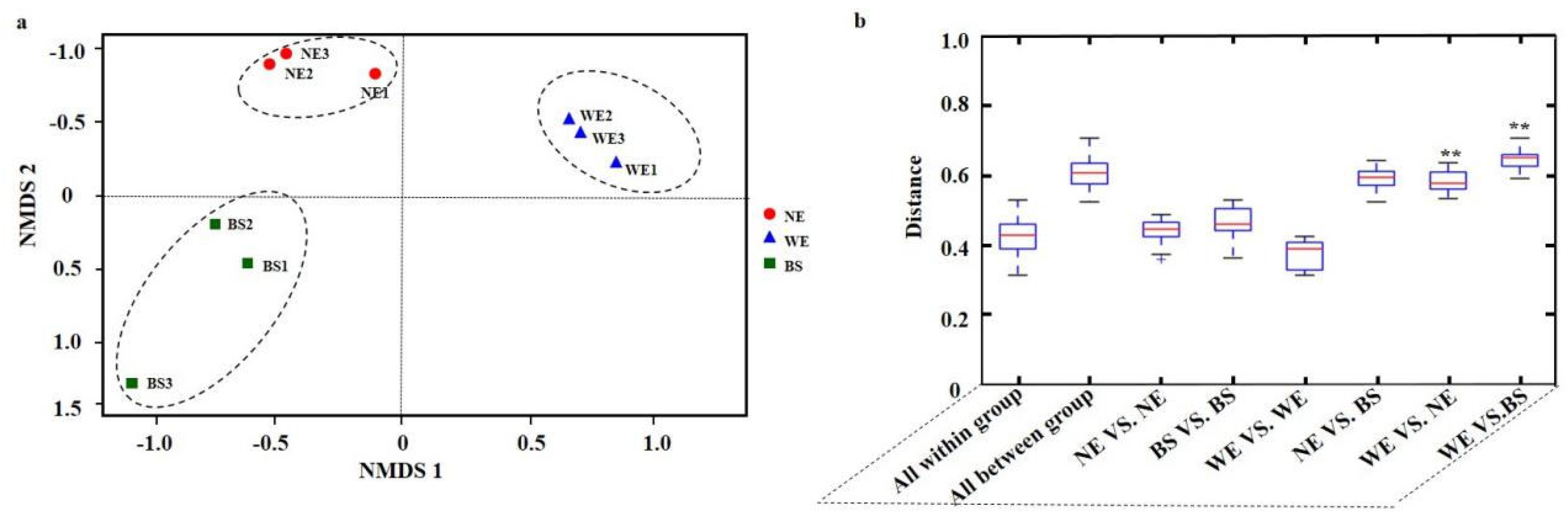
3.4. Variation in Fungal-Community Structure
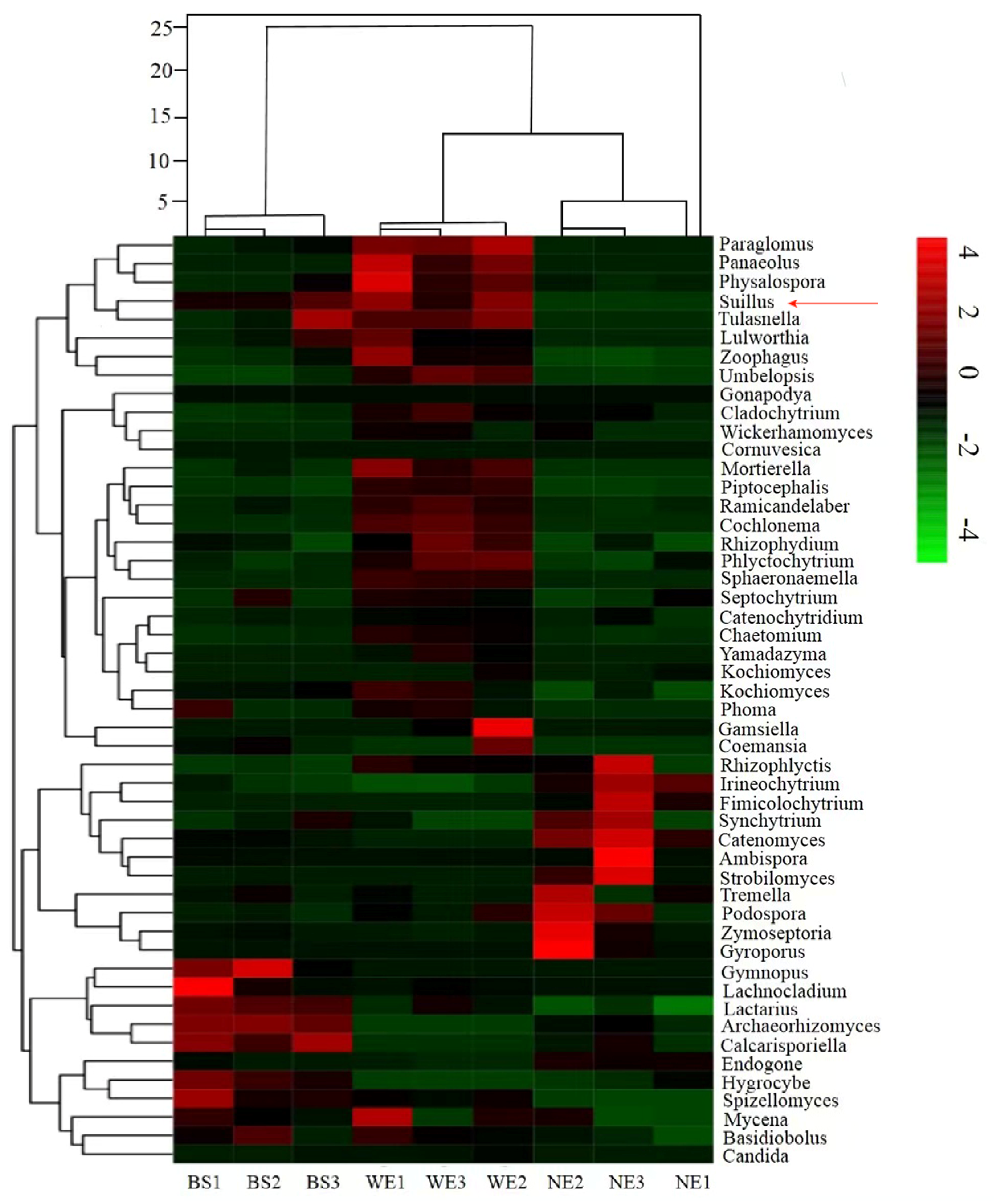
3.5. Phylum-Level Taxonomic Affiliation of OTUs
3.6. Responses of the Fungal Community in the Soil Inoculated with Ectomycorrhizal Fungi
3.7. Relationship between Fungal Communities and Environmental Variables
4. Discussion
4.1. Fungal Diversity of the Rhizosphere
4.2. Indicators of the Representative Fungal Genus
4.3. Relationships between Fungal-Community Structures and Soil Properties
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gall, J.E.; Boyd, R.S.; Rajakaruna, N. Transfer of heavy metals through terrestrial food webs: A review. Environ. Monit. Assess. 2015, 187, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, S.; Cao, Y.; Li, S.; Zhou, W.; Liu, S.; Bai, Z. Ecological Restoration of a Loess Open-Cast Mining Area in China: Perspective from an Ecological Security Pattern. Forests 2022, 13, 269. [Google Scholar] [CrossRef]
- Zhu, D.; Ke, X.; Wu, L.H.; Christie, P.; Luo, Y.M. Biological transfer of dietary cadmium in relation to nitrogen transfer and 15N fractionation in a soil collembolan-predatory mite food chain. Soil Biol. Biochem. 2016, 101, 207–216. [Google Scholar] [CrossRef]
- Chen, L.; Swenson, N.G.; Ji, N.; Mi, X.; Ren, H.; Guo, L.; Ma, K. Differential soil fungus accumulation and density dependence of trees in a subtropical forest. Science 2019, 366, 124–128. [Google Scholar] [CrossRef]
- Lim, B.S.; Kim, A.R.; Seol, J.; Oh, W.S.; An, J.H.; Lim, C.H.; Lee, C.S. Effects of Soil Amelioration and Vegetation Introduction on the Restoration of Abandoned Coal Mine Spoils in South Korea. Forests 2022, 13, 483. [Google Scholar] [CrossRef]
- Dixit, R.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.P.; Rai, J.P.; Sharma, P.K.; Lade, H.; et al. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef] [Green Version]
- Mueller, R.C.; Paula, F.S.; Mirza, B.S.; Rodrigues, J.L.; Nüsslein, K.; Bohannan, B.J. Links between plant and fungal communities across a deforestation chronosequence in the Amazon rainforest. ISME J. 2014, 8, 1548–1550. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Van den Hoogen, J.; Wan, J.; Mayes, M.A.; Keiser, A.D.; Mo, L.; Averill, C.; Maynard, D.S. The global soil community and its influence on biogeochemistry. Science 2019, 365, eaav0550. [Google Scholar] [CrossRef]
- Schneider, T.; Keiblinger, K.M.; Schmid, E.; Sterflinger-Gleixner, K.; Ellersdorfer, G.; Roschitzki, B.; Richter, A.; Eberl, L.; Zechmeister-Boltenstern, S.; Riedel, K. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. ISME J. 2012, 6, 1749–1762. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Niu, D.; Wang, Y.; Boydston, A.; Guo, D.; Li, X.; Wen, H.Y.; Qin, Y.; Fu, H. Litter decomposition in fenced and grazed grasslands: A test of the home-field advantage hypothesis. Geoderma 2019, 354, 113876. [Google Scholar] [CrossRef]
- Dean, S.L.; Farrer, E.C.; Taylor, D.L.; Porras-Alfaro, A.; Suding, K.N.; Sinsabaugh, R.L. Nitrogen deposition alters plant–fungal relationships: Linking belowground dynamics to aboveground vegetation change. Mol. Ecol. 2014, 23, 1364–1378. [Google Scholar] [CrossRef]
- Wilgan, R.; Leski, T. Ectomycorrhizal Assemblages of Invasive Quercus rubra L. and Non-Invasive Carya Nutt. Trees under Common Garden Conditions in Europe. Forests 2022, 13, 676. [Google Scholar] [CrossRef]
- Sousa, N.R.; Ramos, M.A.; Marques, A.P.; Castro, P.M. The effect of ectomycorrhizal fungi forming symbiosis with Pinus pinaster seedlings exposed to cadmium. Sci. Total Environ. 2012, 414, 63–67. [Google Scholar] [CrossRef]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Thion, C.; Cébron, A.; Beguiristain, T.; Leyval, C. Long-term in situ dynamics of the fungal communities in a multi-contaminated soil are mainly driven by plants. FEMS Microbiol. Ecol. 2012, 82, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kang, W.; Liu, S.; Yin, H.; Lyu, Q.; Su, Y.; Liu, J.J.; Liu, J.L.; Fan, C.; Chen, G.; et al. Diversity of Ectomycorrhizal Fungal Communities in Four Types of Stands in Pinus massoniana Plantation in the West of China. Forests 2021, 12, 719. [Google Scholar] [CrossRef]
- Peay, K.G. The mutualistic niche: Mycorrhizal symbiosis and community dynamics. Annual Review of Ecology. Evol. Syst. 2016, 47, 143–164. [Google Scholar] [CrossRef]
- Liu, J.; Ha, V.N.; Shen, Z.; Dang, P.; Zhu, H.; Zhao, F.; Zhao, Z. Response of the rhizosphere microbial community to fine root and soil parameters following Robinia pseudoacacia L. afforestation. Appl. Soil Ecol. 2018, 132, 11–19. [Google Scholar] [CrossRef]
- Fernandez, C.W.; Kennedy, P.G. Revisiting the ‘Gadgil effect’: Do interguild fungal interactions control carbon cycling in forest soils? New Phytol. 2016, 209, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Nara, K.; Lian, C.; Zong, K.; Peng, K.; Xue, S.; Shen, Z. Ectomycorrhizal fungal communities associated with Masson pine (Pinus massoniana Lamb.) in Pb-Zn mine sites of central south China. Mycorrhiza 2012, 22, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Li, N.Y.; Guo, B.; Li, H.; Fu, Q.L.; Feng, R.W.; Ding, Y.Z. Effects of double harvesting on heavy metal uptake by six forage species and the potential for phytoextraction in field. Pedosphere 2016, 26, 717–724. [Google Scholar] [CrossRef]
- Shi, L.; Deng, X.; Yang, Y.; Jia, Q.; Wang, C.; Shen, Z.; Chen, Y. A Cr (VI)-tolerant strain, Pisolithus sp1, with a high accumulation capacity of Cr in mycelium and highly efficient assisting Pinus thunbergii for phytoremediation. Chemosphere 2019, 224, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Policelli, N.; Bruns, T.D.; Vilgalys, R.; Nuñez, M.A. Suilloid fungi as global drivers of pine invasions. New Phytol. 2019, 222, 714–725. [Google Scholar] [CrossRef]
- Colpaert, J.V.; Wevers, J.H.; Krznaric, E.; Adriaensen, K. How metal-tolerant ecotypes of ectomycorrhizal fungi protect plants from heavy metal pollution. Ann. For. Sci. 2011, 68, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Op De Beeck, M.; Lievens, B.; Busschaert, P.; Rineau, F.; Smits, M.; Vangronsveld, J.; Colpaert, J.V. Impact of metal pollution on fungal diversity and community structures. Environ. Microbiol. 2014, 17, 2035–2047. [Google Scholar] [CrossRef]
- Yu, P.; Sun, Y.; Huang, Z.; Zhu, F.; Sun, Y.; Jiang, L. The effects of ectomycorrhizal fungi on heavy metals’ transport in Pinus massoniana and bacteria community in rhizosphere soil in mine tailing area. J. Hazard. Mater. 2020, 381, 121203. [Google Scholar] [CrossRef]
- Li, X.; Rui, J.; Mao, Y.; Yannarell, A.; Mackie, R. Dynamics of the bacterial community structure in the rhizosphere of a maize cultivar. Soil Biol. Biochem. 2014, 68, 392–401. [Google Scholar] [CrossRef]
- Bao, S.D. The Soil Agricultural Chemistry Analysis; Chinese Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Guan, S.Y. Soil Enzyme and Its Research Methods; Agricultural Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340. [Google Scholar] [CrossRef]
- Hu, H.; Jiang, C.; Wu, Y.P.; Cheng, Y.X. Bacterial and fungal communities and contribution of physicochemical factors during cattle farm waste composting. MicrobiologyOpen 2017, 6, e00518. [Google Scholar]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Demenois, J.; Carriconde, F.; Bonaventure, P.; Maeght, J.L.; Stokes, A.; Rey, F. Impact of plant root functional traits and associated mycorrhizas on the aggregate stability of a tropical Ferralsol. Geoderma 2018, 312, 6–16. [Google Scholar] [CrossRef]
- Sun, Y.J.; Yu, P.Y.; Chen, J.Z.; Li, S.M.; Jiang, L.J. Effects of slippery jack (Suillus luteus) on the heavy metal accumulation and soil properties of masson’s pine (pinus massoniana lamb) in a mining area of china. Appl. Ecol. Environ. Res. 2020, 18, 3741–3755. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Hannula, S.E.; De Boer, W.; Van Veen, J.A. In situ dynamics of soil fungal communities under different genotypes of potato, including a genetically modified cultivar. Soil Biol. Biochem. 2010, 42, 2211–2223. [Google Scholar] [CrossRef]
- De Miguel, A.M.; Águeda, B.; Sánchez, S.; Parladé, J. Ectomycorrhizal fungus diversity and community structure with natural and cultivated truffle hosts: Applying lessons learned to future truffle culture. Mycorrhiza 2014, 24, 5–18. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, J.; Xiong, C.; Li, X.; Chen, Z.; Li, P.; Huang, W. Tuber indicum shapes the microbial communities of ectomycorhizosphere soil and ectomycorrhizae of an indigenous tree (Pinus armandii). PLoS ONE 2017, 12, e0175720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Wang, X.H.; Li, J.G.; Qin, W.; Xiao, C.Z.; Zhao, X.; Jiang, H.X.; Sui, J.K.; Sa, R.B.; Wang, W.Y.; et al. Pyrosequencing Reveals Fungal Communities in the Rhizosphere of Xinjiang Jujube. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gosling, P.; Proctor, M.; Jones, J.; Bending, G.D. Distribution and diversity of Paraglomus spp. in tilled agricultural soils. Mycorrhiza 2014, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.S.; Li, G.H.; Zou, Y.N. Roles of arbuscular mycorrhizal fungi on growth and nutrient acquisition of peach (Prunus persica L. Batsch) seedlings. J. Anim. Plant Sci. 2011, 21, 746–750. [Google Scholar]
- Aggarwal, A.; Kadian, N.; Tanwar, A.; Yadav, A.; Gupta, K.K. Role of arbuscular mycorrhizal fungi (AMF) in global sustainable development. J. Appl. Nat. Sci. 2011, 3, 340–351. [Google Scholar] [CrossRef]
- Giordano, L.; Gonthier, P.; Lione, G.; Garbelotto, M. The invasiveness of a non-native fungal forest pathogen is boosted by the presence of a congeneric native species. For. Int. J. For. Res. 2019, 92, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Lastovetsky, O.A.; Gaspar, M.L.; Mondo, S.J.; LaButti, K.M.; Sandor, L.; Grigoriev, I.V.; Henry, S.A.; Pawlowska, T.E. Lipid metabolic changes in an early divergent fungus govern the establishment of a mutualistic symbiosis with endobacteria. Proc. Natl. Acad. Sci. USA 2016, 113, 15102–15107. [Google Scholar] [CrossRef] [Green Version]
- Kyaschenko, J.; Clemmensen, K.E.; Hagenbo, A.; Karltun, E.; Lindahl, B.D. Shift in fungal communities and associated enzyme activities along an age gradient of managed Pinus sylvestris stands. ISME J. 2017, 11, 863–874. [Google Scholar] [CrossRef]
- Bödeker, I.T.; Lindahl, B.D.; Olson, Å.; Clemmensen, K.E. Mycorrhizal and saprotrophic fungal guilds compete for the same organic substrates but affect decomposition differently. Funct. Ecol. 2016, 30, 1967–1978. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Azarbad, H.; Niklińska, M.; van Gestel, C.A.; van Straalen, N.M.; Röling, W.F.; Laskowski, R. Microbial community structure and functioning along metal pollution gradients. Environ. Toxicol. Chem. 2013, 32, 1992–2002. [Google Scholar] [CrossRef]
- Luo, Z.B.; Wu, C.H.; Zhang, C.; Li, H.; Lipka, U.; Polle, A. The role of ectomycorrhizas in heavy metal stress tolerance of host plants. Environ. Exp. Bot. 2014, 108, 47–62. [Google Scholar] [CrossRef]
- Liang, M.; Johnson, D.; Burslem, D.F.; Yu, S.; Fang, M.; Taylor, J.D.; Helgason, T.; Liu, X. Soil fungal networks maintain local dominance of ectomycorrhizal trees. Nat. Commun. 2020, 11, 1–7. [Google Scholar] [CrossRef]
- Shi, L.; Xue, J.; Liu, B.; Dong, P.; Wen, Z.; Shen, Z.; Chen, Y. Hydrogen ions and organic acids secreted by ectomycorrhizal fungi, Pisolithus sp1, are involved in the efficient removal of hexavalent chromium from waste water. Ecotoxicol. Environ. Saf. 2018, 161, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wen, X.P.; Ding, G.J. Ectomycorrhizal symbiosis enhances tolerance to low phosphorous through expression of phosphate transporter genes in masson pine (Pinus massoniana). Acta Physiol. Plant. 2017, 39, 101. [Google Scholar] [CrossRef]
- Eichlerová, I.; Homolka, L.; Žifčáková, L.; Lisá, L.; Dobiášová, P.; Baldrian, P. Enzymatic systems involved in decomposition reflects the ecology and taxonomy of saprotrophic fungi. Fungal Ecol. 2015, 13, 10–22. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi-potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef]
- Burke, D.J.; Weintraub, M.N.; Hewins, C.R.; Kalisz, S. Relationship between soil enzyme activities, nutrient cycling and soil fungal communities in a northern hardwood forest. Soil Biol. Biochem. 2011, 43, 795–803. [Google Scholar] [CrossRef]
- Kohler, A.; Kuo, A.; Nagy, L.G.; Morin, E.; Barry, K.W.; Buscot, F.; Canbäck, B.; Choi, C.; Cichocki, N.; Clum, A.; et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 2015, 47, 410–415. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (mg/kg) | Enzymatic Activities (mg/g) | |||||
|---|---|---|---|---|---|---|
| Pb | Zn | CAT | AKP | URE | SAC | |
| WE | 64.27 ± 10.85 b | 27.47 ± 2.55 c | 2.81 ± 0.14 a | 0.28 ± 0.02 a | 0.29 ± 0.02 a | 23.19 ± 1.61 a |
| NE | 66.20 ± 0.81 b | 193.47 ± 9.51 b | 1.97 ± 0.25 b | 0.17 ± 0.03 b | 0.22 ± 0.02 ab | 16.37 ± 0.94 b |
| BS | 125.33 ± 4.49 a | 233.40 ± 23.81 a | 1.08 ± 0.70 c | 0.12 ± 0.01 b | 0.13 ± 0.03 b | 7.28 ± 1.03 c |
| Simpson | Chao1 | ACE | Shannon | |
|---|---|---|---|---|
| Non ECMF | 0.95 ± 0.01 a | 1173.33 ± 75.12 a | 1174.63 ± 71.85 a | 6.56 ± 0.08 ab |
| With ECMF | 0.96 ± 0.01 a | 1143.60 ± 49.48 a | 1049.36 ± 54.31 a | 6.87 ± 0.19 a |
| Bulk Soil | 0.73 ± 0.14 a | 791.87 ± 154.78 b | 792.81 ± 154.06 b | 4.60 ± 0.98 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, P.; Ning, C.; Chen, J.; Zhu, F.; Sun, Y.; Shen, A.; Zeng, W.; Jiang, L. The Effects of Suillus luteus Inoculation on the Diversity of Fungal Communities and Their Structures in the Soil under Pinus massoniana Located in a Mining Area. Forests 2022, 13, 2162. https://doi.org/10.3390/f13122162
Yu P, Ning C, Chen J, Zhu F, Sun Y, Shen A, Zeng W, Jiang L. The Effects of Suillus luteus Inoculation on the Diversity of Fungal Communities and Their Structures in the Soil under Pinus massoniana Located in a Mining Area. Forests. 2022; 13(12):2162. https://doi.org/10.3390/f13122162
Chicago/Turabian StyleYu, Peiyi, Chen Ning, Jingzhen Chen, Fan Zhu, Yujing Sun, Airong Shen, Wenbin Zeng, and Lijuan Jiang. 2022. "The Effects of Suillus luteus Inoculation on the Diversity of Fungal Communities and Their Structures in the Soil under Pinus massoniana Located in a Mining Area" Forests 13, no. 12: 2162. https://doi.org/10.3390/f13122162