The Long-Lasting Territories of Forest Apex Predators Sustain Diverse Bird Communities throughout the Year


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Historic and Contemporary Distribution
2.3. Bird Survey
2.4. Bird Indices
2.5. Environmental Variables
2.6. Statistical Analyses
3. Results
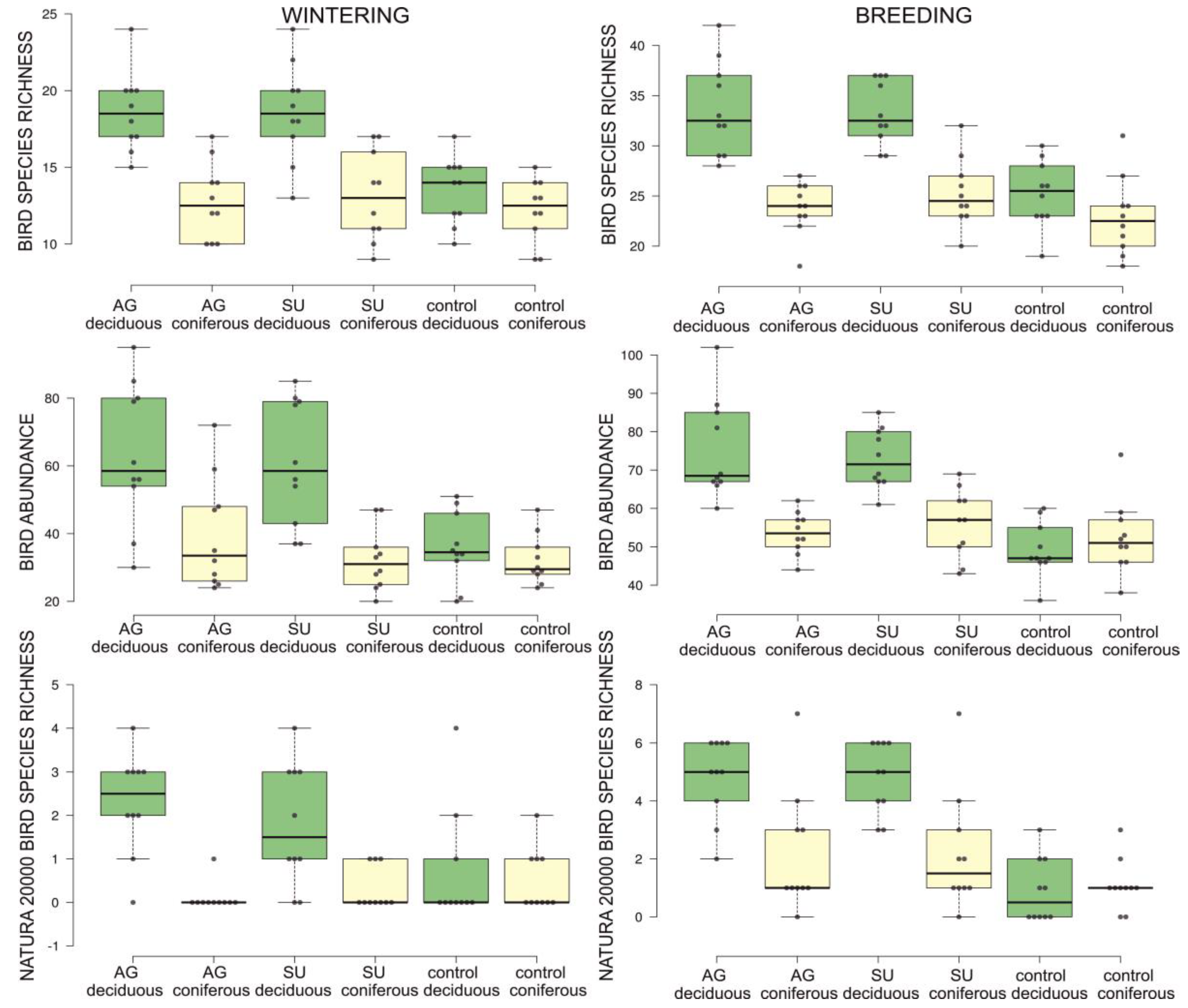
3.1. Bird Diversity Metrics in Raptor Territories
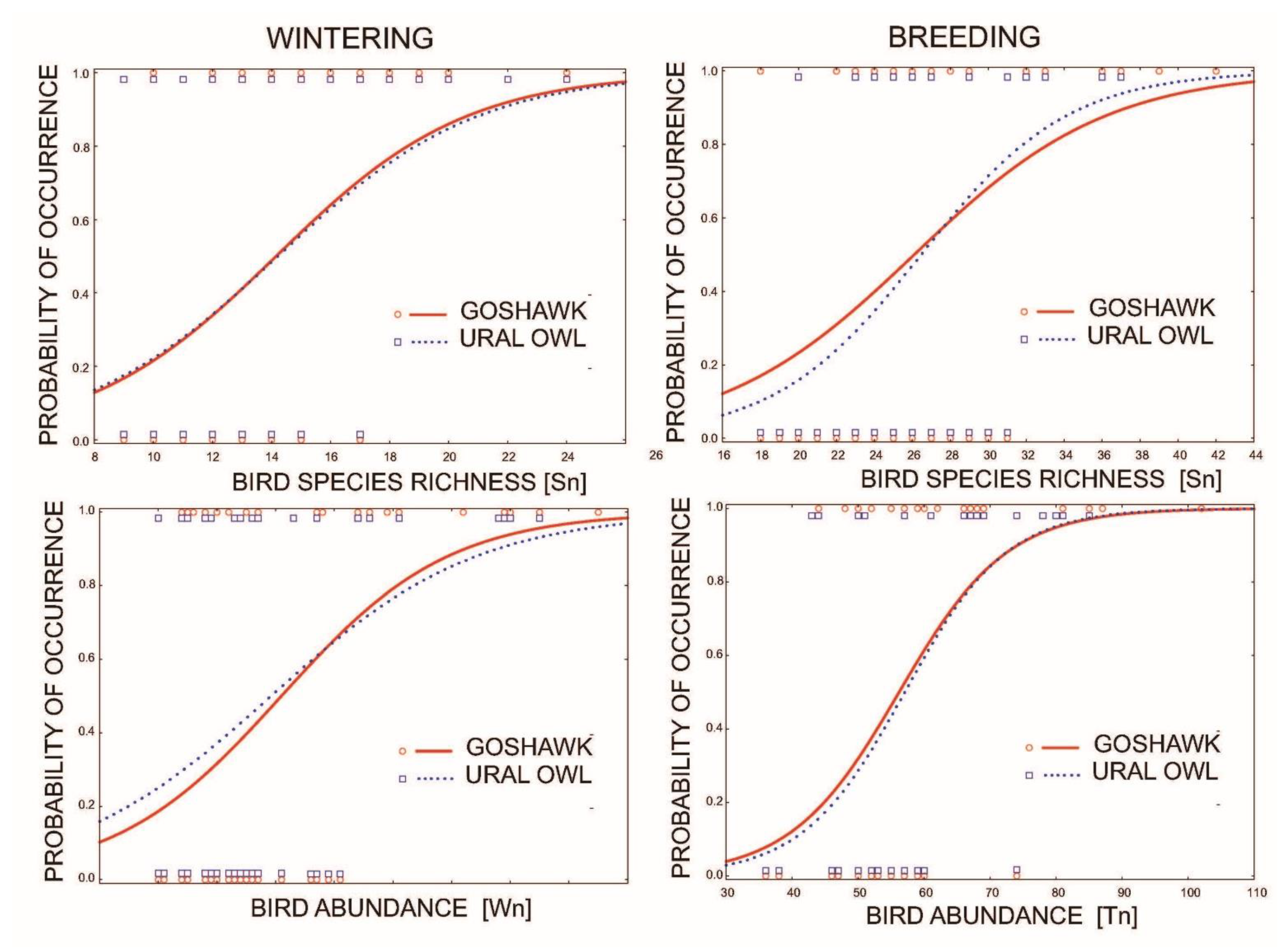
3.2. Diverity of Birds in Goshawk and Owl Territories
3.3. Environmental Variables in Raptor Territories
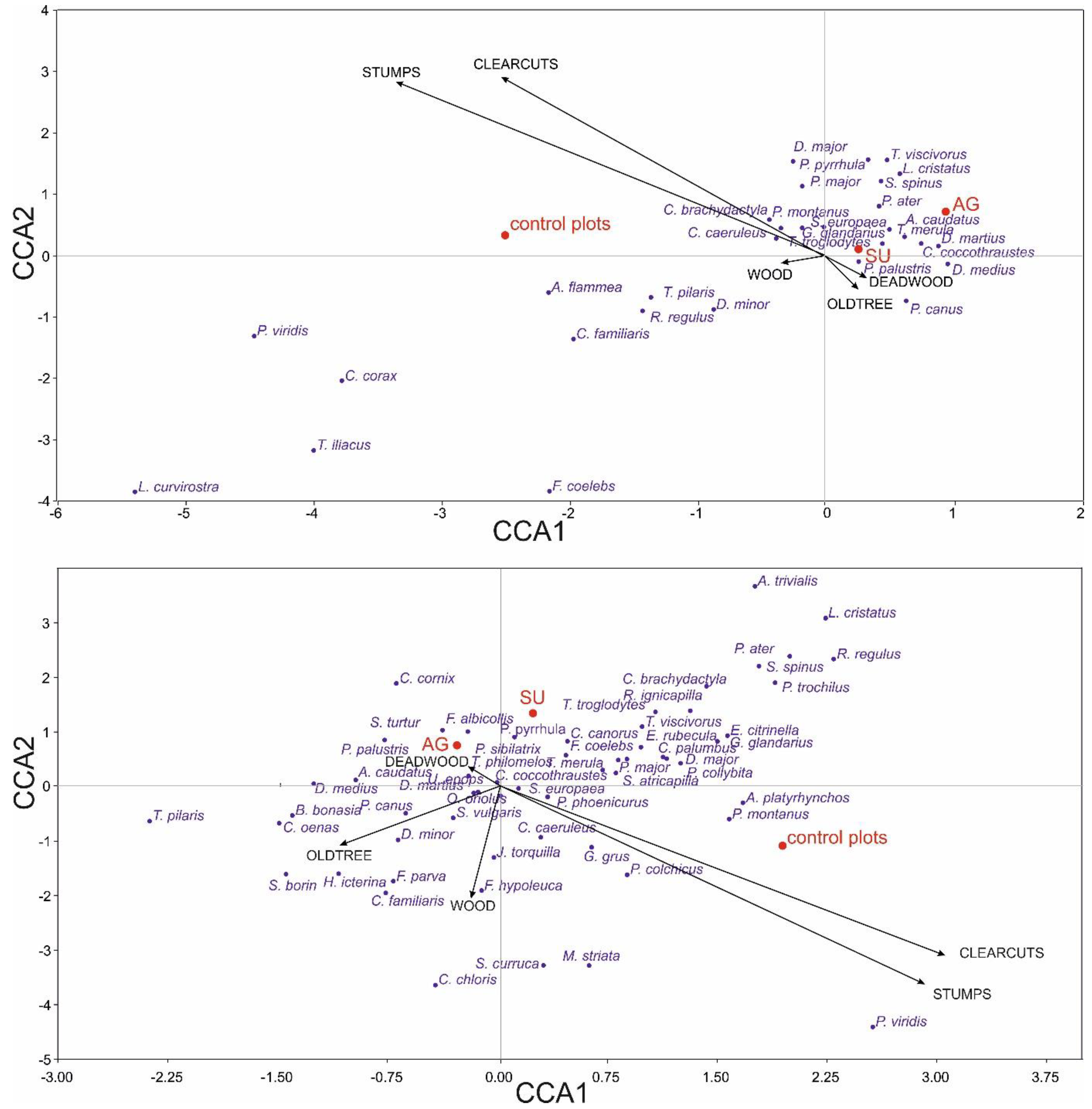
3.4. Diverity of Birds in Relation to Environment and Raptor Presence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andelman, S.J.; Fagan, W.F. Umbrellas and flagships: Efficient conservation surrogates or expensive mistakes? Proc. Natl. Acad. Sci. USA 2000, 97, 5954–5959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberge, J.M.; Angelstam, P. Usefulness of the umbrella species concept as a conservation tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Natsukawa, H.; Sergio, F. Top predators as biodiversity indicators: A meta-analysis. Ecol. Lett. 2022, 25, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Sergio, F.; Newton, I.; Marchesi, L. Top predators and biodiversity. Nature 2005, 436, 192. [Google Scholar] [CrossRef] [Green Version]
- Sergio, F.; Newton, I.; Marchesi, L.; Pedrini, P. Ecologically justified charisma: Preservation of top predators delivers biodiversity conservation. J. Appl. Ecol. 2006, 43, 1049–1055. [Google Scholar] [CrossRef] [Green Version]
- Roth, T.; Weber, D. Top predators as indicators for species richness? Prey species are just as useful. J. App. Ecol. 2008, 45, 987–991. [Google Scholar]
- Canterbury, G.E.; Martin, T.E.; Petit, D.R.; Petit, L.J.; Bradford, D.F. Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conserv. Biol. 2000, 14, 544–558. [Google Scholar] [CrossRef]
- Sergio, F.; Caro, T.; Brown, D.; Clucas, B.; Hunter, J.; Ketchum, J.; McHugh, K.; Hiraldo, F. Top predators as conservation tools: Ecological rationale, assumptions, and efficacy. Ann. Rev. Ecol. Evol. Syst. 2008, 39, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Sergio, F.; Newton, I.; Marchesi, L. Top predators and biodiversity: Much debate, few data. J. App. Ecol. 2008, 45, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Wiens, J.A.; Hayward, G.D.; Holthausen, R.S.; Wisdom, M.J. Using surrogate species and groups for conservation planning and management. BioScience 2008, 58, 241–252. [Google Scholar] [CrossRef]
- Burgas, D.; Byholm, P.; Parkkima, T. Raptors as surrogates of biodiversity along a landscape gradient. J. Appl. Ecol. 2014, 51, 786–794. [Google Scholar] [CrossRef]
- Senzaki, M.; Yamaura, Y. Surrogate species versus landscape metric: Does presence of a raptor species explains diversity of multiple taxa more than patch area? Wetl. Ecol. Manag. 2016, 24, 427–441. [Google Scholar] [CrossRef]
- Lyly, M.S.; Villers, A.; Koivisto, E.; Helle, P.; Ollila, T.; Korpimäki, E. Guardian or threat: Does golden eagle predation risk have cascading effects on forest grouse? Oecologia 2016, 182, 487–498. [Google Scholar] [CrossRef]
- Hakkarainen, H.; Mykrä, S.; Kurki, S.; Tornberg, R.; Jungell, S.; Nikula, A. Long-term change in territory occupancy pattern of Goshawks (Accipiter gentilis). Ecoscience 2004, 11, 399–403. [Google Scholar] [CrossRef]
- Sarasola, J.H.; Grande, J.M.; Negro, J.J. (Eds.) Birds of Prey Biology and Conservation in the XXI Century; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Sergio, F.; Blas, J.; Baos, R.; Forero, M.G.; Donázar, J.A.; Hiraldo, F. Short- and long-term consequences of individual and territory quality in a long-lived bird. Oecologia 2009, 160, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Rodenhouse, N.L.; Sherry, T.W.; Holmes, R.T. Site-dependent regulation of population size: A new synthesis. Ecology 1997, 78, 2025–2042. [Google Scholar] [CrossRef]
- Pärt, T. Experimental evidence of environmental effects on age-specific reproductive success: The importance of resource quality. Proc. R. Soc. London Ser. B 2001, 268, 2267–2271. [Google Scholar] [CrossRef] [Green Version]
- Natsukawa, H. Raptor breeding sites indicate high taxonomic and functional diversities of wintering birds in urban ecosystems. Urban Urban Green 2021, 60, 127066. [Google Scholar] [CrossRef]
- Kostrzewa, R.; Kostrzewa, A. Winter weather, spring and summer density, and subsequent breeding success of Eurasian kestrels, common buzzards, and northern goshawks. Auk 1991, 108, 342–347. [Google Scholar]
- Reynolds, R.T.; Lambert, J.S.; Flather, C.H.; White, G.C.; Bird, B.J.; Baggett, L.S.; Lambert, C.; Bayard De Volo, S. Long-term demography of the Northern Goshawk in a variable environment. Wildl. Monogr. 2017, 197, 1–40. [Google Scholar] [CrossRef]
- Brambilla, M.; Bassi, E.; Ceci, C.; Rubolini, D. Environmental factors affecting patterns of distribution. Ibis 2010, 152, 310–322. [Google Scholar] [CrossRef]
- Chakarov, N. Krüger, O.; Peter, H.U. Mesopredator release by an emergent superpredator: A natural experiment of predation in a three-level guild. PLoS ONE 2010, 5, e15229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solonen, T. Impact of dominant predators on territory occupancy and reproduction of subdominant ones within a guild of birds of prey. Open Ornith. J. 2011, 4, 23–29. [Google Scholar] [CrossRef]
- Byholm, P.; Burgas, D.; Virtanen, T.; Valkama, J. Competitive exclusion within the predator community influences the distribution of a threatened prey species. Ecology 2012, 93, 1802–1808. [Google Scholar] [CrossRef] [Green Version]
- Burgas, D.; Juutinen, A.; Byholm, P. The cost effectiveness of using raptor nest sites to identify areas with high species richness of other taxa. Ecol. Indic. 2016, 70, 518–530. [Google Scholar] [CrossRef]
- Czuchnowski, R. Breeding ecology of the Goshawk, Accipiter gentilis, in the Niepołomice Forest. Remiz 1992, 1, 8–11. [Google Scholar]
- Bielański, W. Nesting preferences of Common Buzzard Buteo buteo and Goshawk Accipiter gentilis in forest stands of different structure (Niepołomice Forest, Southern Poland). Biologia 2006, 61, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Kajtoch, Ł. Raptors Falconiformes of Bocheński and Wielicko-Wiśnicki Foothills and Beskid Wyspowy Mts. Kulon 2009, 14, 81–90. [Google Scholar]
- Wiehle, D.; Wyka, J.; Matysek, M.; Sobas, P.; Kusal, B.; Kajtoch, Ł. Distribution and numbers of goshawks Accipiter gentilis in middle part of the Małopolska province. Ornis Pol. 2020, 61, 47–56. [Google Scholar]
- Czuchnowski, R. Ecology of the Ural owl Strix uralensis in the Niepołomice Forest. Remiz 1993, 2, 7–12. [Google Scholar]
- Kus, K.; Szczepaniak, P.; Bracik, T.; Mędrzak, R. Inwentaryzacja Ornitologiczna Wybranych Gatunków Ptaków Obszaru Europejskiej Sieci Ekologicznej Natura 2000 Puszcza Niepołomicka PLB120002; Fundacji Wspierania Inicjatyw Ekologicznych i Stowarzyszenia Ochrony Sów: Kraków, Poland, 2008. [Google Scholar]
- Kajtoch, Ł. Owls Strigidae of Wieliczka-Wiśnicz Foothills and Beskid Wyspowy Mts. Not. Ornit. 2006, 47, 252–259. [Google Scholar]
- Matysek, M.; Figarski, T.; Wieczorek, P.; Wyka, J.; Kajtoch, Ł. Distribution of tawny owl Strix aluco and Ural owl Strix uralensis in fragmented forests of the middle part of Małopolska province. Ornis Pol. 2015, 56, 287–297. [Google Scholar]
- Wiehle, D.; Sobas, P. Rozmieszczenie i liczebność puszczyka uralskiego Strix uralensis w Puszczy Niepołomickiej. Ornis Pol. 2018, 59, 107–118. [Google Scholar]
- Bibby, C.; Burguess, N.D.; Hill, D.A. Bird Census Techniques; Academic Press: London, UK, 1992; p. 257. [Google Scholar]
- Gregory, R.D.; Gibbons, D.W.; Donald, P.F. Bird Census and Survey Techniques. In Bird Ecology and Conservation a Handbook of Techniques; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Fedyń, I.; Grzędzicka, E.; Wiehle, D.; Kajtoch, Ł. The co-occurrence of Ural Owl and Northern Goshawk differs between extensive and fragmented forests. Bird Study 2021, 68, 267–278. [Google Scholar] [CrossRef]
- Colwell, R.K. Estimates Statistical Estimation of Species Richness and Shared Species from Samples. In Version 9. User’s Guide and Application; 2013; Available online: http://purl.oclc.org/estimates (accessed on 10 August 2022).
- Lebreton, J.-D.; Burnham, K.P.; Clobert, J.; Anderson, D.R. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case-studies. Ecol. Monogr. 1992, 62, 67e118. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Multimodel inference. Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 10 August 2022).
- StatSoft, Inc. Electronic Statistics Textbook; StatSoft: Tulsa, OK, USA, 2012; Available online: http://www.statsoft.com/textbook/ (accessed on 10 August 2022).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Askeyev, A.; Askeyev, O.; Askeyev, I. Owls as bioindicators: Their spatial and temporal aspects in Eastern Europe. Eur. J. Ecol. 2019, 5, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Byholm, P.; Gunko, R.; Burgas, D.; Karell, P. Losing your home: Temporal changes in forest landscape structure due to timber harvest accelerate Northern goshawk (Accipiter gentilis) nest stand losses. Ornis Fennica 2020, 97, 1–11. [Google Scholar]
- Natsukawa, H.; Yuasa, H.; Komuro, S.; Sergio, F. Raptor breeding sites indicate high plant biodiversity in urban ecosystems. Sci. Rep. 2021, 11, 21139. [Google Scholar] [CrossRef] [PubMed]
- Penteriani, V.; Faivre, B. Effects of harvesting timber stands on Goshawk nesting in two European areas. Biol. Conserv. 2001, 101, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Finn, S.P.; Marzluff, J.M.; Varland, D.E. Effects of landscape and local habitat attributes on Northern Goshawk site occupancy in western Washington. For. Sci. 2002, 48, 427–436. [Google Scholar]
- Lõhmus, A. Do Ural Owls (Strix uralensis) suffer from the lack of nest sites in managed forests? Biol. Conserv. 2003, 110, 1–9. [Google Scholar] [CrossRef]
- Saga, Ø.; Selås, V. Nest reuse by Goshawks after timber harvesting: Importance of distance to logging, remaining mature forest area and tree species composition. For. Ecol. Manag. 2012, 270, 66–70. [Google Scholar] [CrossRef]
- Björklund, H.; Parkkinen, A.; Hakkari, T.; Heikkinen, R.K.; Virkkala, R.; Lensu, A. Predicting valuable forest habitats using an indicator species for biodiversity. Biol. Conserv. 2020, 249, 108682. [Google Scholar] [CrossRef]
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Martí, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; et al. European Breeding Bird Atlas 2: Distribution, Abundance and Change; European Bird Census Council & Lynx Edicions: Barcelona, Spain, 2020. [Google Scholar]
- Tomiałojć, L.; Stawarczyk, T. Awifauna Polski. Rozmieszczenie, liczebność i zmiany. PTPP „pro Natura”, Wrocław. Not. Orn. 2003, 45, 274–276. [Google Scholar]
- Vrezec, A. Breeding density and altitudinal distribution of the Ural, tawny, and boreal owls in North Dinaric Alps (Central Slovenia). J. Raptor Res. 2003, 37, 55–62. [Google Scholar]
- Vrezec, A.; Tome, D. Altitudinal segregation between Ural Owl Strix uralensis and Tawny Owl Strix aluco: Evidence for competitive exclusion in raptorial birds. Bird Study 2004, 51, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Vrezec, A.; Tome, D. Habitat selection and patterns of distribution in a hierarchic forest owl guild. Ornis Fenn. 2004, 81, 109–118. [Google Scholar]
- Bylicka, M.; Kajtoch, Ł.; Figarski, T. Habitat and landscape characteristics affecting the Ural Owl Strix uralensis occurrence in agro-forestry mosaic. Acta Ornith. 2010, 45, 33–42. [Google Scholar] [CrossRef]
- Matuszkiewicz, J.M. Zespoły leśne Polski; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2001. [Google Scholar]
- Wilk, T.; Jujka, M.; Krogulec, J.; Chylarecki, P. (Eds.) Ostoje Ptaków o Znaczeniu Międzynarodowym w Polsce; OTOP: Marki, Poland, 2010. [Google Scholar]
- Dawidziuk, J.; Zajączkowski, S. Ochrona przyrody w planach urządzenia lasu w Lasach Państwowych. Zarządzanie Ochr. Przyr. Lasach 2011, 5, 228–244. [Google Scholar]
- Mizera, T. 20 lat funkcjonowania ochrony strefowej w Polsce. Stud. I Mater. Cent. Edukac. Przyr. Leśnej 2006, 8, 29–35. [Google Scholar]
- Kajtoch, Ł. Puszczyk uralski Strix uralensis. In Materiały do Wyznaczania i Określania Stanu Zachowania Siedlisk Ptasich w Obszarach Specjalnej Ochrony Ptaków Natura 2000 [Designation and quality evaluation of bird habitats in Natura 2000 Special Protection Areas]; Zawadzka, D., Ciach, M., Figarski, T., Kajtoch, Ł., Rejt, Ł., Eds.; GDOŚ: Warszawa, Poland, 2013; pp. 203–208. [Google Scholar]
- Morant, J.; Zabala, J.; Martínez, J.E.; Zuberogoitia, I. Out of sight, out of mind? Testing the effects of overwinter habitat alterations on breeding territories of a migratory endangered species. Anim. Conserv. 2018, 21, 465–473. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Period | Metric | Z | p |
|---|---|---|---|---|
| goshawk | wintering | Richness | 2.46 | 0.014 |
| Abundance | 2.50 | 0.012 | ||
| Natura | 1.33 | 0.184 | ||
| breeding | Richness | 2.47 | 0.014 | |
| Abundance | 3.28 | 0.001 | ||
| Natura | 3.64 | 0.000 | ||
| Ural owl | wintering | Richness | 2.41 | 0.016 |
| Abundance | 1.91 | 0.056 | ||
| Natura | 1.27 | 0.204 | ||
| breeding | Richness | 3.12 | 0.002 | |
| Abundance | 3.45 | 0.001 | ||
| Natura | 3.78 | 0.000 |
| Metric | ANOVA | p |
|---|---|---|
| wintering | ||
| Richness | 10.9 | 0.004 |
| Abundance | 15.5 | 0.004 |
| Natura | 18.4 | 0.000 |
| breeding | ||
| Richness | 16.2 | 0.000 |
| Abundance | 7.1 | 0.029 |
| Natura | 2.3 | 0.323 |
| Wald | p | Wald | p | Wald | p | |
|---|---|---|---|---|---|---|
| Diversity Metrics/Variables | Richness | Abundance | Natura | |||
| goshawk wintering | ||||||
| Intercept | 1089.8 | 0.00 | 6291.9 | 0.00 | 2.3 | 0.13 |
| Wood | 6.8 | 0.01 | 29.6 | 0.00 | 10.3 | 0.00 |
| OldTrees | 0.3 | 0.56 | 0.0 | 0.88 | 0.1 | 0.72 |
| DeadWood | 0.4 | 0.53 | 0.1 | 0.75 | 0.4 | 0.51 |
| Stumps | 0.1 | 0.73 | 0.2 | 0.68 | 0.3 | 0.56 |
| ClearCuts | 0.2 | 0.65 | 1.2 | 0.28 | 0.2 | 0.62 |
| goshawk | 3.3 | 0.07 | 61.9 | 0.00 | 1.3 | 0.26 |
| goshawk breeding | ||||||
| Intercept | 3042.3 | 0.00 | 10161.5 | 0.00 | 2.2 | 0.14 |
| Wood | 5.6 | 0.02 | 1.1 | 0.29 | 5.6 | 0.02 |
| OldTrees | 0.3 | 0.59 | 3.2 | 0.08 | 1.9 | 0.17 |
| DeadWood | 0.0 | 0.88 | 0.1 | 0.74 | 3.4 | 0.07 |
| Stumps | 0.0 | 0.95 | 0.0 | 0.86 | 0.6 | 0.42 |
| ClearCuts | 0.0 | 0.97 | 0.5 | 0.48 | 2.8 | 0.09 |
| goshawk | 3.7 | 0.05 | 11.0 | 0.00 | 17.1 | 0.00 |
| Ural owl wintering | ||||||
| Intercept | 10159.1 | 0.00 | 3.7 | 0.06 | 3.7 | 0.06 |
| Wood | 0.2 | 0.66 | 4.6 | 0.03 | 4.6 | 0.03 |
| OldTrees | 2.4 | 0.12 | 0.4 | 0.54 | 0.4 | 0.54 |
| DeadWood | 0.1 | 0.74 | 0.8 | 0.37 | 0.8 | 0.37 |
| Stumps | 0.2 | 0.69 | 0.4 | 0.54 | 0.4 | 0.54 |
| ClearCuts | 0.2 | 0.65 | 1.0 | 0.33 | 1.0 | 0.33 |
| Ural owl | 17.2 | 0.00 | 15.9 | 0.00 | 15.9 | 0.00 |
| Ural owl breeding | ||||||
| Intercept | 3020.2 | 0.00 | 10159.1 | 0.00 | 3.7 | 0.06 |
| Wood | 4.0 | 0.05 | 0.2 | 0.66 | 4.6 | 0.03 |
| OldTrees | 0.1 | 0.76 | 2.4 | 0.12 | 0.4 | 0.54 |
| DeadWood | 0.1 | 0.79 | 0.1 | 0.74 | 0.8 | 0.37 |
| Stumps | 0.1 | 0.74 | 0.2 | 0.69 | 0.4 | 0.54 |
| ClearCuts | 0.1 | 0.81 | 0.2 | 0.65 | 1.0 | 0.33 |
| Ural owl | 7.8 | 0.01 | 17.2 | 0.00 | 15.9 | 0.00 |
| Model | AIC | w |
|---|---|---|
| period: wintering explained metric: Richness | ||
| OldTrees + Stumps + DeadWood + goshawk + Ural owl + Wood | 302.4 | 0.2 |
| Stumps + goshawk + Ural owl + Wood | 302.6 | 0.2 |
| OldTrees + DeadWood + goshawk + Ural owl + Wood | 303.7 | 0.1 |
| OldTrees + Stumps + goshawk + Ural owl + Wood | 303.8 | 0.1 |
| goshawk + Ural owl + Wood | 303.9 | 0.1 |
| OldTrees + ClearCuts + DeadWood + goshawk + Ural owl + Wood | 304.2 | 0.1 |
| OldTrees + Stumps + ClearCuts + DeadWood + goshawk + Ural owl | 304.3 | 0.1 |
| Intercept | 383.7 | 0.0 |
| period: wintering metric: Abundance | ||
| Stumps + ClearCuts + goshawk + Ural + owl | 526.6 | 0.3 |
| Stumps + goshawk + Ural owl + Wood | 526.7 | 0.2 |
| OldTrees + Stumps + ClearCuts + goshawk + Ural owl + Wood | 527.7 | 0.1 |
| OldTrees + Stumps + goshawk + Ural owl + Wood | 527.8 | 0.1 |
| Stumps + DeadWood + goshawk + Ural owl + Wood | 528.3 | 0.1 |
| Stumps + ClearCuts + DeadWood + goshawk | 528.4 | 0.1 |
| Intercept | 677.8 | 0.0 |
| period: breeding metric: Richness | ||
| OldTrees + Ural owl + Wood | 296.1 | 0.3 |
| OldTrees + goshawk + Ural owl | 297.5 | 0.1 |
| OldTrees + ClearCuts + Ural owl | 297.8 | 0.1 |
| OldTrees + DeadWood + Ural owl | 297.8 | 0.1 |
| DeadWood + Ural owl + Wood | 298.0 | 0.1 |
| OldTrees + Stumps + Ural owl + Wood | 298.1 | 0.1 |
| Intercept | 319.7 | 0.0 |
| period: breeding metric: Abundance | ||
| OldTrees + Ural owl | 383.2 | 0.3 |
| OldTrees + goshawk + Ural owl | 383.7 | 0.2 |
| OldTrees + DeadWood + Ural owl | 384.2 | 0.2 |
| OldTrees + DeadWood + goshawk + Ural owl | 384.2 | 0.2 |
| OldTrees + Ural owl + Wood | 384.7 | 0.1 |
| OldTrees + goshawk + Ural owl + Wood | 384.8 | 0.1 |
| OldTrees + ClearCuts + Ural owl | 384.9 | 0.1 |
| OldTrees + Stumps + Ural owl | 385.1 | 0.1 |
| Intercept | 448.0 | 0.0 |
| period: wintering metric: Natura | ||
| goshawk + Wood | 118.0 | 0.3 |
| DeadWood + goshawk + Wood | 119.1 | 0.1 |
| goshawk + Ural owl + Wood | 119.3 | 0.1 |
| OldTrees + goshawk + Wood | 119.5 | 0.1 |
| Stumps + goshawk + Wood | 119.6 | 0.1 |
| DeadWood + Wood | 119.7 | 0.1 |
| Intercept | 135.9 | 0.0 |
| period: breeding metric: Natura | ||
| OldTrees + Stumps + ClearCuts + goshawk + Ural owl | 308.3 | 0.3 |
| OldTrees + Stumps + goshawk + Ural owl | 308.7 | 0.2 |
| OldTrees + Stumps + goshawk + Ural owl + Wood | 309.5 | 0.1 |
| OldTrees + Stumps + ClearCuts + goshawk + Ural owl + Wood | 309.9 | 0.1 |
| OldTrees + Stumps + ClearCuts + DeadWood + goshawk + Ural owl | 310.3 | 0.1 |
| Intercept | 1136.5 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kajtoch, Ł.; Kusal, B. The Long-Lasting Territories of Forest Apex Predators Sustain Diverse Bird Communities throughout the Year. Forests 2022, 13, 2128. https://doi.org/10.3390/f13122128
Kajtoch Ł, Kusal B. The Long-Lasting Territories of Forest Apex Predators Sustain Diverse Bird Communities throughout the Year. Forests. 2022; 13(12):2128. https://doi.org/10.3390/f13122128
Chicago/Turabian StyleKajtoch, Łukasz, and Bartłomiej Kusal. 2022. "The Long-Lasting Territories of Forest Apex Predators Sustain Diverse Bird Communities throughout the Year" Forests 13, no. 12: 2128. https://doi.org/10.3390/f13122128