
In Situ Rainwater Collection and Infiltration System Alleviates the Negative Effects of Drought on Plant-Available Water, Fine Root Distribution and Plant Hydraulic Conductivity
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Plot
2.3. Fine-Root Measurement
2.4. Soil Water Content and Rainfall Measurement
2.5. The Percent Loss of Hydraulic Conductivity (PLC)
2.6. Statistical Analysis
3. Results
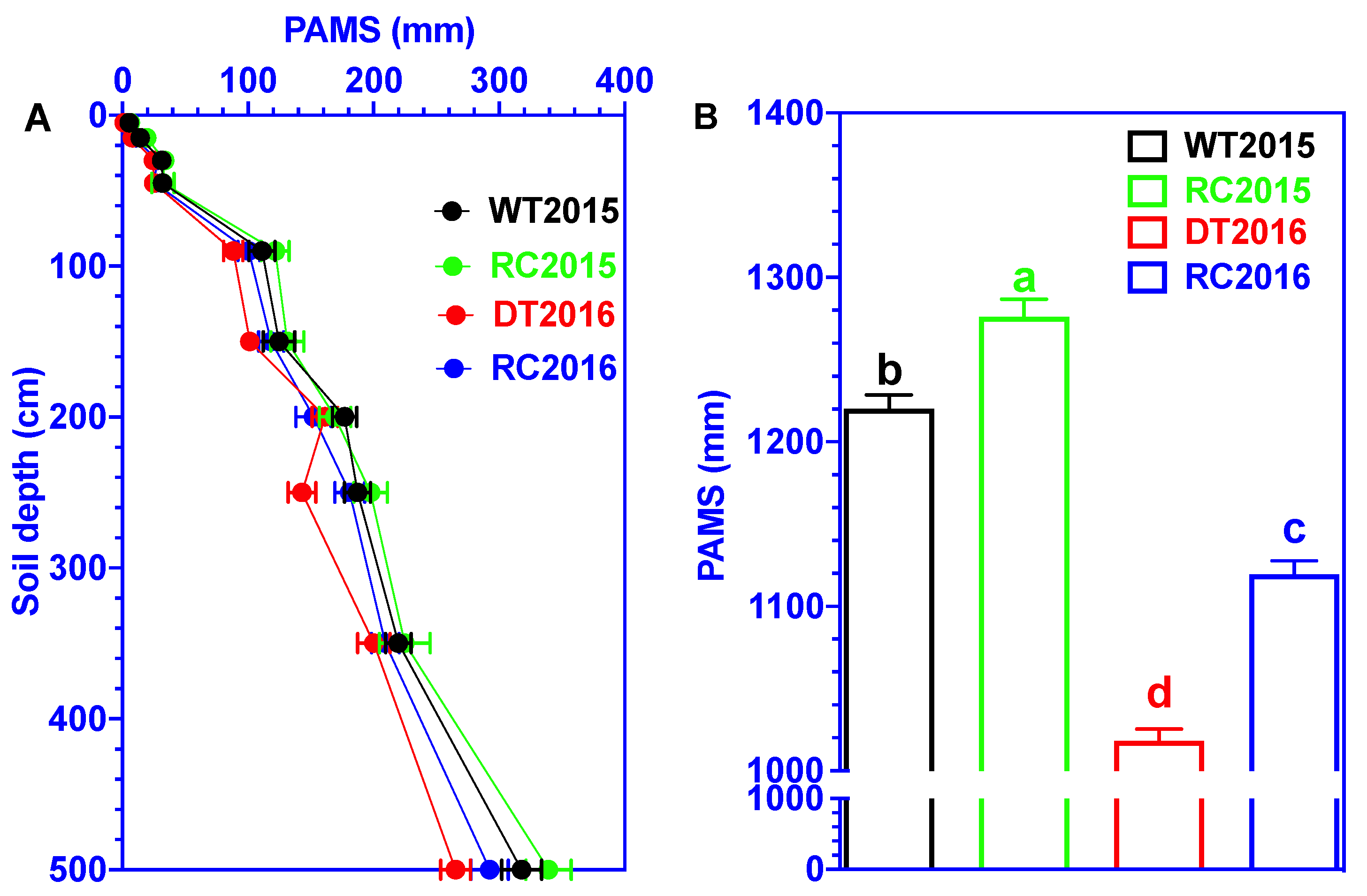
3.1. Plant-Available Moisture Storage (PAMS)
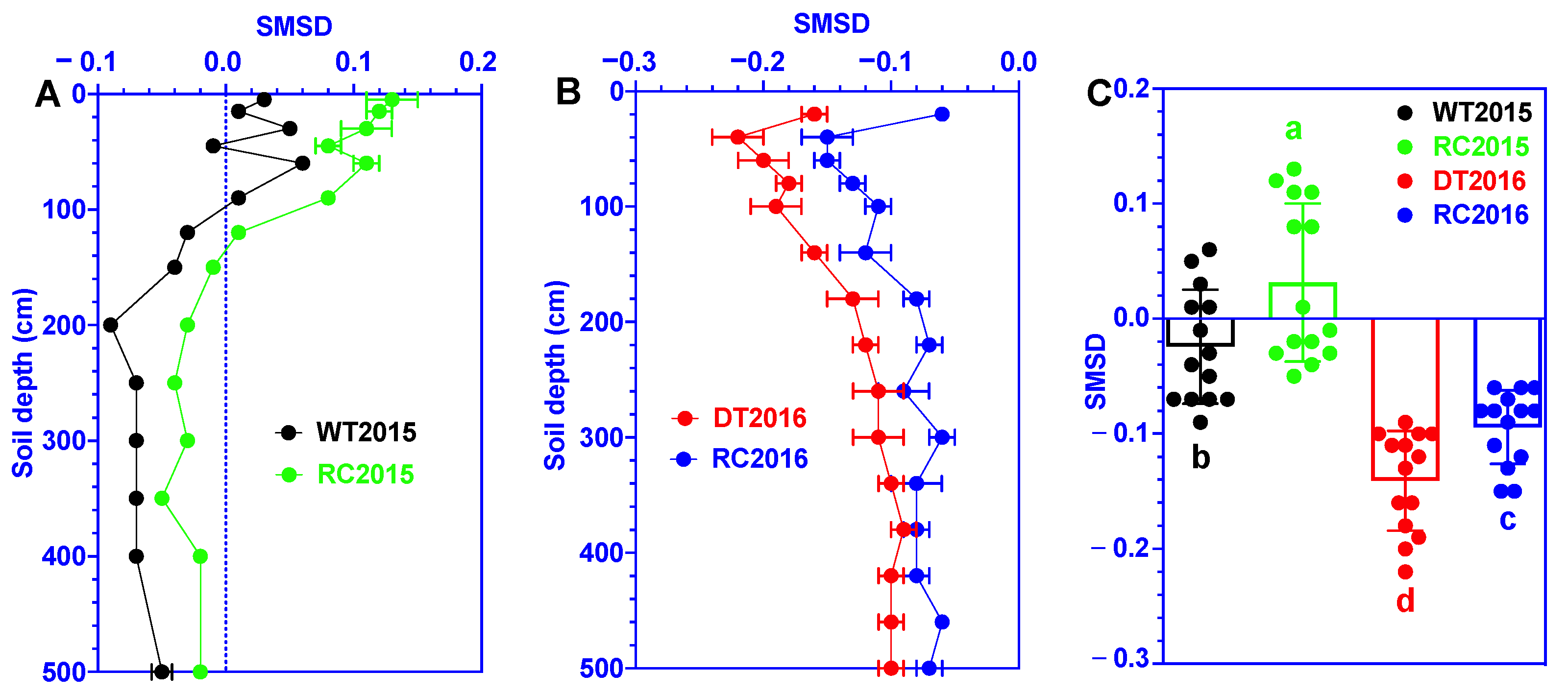
3.2. Soil Moisture Storage Deficit (SMSD)
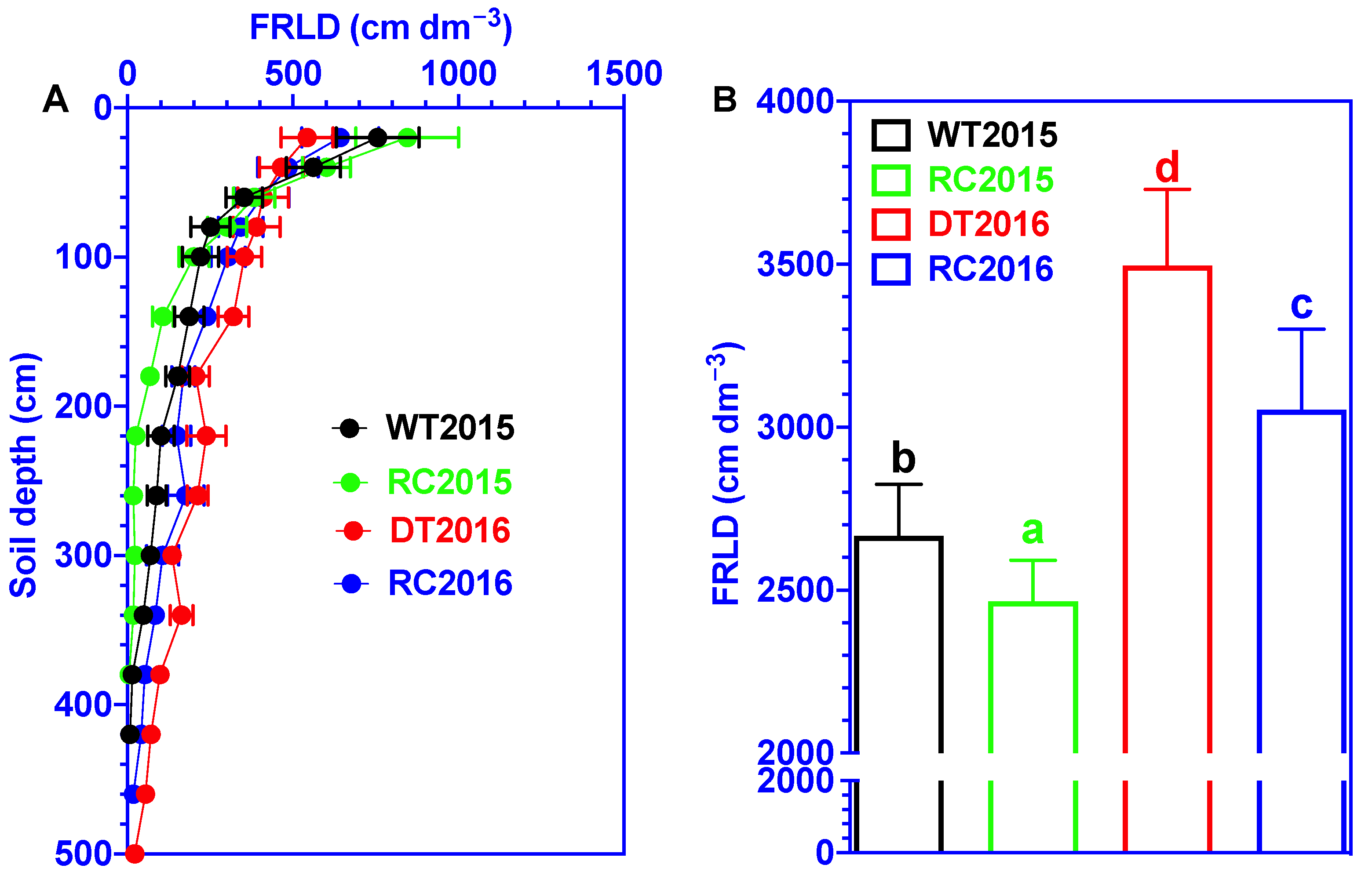
3.3. Fine-Root Distribution (FRLD)
3.4. The Percentage Loss of Hydraulic Conductivity (PLC)
4. Discussion
4.1. Soil Moisture Profile
4.2. Root Distribution Pattern
4.3. PLC
4.4. Implications for Afforest Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.-S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Cheng, X.; Huang, M.; Shao, M.; Warrington, D.N. A comparison of fine root distribution and water consumption of mature Caragana korshinkii Kom grown in two soils in a semiarid region, China. Plant Soil 2009, 315, 149–161. [Google Scholar] [CrossRef]
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial water fluxes dominated by transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. The global biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Gao, X.; Li, H.; Zhao, X.; Ma, W.; Wu, P. Identifying a suitable revegetation technique for soil restoration on water-limited and degraded land: Considering both deep soil moisture deficit and soil organic carbon sequestration. Geoderma 2018, 319, 61–69. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Song, X.; Gao, X.; Wu, P.; Zhao, X.; Zhang, W.; Zou, Y.; Siddique, K.H.M. Drought responses of profile plant-available water and fine-root distributions in apple (Malus pumila Mill.) orchards in a loessial, semi-arid, hilly area of China. Sci. Total Environ. 2020, 723, 137739. [Google Scholar] [CrossRef]
- Leonova, A.; Heger, A.; Vásconez Navas, L.K.; Jensen, K.; Reisdorff, C. Fine root mortality under severe drought reflects different root distribution of Quercus robur and Ulmus laevis trees in hardwood floodplain forests. Trees 2022, 36, 1105–1115. [Google Scholar] [CrossRef]
- Zwetsloot, M.J.; Bauerle, T.L. Repetitive seasonal drought causes substantial species-specific shifts in fine-root longevity and spatio-temporal production patterns in mature temperate forest trees. New Phytol. 2021, 231, 974–986. [Google Scholar] [CrossRef]
- Nikolova, P.S.; Bauerle, T.L.; Haberle, K.H.; Blaschke, H.; Brunner, I.; Matyssek, R. Fine-root traits reveal contrasting ecological strategies in european beech and norway spruce during extreme drought. Front. Plant Sci. 2020, 11, 1211. [Google Scholar] [CrossRef]
- Germon, A.; Jourdan, C.; Bordron, B.; Robin, A.; Nouvellon, Y.; Chapuis-Lardy, L.; de Moraes Gonçalves, J.L.; Pradier, C.; Guerrini, I.A.; Laclau, J.-P. Consequences of clear-cutting and drought on fine root dynamics down to 17 m in coppice-managed eucalypt plantations. For. Ecol. Manag. 2019, 445, 48–59. [Google Scholar] [CrossRef]
- Fuchs, S.; Hertel, D.; Schuldt, B.; Leuschner, C. Effects of summer drought on the fine root system of five broadleaf tree species along a precipitation gradient. Forests 2020, 11, 289. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Gao, X.; Zhao, X.; Wu, P.; Dyck, M. Spatial distribution of soil moisture and fine roots in rain-fed apple orchards employing a Rainwater Collection and Infiltration (RWCI) system on the Loess Plateau of China. Agric. Water Manag. 2017, 184, 170–177. [Google Scholar] [CrossRef]
- Ma, C.; Luo, Y.; Shao, M.; Li, X.; Sun, L.; Jia, X. Environmental controls on sap flow in black locust forest in Loess Plateau, China. Sci. Rep. 2017, 7, 13160. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Zhu, Y.; Luo, Y. Soil moisture decline due to afforestation across the Loess Plateau, China. J. Hydrol. 2017, 546, 113–122. [Google Scholar] [CrossRef]
- Böhm, W. Methods of Studying Root Systems; Springer: Berlin/Heidelberg, Germany, 1979. [Google Scholar]
- Vijaya Kumar, P.; Bindi, M.; Crisci, A.; Maracchi, G. Detection of variations in precipitation at different time scales of twentieth century at three locations of Italy. Weather Clim. Extrem. 2013, 2, 7–15. [Google Scholar] [CrossRef]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ. 1988, 11, 35–40. [Google Scholar] [CrossRef]
- Wang, Y.; Brandt, M.; Zhao, M.; Tong, X.; Xing, K.; Xue, F.; Kang, M.; Wang, L.; Jiang, Y.; Fensholt, R. Major forest increase on the Loess Plateau, China (2001–2016). Land Degrad. Dev. 2018, 29, 4080–4091. [Google Scholar] [CrossRef]
- Ma, C.; Luo, Y.; Shao, M. Comparative modeling of the effect of thinning on canopy interception loss in a semiarid black locust (Robinia pseudoacacia) plantation in Northwest China. J. Hydrol. 2020, 590, 125234. [Google Scholar] [CrossRef]
- Ma, L.H.; Liu, X.L.; Wang, Y.K.; Wu, P.T. Effects of drip irrigation on deep root distribution, rooting depth, and soil water profile of jujube in a semiarid region. Plant Soil 2013, 373, 995–1006. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How tree roots respond to drought. Front. Plant Sci. 2015, 6, 547. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.Q.; Wang, J.M.; Wang, W.J.; Zhang, T.H.; Li, J.W. Effects of root phenotypic changes on the deep rooting of Populus euphratica seedlings under drought stresses. PeerJ 2019, 7, e6513. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Luo, Y.; Sun, L.; Li, X.; Ma, C.; Wang, X.; Jiang, T.; Zhu, H. Modelling the artificial forest (Robinia pseudoacacia L.) root–soil water interactions in the Loess Plateau, China. Hydrol. Earth Syst. Sci. 2022, 26, 17–34. [Google Scholar] [CrossRef]
- Ma, L.H.; Wu, P.T.; Wang, Y.K. Spatial distribution of roots in a dense jujube plantation in the semiarid hilly region of the Chinese Loess Plateau. Plant Soil 2011, 354, 57–68. [Google Scholar] [CrossRef]
- Kleidon, A.; Heiman, M. A method of determining rooting depth from a terrestrial biosphere model and its impacts on the global water and carbon cycle. Glob. Change Biol. 1998, 4, 275–286. [Google Scholar] [CrossRef]
- Burgess, S.S.O.; Adams, M.A.; Turner, N.C.; White, D.A.; Ong, C.K. Tree roots: Conduits for deep recharge of soil water. Oecologia 2001, 126, 158–165. [Google Scholar] [CrossRef]
- Anderegg, W.R.; Klein, T.; Bartlett, M.; Sack, L.; Pellegrini, A.F.; Choat, B.; Jansen, S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proc. Natl. Acad. Sci. USA 2016, 113, 5024–5029. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Gao, X.; Dyck, M.; Zhang, W.; Wu, P.; Yao, J.; Zhao, X. Soil water and root distribution of apple tree (Malus pumila Mill) stands in relation to stand age and rainwater collection and infiltration system (RWCI) in a hilly region of the Loess Plateau, China. Catena 2018, 170, 324–334. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Meng, H.; Xie, B.; Li, Q.; Li, X.; Zhou, B.; Wang, Q.; Luo, Y. In Situ Rainwater Collection and Infiltration System Alleviates the Negative Effects of Drought on Plant-Available Water, Fine Root Distribution and Plant Hydraulic Conductivity. Forests 2022, 13, 2082. https://doi.org/10.3390/f13122082
Ma C, Meng H, Xie B, Li Q, Li X, Zhou B, Wang Q, Luo Y. In Situ Rainwater Collection and Infiltration System Alleviates the Negative Effects of Drought on Plant-Available Water, Fine Root Distribution and Plant Hydraulic Conductivity. Forests. 2022; 13(12):2082. https://doi.org/10.3390/f13122082
Chicago/Turabian StyleMa, Changkun, Haobo Meng, Biao Xie, Qian Li, Xiangdong Li, Beibei Zhou, Quanjiu Wang, and Yi Luo. 2022. "In Situ Rainwater Collection and Infiltration System Alleviates the Negative Effects of Drought on Plant-Available Water, Fine Root Distribution and Plant Hydraulic Conductivity" Forests 13, no. 12: 2082. https://doi.org/10.3390/f13122082