
Contrasting Soil Microbial Functional Potential for Phosphorus Cycling in Subtropical and Temperate Forests

Abstract
:1. Introduction
2. Material and Methods
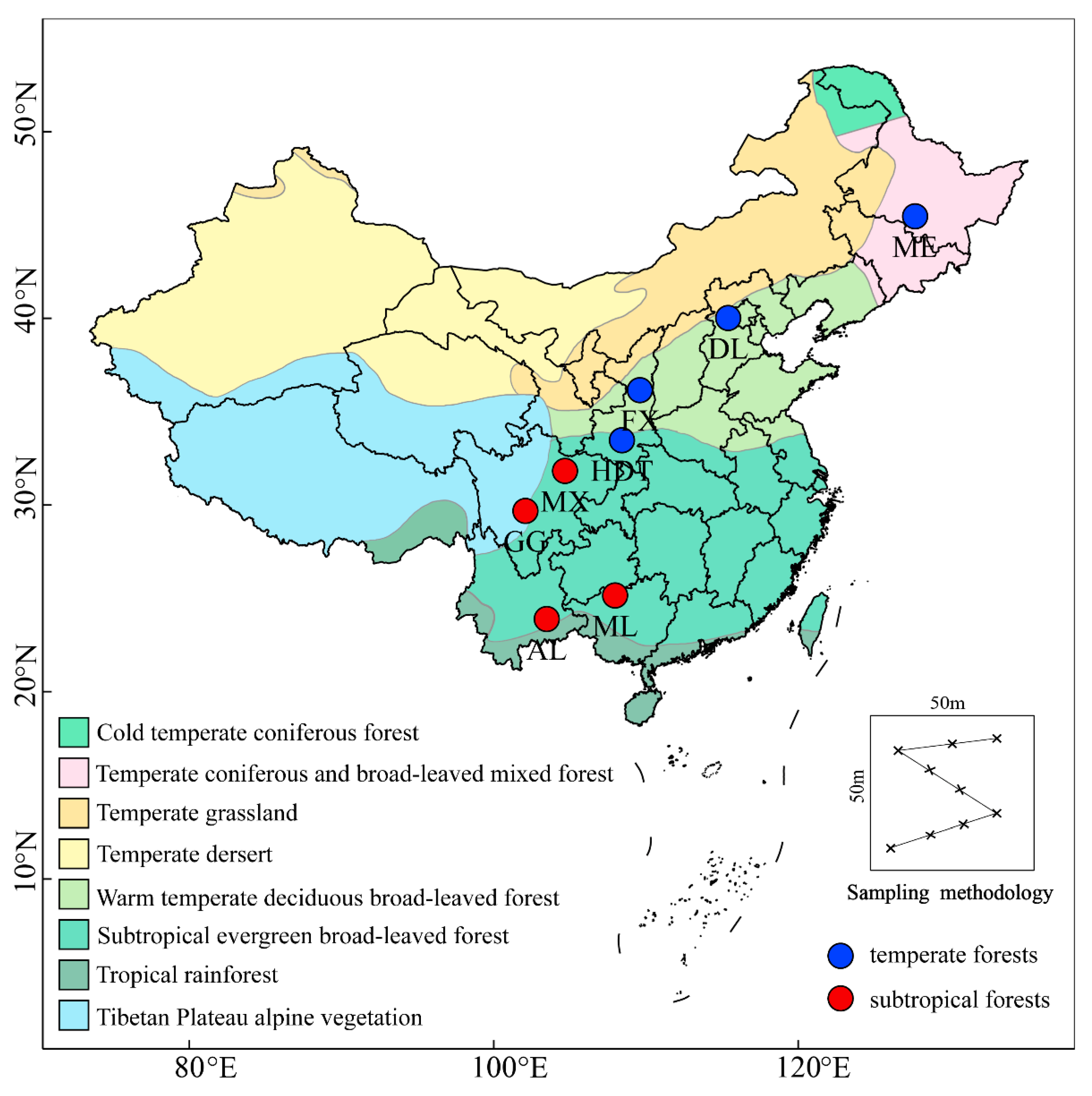
2.1. Sites and Soil Sampling
2.2. Soil Physicochemical Analyses
2.3. Microbial Community Analysis
2.4. Metagenomics Analysis
2.5. Statistical Analysis
3. Results
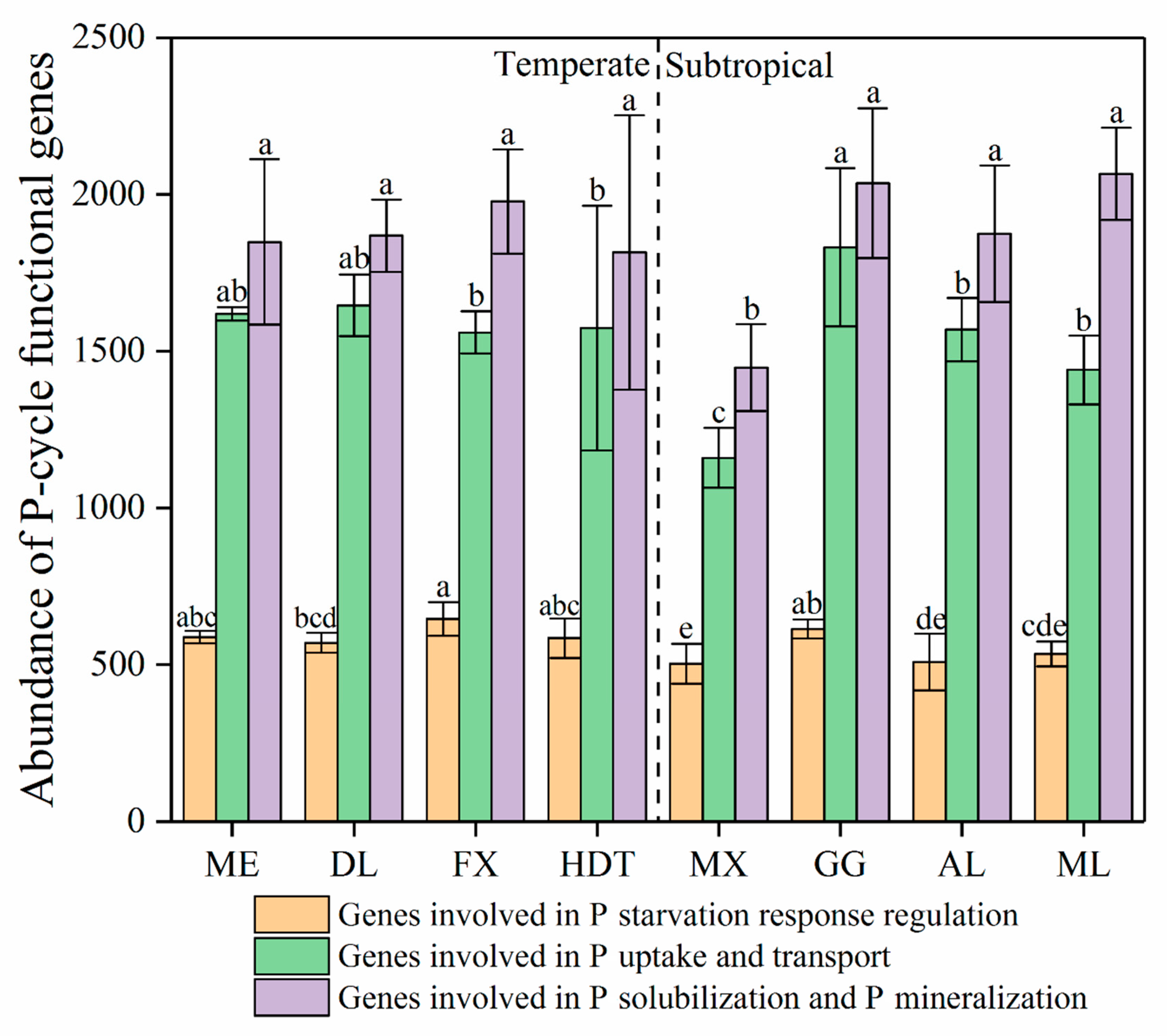
3.1. P Functional Genes in Temperate and Subtropical Zones
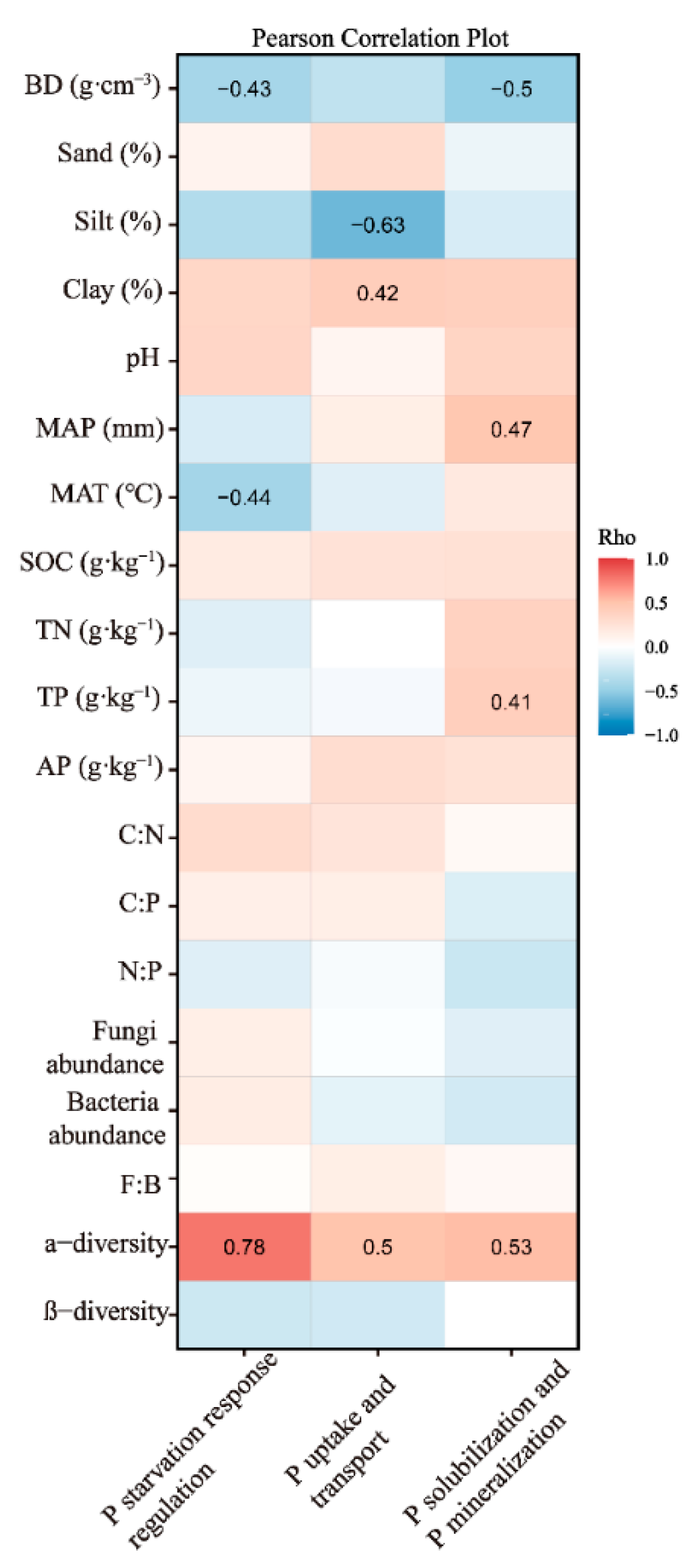
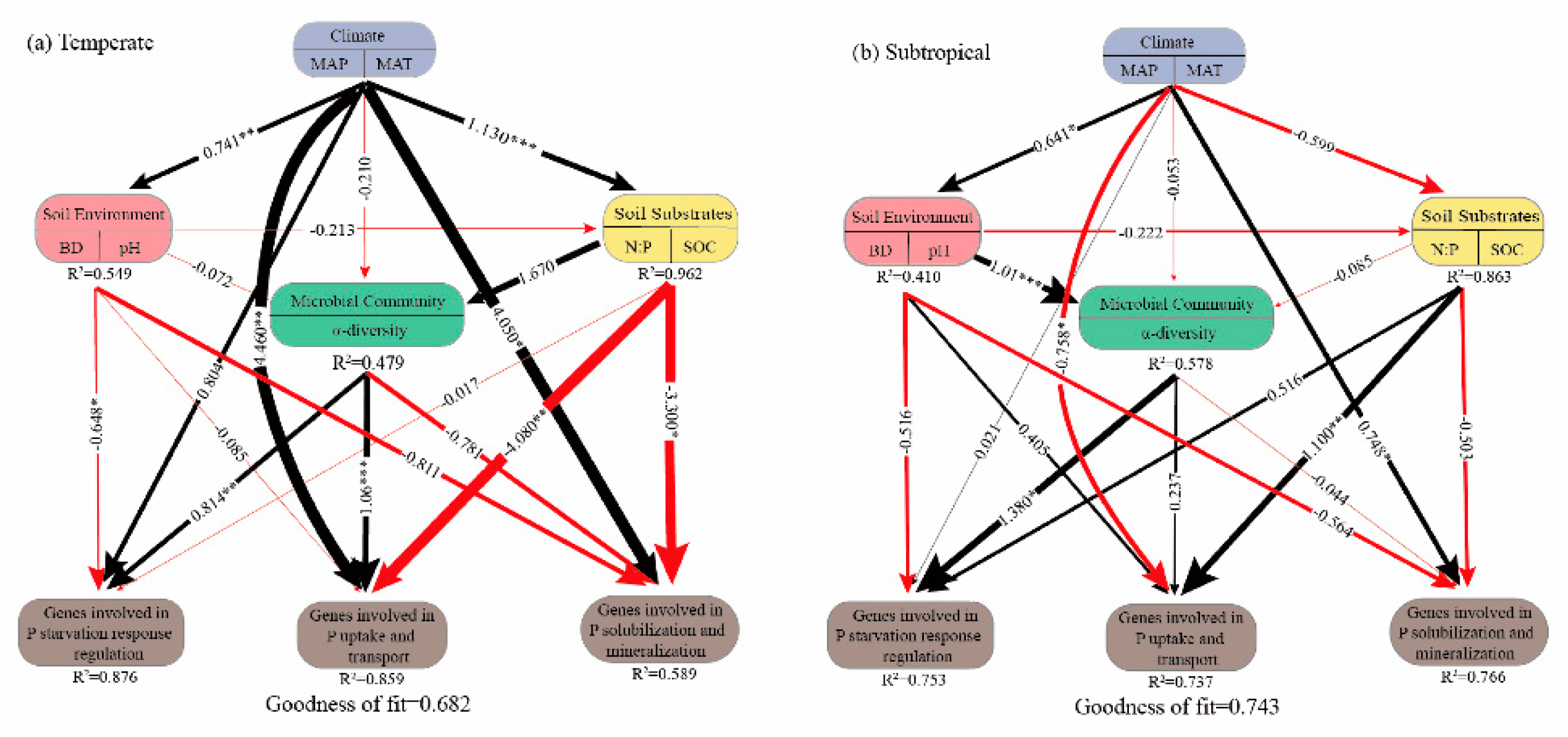
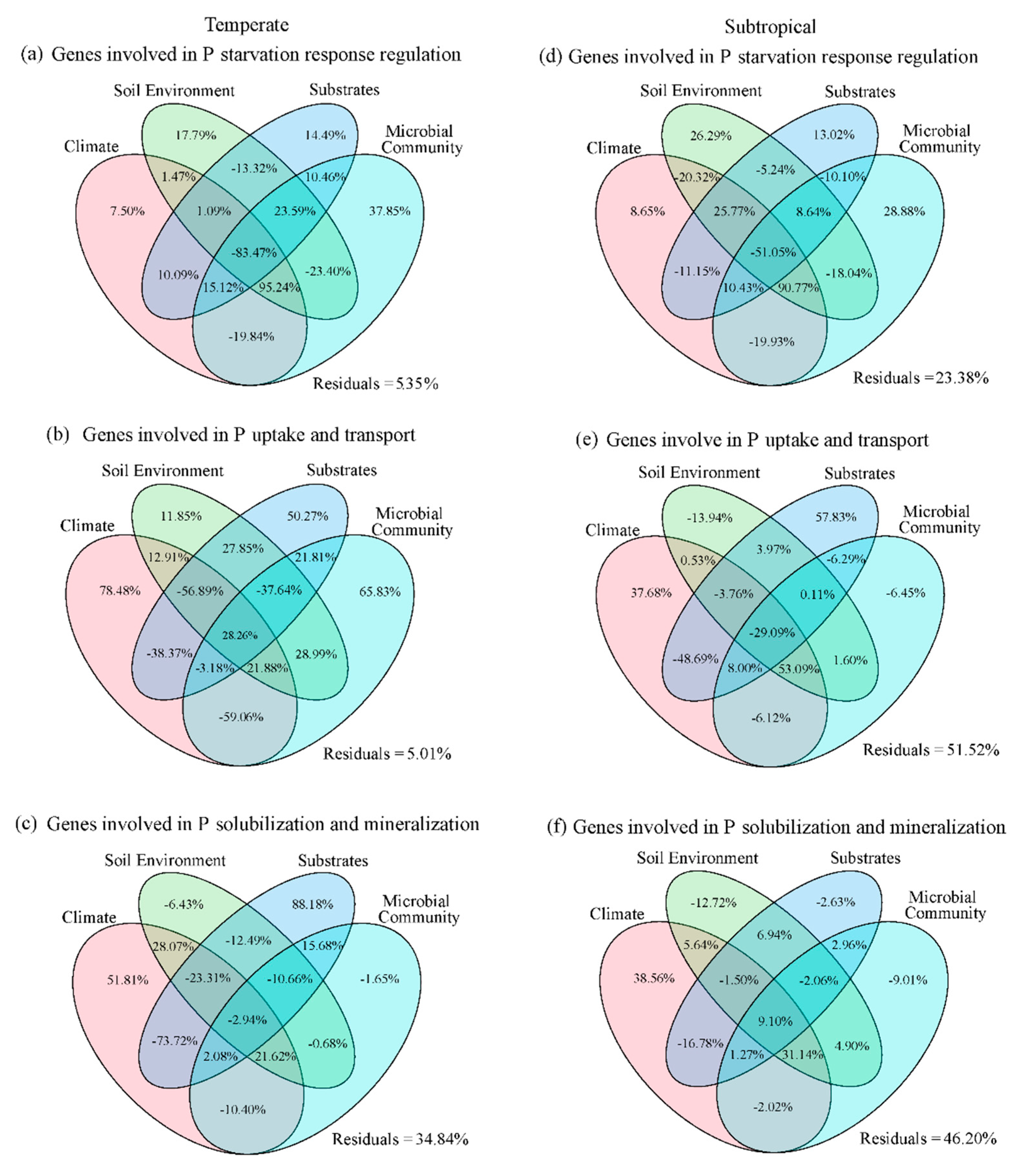
3.2. Factors Influencing P Functional Genes in the Temperate and Subtropical Zones
4. Discussion
4.1. P Functional Genes in Temperate and Subtropical Zones
4.2. Factors Affecting Soil P Functional Genes in Temperate and Subtropical Zones
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pastore, G.; Kernchen, S.; Spohn, M. Microbial solubilization of silicon and phosphorus from bedrock in relation to abundance of phosphorus-solubilizing bacteria in temperate forest soils. Soil Biol. Biochem. 2020, 151, 108050. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Guo, Y.P.; Tang, Z.Y.; Feng, Y.H.; Zhu, X.R.; Xu, W.T.; Bai, Y.F.; Zhou, G.Y.; Xie, Z.Q.; Fang, J.Y. Patterns of nitrogen and phosphorus pools in terrestrial ecosystems in China. Earth Syst. Sci. Data 2021, 13, 5337–5351. [Google Scholar] [CrossRef]
- Powers, S.; Mirsky, E.; Bandaranayake, A.; Thavarajah, P.; Shipe, E.; Bridges, W.; Thavarajah, D. Field pea (Pisum sativum L.) shows genetic variation in phosphorus use efficiency in different P environments. Sci. Rep. 2020, 10, 18940. [Google Scholar] [CrossRef]
- Tian, J.; Dungait, J.A.J.; Lu, X.K.; Yang, Y.F.; Hartley, I.P.; Zhang, W.; Mo, J.M.; Yu, G.R.; Zhou, J.Z.; Kuzyakov, Y. Long-term nitrogen addition modifies microbial composition and functions for slow carbon cycling and increased sequestration in tropical forest soil. Glob. Change Biol. 2019, 25, 3267–3281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavka, M.; Majcherczyk, A.; Kues, U.; Polle, A. Phylogeny, tissue-specific expression, and activities of root-secreted purple acid phosphatases for P uptake from ATP in P starved poplar. Plant Sci. 2021, 307, 110906. [Google Scholar] [CrossRef] [PubMed]
- Bechtaoui, N.; Rabiu, M.K.; Raklami, A.; Oufdou, K.; Hafidi, M.; Jemo, M. Phosphate-dependent regulation of growth and stresses management in plants. Front. Plant Sci. 2021, 12, 679916. [Google Scholar] [CrossRef]
- Fleischer, K.; Rammig, A.; De Kauwe, M.G.; Walker, A.P.; Domingues, T.F.; Fuchslueger, L.; Garcia, S.; Goll, D.S.; Grandis, A.; Jiang, M.K.; et al. Amazon forest response to CO2 fertilization dependent on plant phosphorus acquisition. Nat. Geosci. 2019, 12, 736–741. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.M.; Liu, G.F.; Chen, H.H.; Chen, C.R.; Wang, J.K.; Ai, S.Y.; Wei, D.; Li, D.M.; Ma, B.; Tang, C.X.; et al. Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. Isme J. 2020, 14, 757–770. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Li, X.Z.; Datta, R.H.; Chen, J.; Du, Y.Z.; Du, D.L. Key factors shaping prokaryotic communities in subtropical forest soils. Appl. Soil Ecol. 2022, 169, 104162. [Google Scholar] [CrossRef]
- Bergkemper, F.; Scholer, A.; Engel, M.; Lang, F.; Kruger, J.; Schloter, M.; Schulz, S. Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 2016, 18, 1988–2000. [Google Scholar] [CrossRef]
- Li, M.; Hao, Y.B.; Yan, Z.Q.; Kang, E.Z.; Wang, J.Z.; Zhang, K.R.; Li, Y.; Wu, H.D.; Kang, X.M. Long-term degradation from marshes into meadows shifts microbial functional diversity of soil phosphorus cycling in an alpine wetland of the Tibetan Plateau. Land Degrad. Dev. 2022, 33, 628–637. [Google Scholar] [CrossRef]
- Oliverio, A.M.; Bissett, A.; McGuire, K.; Saltonstall, K.; Turner, B.L.; Fierer, N. The role of phosphorus limitation in shaping soil bacterial communities and their metabolic capabilities. Mbio 2020, 11, e01718-20. [Google Scholar] [CrossRef]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-solubilizing microorganisms: Mechanism and their role in phosphate solubilizing and uptake. J. Soil Sci. Plant Nutr. 2021, 21, 49–68. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Park, Y.S.; Kim, S.J.; Lee, B.J.; Suh, S.W. Crystal structure of PhoU from Pseudomonas aeruginosa, a negative regulator of the Pho regulon. J. Struct. Biol. 2014, 188, 22–29. [Google Scholar] [CrossRef]
- Wu, X.J.; Rensing, C.; Han, D.F.; Xiao, K.Q.; Dai, Y.X.; Tang, Z.X.; Liesack, W.; Peng, J.J.; Cui, Z.L.; Zhang, F.S. Genome-Resolved Metagenomics Reveals Distinct Phosphorus Acquisition Strategies between Soil Microbiomes. Msystems 2022, 7, e01107-21. [Google Scholar] [CrossRef]
- Wei, X.M.; Hu, Y.J.; Cai, G.; Yao, H.Y.; Ye, J.; Sun, Q.; Veresoglou, S.D.; Li, Y.Y.; Zhu, Z.K.; Guggenberger, G.; et al. Organic phosphorus availability shapes the diversity of phoD-harboring bacteria in agricultural soil. Soil Biol. Biochem. 2021, 161, 108364. [Google Scholar] [CrossRef]
- Siegenthaler, M.B.; Ramoneda, J.; Frossard, E.; Meszaros, E. Microbial community responses to phosphorus and nitrogen inputs in the organic soil horizons of two contrasting temperate beech forests. Appl. Soil Ecol. 2022, 172, 104357. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.L.; Liu, J.; Jia, P.; Yang, T.T.; Zeng, Q.W.; Zhang, S.C.; Liao, B.; Shu, W.S.; Li, J.T. Novel phosphate-solubilizing bacteria enhance soil phosphorus cycling following ecological restoration of land degraded by mining. Isme J. 2020, 14, 1600–1613. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Sun, X.T.; Sun, F.W.; Zhao, Y.A.; Sun, W.; Guo, J.X.; Zhang, T. Sensitivity of soil fungal and bacterial community compositions to nitrogen and phosphorus additions in a temperate meadow. Plant Soil 2022, 471, 477–490. [Google Scholar] [CrossRef]
- Li, J.B.; Xie, T.; Zhu, H.; Zhou, J.; Li, C.N.; Xiong, W.J.; Xu, L.; Wu, Y.H.; He, Z.L.; Li, X.Z. Alkaline phosphatase activity mediates soil organic phosphorus mineralization in a subalpine forest ecosystem. Geoderma 2021, 404, 115376. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, Y.; Xia, Y.; Cui, Z.B.; Cao, C.Y. Soil microbial community succession based on PhoD and Gcd genes along a chronosequence of sand-fixation forest. Forests 2021, 12, 1707. [Google Scholar] [CrossRef]
- LeBrun, E.S.; King, R.S.; Back, J.A.; Kang, S. A metagenome-based investigation of gene relationships for non-substrate-associated microbial phosphorus cycling in the water column of streams and rivers. Microb. Ecol. 2018, 76, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Stirling, E.; Liu, Y.H.; Zhao, K.K.; Zhou, J.Z.; Singh, B.K.; Tang, C.X.; Dahlgren, R.A.; Xu, J.M. Soil biogeochemical cycle couplings inferred from a function-taxon network. Research 2021, 7102769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xue, S.; Liu, G.B.; Song, Z.L. A comparison of soil qualities of different revegetation types in the Loess Plateau, China. Plant Soil 2011, 347, 163–178. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Ren, C.J.; Zhang, X.Y.; Zhang, S.H.; Wang, J.Y.; Xu, M.P.; Guo, Y.X.; Wang, J.; Han, X.H.; Zhao, F.Z.; Yang, G.H.; et al. Altered microbial CAZyme families indicated dead biomass decomposition following afforestation. Soil Biol. Biochem. 2021, 160, 108362. [Google Scholar] [CrossRef]
- Ren, C.J.; Chen, J.; Deng, J.; Zhao, F.Z.; Han, X.H.; Yang, G.H.; Tong, X.G.; Feng, Y.Z.; Shelton, S.; Ren, G.X. Response of microbial diversity to C:N:P stoichiometry in fine root and microbial biomass following afforestation. Biol. Fertil. Soils 2017, 53, 457–468. [Google Scholar] [CrossRef]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.J.; Zhou, Z.H.; Guo, Y.X.; Yang, G.H.; Zhao, F.Z.; Wei, G.H.; Han, X.H.; Feng, L.; Feng, Y.Z.; Ren, G.X. Contrasting patterns of microbial community and enzyme activity between rhizosphere and bulk soil along an elevation gradient. Catena 2021, 196, 104921. [Google Scholar] [CrossRef]
- Zhang, X.M.; Johnston, E.R.; Barberan, A.; Ren, Y.; Lu, X.T.; Han, X.G. Decreased plant productivity resulting from plant group removal experiment constrains soil microbial functional diversity. Glob. Change Biol. 2017, 23, 4318–4332. [Google Scholar] [CrossRef] [PubMed]
- Li, D.H.; Liu, C.M.; Luo, R.B.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [Green Version]
- Henseler, J.; Hubona, G.; Ray, P.A. Using PLS path modeling in new technology research: Updated guidelines. Ind. Manag. Data Syst. 2016, 116, 2–20. [Google Scholar] [CrossRef]
- Cheng, J.M.; Han, Z.J.; Cong, J.; Yu, J.J.; Zhou, J.Z.; Zhao, M.X.; Zhang, Y.G. Edaphic variables are better indicators of soil microbial functional structure than plant-related ones in subtropical broad-leaved forests. Sci. Total Environ. 2021, 773, 145630. [Google Scholar] [CrossRef]
- Chu, H.Y.; Neufeld, J.D.; Walker, V.K.; Grogan, P. The influence of vegetation type on the dominant soil bacteria, archaea, and fungi in a low arctic tundra landscape. Soil Sci. Soc. Am. J. 2011, 75, 1756–1765. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.G.; Zhang, Z.S.; Huang, L.; Qi, Q.; Liu, L.C.; Zhao, Y.; Wang, Z.R.; Zhou, H.K.; Lv, X.Y.; Mao, Z.C.; et al. Shifts in soil microbial community functional gene structure across a 61-year desert revegetation chronosequence. Geoderma 2019, 347, 126–134. [Google Scholar] [CrossRef]
- Prathap, V.; Kumar, A.; Maheshwari, C.; Tyagi, A. Phosphorus homeostasis: Acquisition, sensing, and long-distance signaling in plants. Mol. Biol. Rep. 2022, 49, 8071–8086. [Google Scholar] [CrossRef] [PubMed]
- Maranguit, D.; Guillaume, T.; Kuzyakov, Y. Effects of flooding on phosphorus and iron mobilization in highly weathered soils under different land-use types: Short-term effects and mechanisms. Catena 2017, 158, 161–170. [Google Scholar] [CrossRef]
- Billah, M.; Khan, M.; Bano, A.; Ul Hassan, T.; Munir, A.; Gurmani, A.R. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Mason, L.M.; Eagar, A.; Patel, P.; Blackwood, C.B.; DeForest, J.L. Potential microbial bioindicators of phosphorus mining in a temperate deciduous forest. J. Appl. Microbiol. 2021, 130, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Gai, X.; Li, S.C.; Zhang, X.P.; Bian, F.Y.; Yang, C.A.B.; Zhong, Z.K. Changes in soil phosphorus availability and associated microbial properties after chicken farming in Lei bamboo (Phyllostachys praecox) forest ecosystems. Land Degrad. Dev. 2021, 32, 3008–3022. [Google Scholar] [CrossRef]
- Yi, X.Y.; Ning, C.; Feng, S.L.; Gao, H.Q.; Zhao, J.L.; Liao, J.Y.; Peng, Y.H.; Zhao, S.Q.; Liu, S.G. Urbanization-induced environmental changes strongly affect wetland soil bacterial community composition and diversity. Environ. Res. Lett. 2022, 17, 014027. [Google Scholar] [CrossRef]
- Louis, B.P.; Maron, P.A.; Menasseri-Aubry, S.; Sarr, A.; Leveque, J.; Mathieu, O.; Jolivet, C.; Leterme, P.; Viaud, V. Microbial diversity indexes can explain soil carbon dynamics as a function of carbon source. PLoS ONE 2016, 11, e0161251. [Google Scholar] [CrossRef] [Green Version]
- Custer, G.F.; Diepen, L. Plant invasion has limited impact on ssoil microbial α-diversity: A meta-analysis. Diversity 2020, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.G.; Wang, J.W.; Zhou, G.Y.; Bai, S.H.; Zhou, L.Y.; Fu, Y.L.; Wang, C.K.; Wang, H.M.; Yu, G.R.; Zhou, X.H. Differential effects of nitrogen vs. phosphorus limitation on terrestrial carbon storage in two subtropical forests: A bayesian approach. Sci. Total Environ. 2021, 795, 148485. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rodriguez, J.V.; Salazar-Vidal, M.N.; Chavez Montes, R.A.; Massange-Sanchez, J.A.; Gillmor, C.S.; Sawers, R.J.H. Low nitrogen availability inhibits the phosphorus starvation response in maize (Zea mays ssp. mays L.). BMC Plant Biol. 2021, 21, 259. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.L.; Lambers, H.; Condron, L.M.; Cramer, M.D.; Leake, J.R.; Richardson, A.E.; Smith, S.E. Soil microbial biomass and the fate of phosphorus during long-term ecosystem development. Plant Soil 2013, 367, 225–234. [Google Scholar] [CrossRef]
- Zhou, R.J.; Hou, D.W.; Zeng, S.Z.; Wei, D.D.; Yu, L.F.; Bao, S.C.; Weng, S.P.; He, J.G.; Huang, Z.J. Sedimentary nitrogen and sulfur reduction functional-couplings interplay with the microbial community of anthropogenic shrimp culture pond ecosystem. Front. Microbiol. 2022, 13, 830777. [Google Scholar] [CrossRef]
- Wu, J.W.; Liang, J.L.; Bjorn, L.O.; Li, J.T.; Shu, W.S.; Wang, Y.T. Phosphorus-arsenic interaction in the ‘soil-plant-microbe’ system and its influence on arsenic pollution. Sci. Total Environ. 2022, 802, 149796. [Google Scholar] [CrossRef]
- Bledsoe, R.B.; Goodwillie, C.; Peralta, A.L. Long-term nutrient enrichment of an oligotroph-dominated wetland increases bacterial diversity in bulk soils and plant rhizospheres. Msphere 2020, 5, e00035-20. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, L.; Wu, R.N.; Meng, H.; Li, X.Y.; Gu, J.D. Seasonal and spatial variations in diversity and abundance of bacterial laccase-like genes in sediments of a subtropical mangrove ecosystem. Int. Biodeterior. Biodegrad. 2016, 114, 260–267. [Google Scholar] [CrossRef]
- Mise, K.; Koyama, Y.; Matsumoto, A.; Fujita, K.; Kunito, T.; Senoo, K.; Otsuka, S. Pectin drives microbial phosphorus solubilization in soil: Evidence from isolation-based and community-scale approaches. Eur. J. Soil Biol. 2020, 97, 103169. [Google Scholar] [CrossRef]
- Hacket-Pain, A.J.; Ascoli, D.; Vacchiano, G.; Biondi, F.; Cavin, L.; Conedera, M.; Drobyshev, I.; Linan, I.D.; Friend, A.D.; Grabner, M.; et al. Climatically controlled reproduction drives interannual growth variability in a temperate tree species. Ecol. Lett. 2018, 21, 1833–1844. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phyla | Temperate Forest Biomes | Subtropical Forest Biomes | ||||
|---|---|---|---|---|---|---|
| P Starvation Response Regulation | P Uptake and Transport | P Solubilization and Mineralization | P Starvation Response Regulation | P Uptake and Transport | P Solubilization and Mineralization | |
| Acidobacteria | 312.08 ± 132.98 c | 981.67 ± 378.20 b | 2731.58 ± 298.35 a* | 287.42 ± 45.53 C | 1261.08 ± 131.38 B | 3768.83 ± 251.88 A* |
| Actinobacteria | 182.08 ± 14.06 c | 954.75 ± 42.00 a | 247.92 ± 27.48 b | 127.75 ± 35.32 B | 776.75 ± 119.54 A | 228.00 ± 43.39 B |
| Proteobacteria | 590.08 ± 40.31 c* | 3586.58 ± 261.20 a | 1403.83 ± 109.03 b | 769.00 ± 90.93 C* | 4856.33 ± 838.79 A | 2146.00 ± 568.13 B |
| Verrucomicrobia | 167.25 ± 63.96 b | 694.08 ± 221.41 a | 188.50 ± 70.74 b | 112.17 ± 15.15 B | 425.08 ± 50.20 A | 121.33 ± 25.27 B |
| Phyla | Genus | Temperature Forest Biomes | Subtropical Forest Biomes | ||||
|---|---|---|---|---|---|---|---|
| P Starvation Response Regulation | P Uptake and Transport | P Solubilization and Mineralization | P Starvation Response Regulation | P Uptake and Transport | P Solubilization and Mineralization | ||
| Acidobacteria | Candidatus Solibacter | 1.67 ± 1.25 | 0 | 529 ± 48.42 | 22.33 ± 14.61 | 0 | 1014 ± 75.75 |
| Acidobacterium | 19.67 ± 8.73 | 78 ± 26.42 | 295.67 ± 118.42 | 10 ± 5.0 | 61.33 ± 23.33 | 120.67 ± 39.20 | |
| Candidatus Sulfopaludibacter | 1.33 ± 1.25 | 2.33 ± 1.25 | 185.67 ± 29.85 | 52.33 ± 32.36 | 10.67 ± 4.92 | 185 ± 113.93 | |
| Luteitalea | 9 ± 4.32 | 4.67.1.25 | 46 ± 5.1 | 7.33 ± 6.18 | 7.67 ± 4.64 | 55.67 ± 29.80 | |
| Edaphobacter | 0 | 8 ± 2.16 | 16.33 ± 6.18 | 0 | 26.33 ± 9.0 | 147.67 ± 58.63 | |
| Terriglobus | 0 | 0 | 29 ± 9.27 | 0 | 0 | 71 ± 12.33 | |
| Granulicella | 0 | 6.33 ± 4.50 | 8.33 ± 4.03 | 0 | 105.67 ± 45.71 | 31.67 ± 8.7 | |
| Occallatibacter | 0 | 0 | 23.67 ± 19.94 | 0 | 0 | 30.33 ± 15.69 | |
| Candidatus Koribacter | 0 | 0 | 17.33 ± 16.78 | 0 | 0 | 29.33 ± 8.22 | |
| Acidipila | 0.66 ± 0.94 | 1.33 ± 1.25 | 4.33 ± 3.30 | 0.33 ± 0.47 | 2 ± 1.41 | 23.33 ± 10.65 | |
| Proteobacteria | Bradyrhizobium | 188.33 ± 28.79 | 1590.33 ± 303.47 | 1609.67 ± 261.41 | 473.67 ± 85.34 | 3309 ± 930.05 | 3200 ± 853.32 |
| Rhodoplanes | 71 ± 6.48 | 533 ± 76.54 | 135.67 ± 23.68 | 117.33 ± 27.88 | 950.67 ± 195.21 | 406.67 ± 162.39 | |
| Sphingomonas | 109.33 ± 18.87 | 314.67 ± 105.09 | 203.67 ± 32.06 | 124.33 ± 73.30 | 323.33 ± 176.36 | 308.67 ± 82.70 | |
| Pseudolabrys | 107.33 ± 10.78 | 247.67 ± 53.51 | 41.33 ± 18.55 | 223.33 ± 40.84 | 416.67 ± 61.67 | 79.67 ± 18.80 | |
| Mesorhizobium | 13 ± 4.90 | 181 ± 38.79 | 91 ± 9.27 | 10.33 ± 4.92 | 264 ± 42.55 | 104.67 ± 32.27 | |
| Rhodopseudomonas | 19.67 ± 2.49 | 164 ± 24.49 | 50 ± 10.42 | 42 ± 10.61 | 225.67 ± 53.39 | 98.67 ± 25.04 | |
| Variibacter | 22 ± 9.93 | 156 ± 25.81 | 9.67 ± 3.01 | 19.33 ± 6.94 | 115 ± 36.08 | 16.67 ± 7.41 | |
| Bosea | 2.67 ± 1.70 | 152.33 ± 24.42 | 9.67 ± 2.05 | 0.67 ± 0.94 | 195 ± 29.80 | 9.33 ± 0.47 | |
| Enhydrobacter | 14.33 ± 5.25 | 93 ± 4.90 | 16.67 ± 4.50 | 30 ± 4.55 | 218.67 ± 29.23 | 37 ± 5.89 | |
| Afipia | 26.67 ± 4.19 | 78.67 ± 8.65 | 96 ± 9.41 | 21 ± 2.94 | 127 ± 35.36 | 138 ± 49.61 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Li, Y.; Wang, J.; He, L.; Wang, J.; Guo, Y.; Zhao, F. Contrasting Soil Microbial Functional Potential for Phosphorus Cycling in Subtropical and Temperate Forests. Forests 2022, 13, 2002. https://doi.org/10.3390/f13122002
Zhou S, Li Y, Wang J, He L, Wang J, Guo Y, Zhao F. Contrasting Soil Microbial Functional Potential for Phosphorus Cycling in Subtropical and Temperate Forests. Forests. 2022; 13(12):2002. https://doi.org/10.3390/f13122002
Chicago/Turabian StyleZhou, Sha, Yi Li, Jieying Wang, Liyuan He, Jun Wang, Yaoxin Guo, and Fazhu Zhao. 2022. "Contrasting Soil Microbial Functional Potential for Phosphorus Cycling in Subtropical and Temperate Forests" Forests 13, no. 12: 2002. https://doi.org/10.3390/f13122002