Deciphering the Ramet System of a Bamboo Plant in Response to Intensive Management


Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Location and Materials
2.2. Experiment Design
2.3. Data Investigation and Analysis
2.3.1. Underground Rhizome Excavation
2.3.2. Morphological Characteristics Investigation
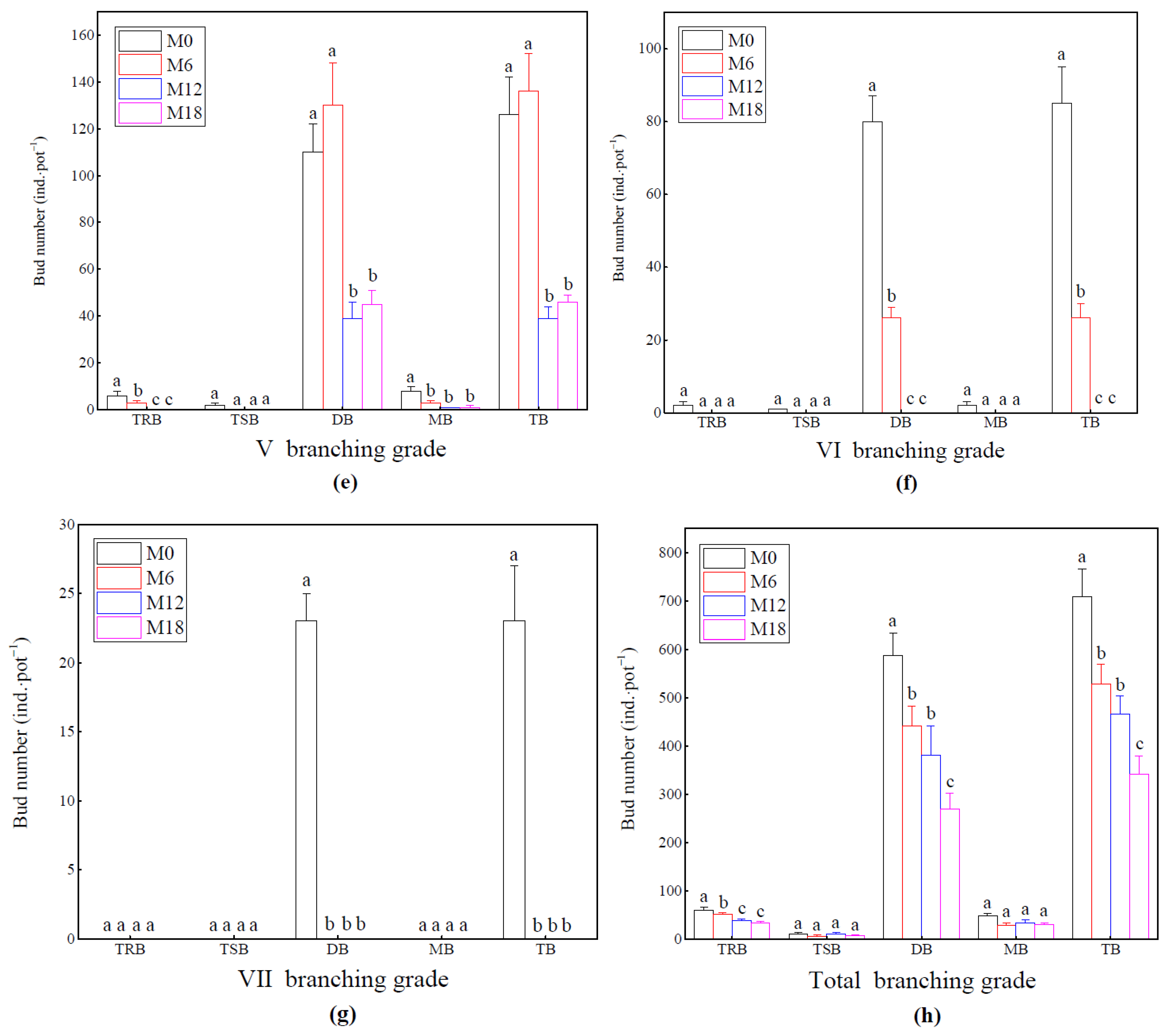
2.3.3. Bud Bank Investigation
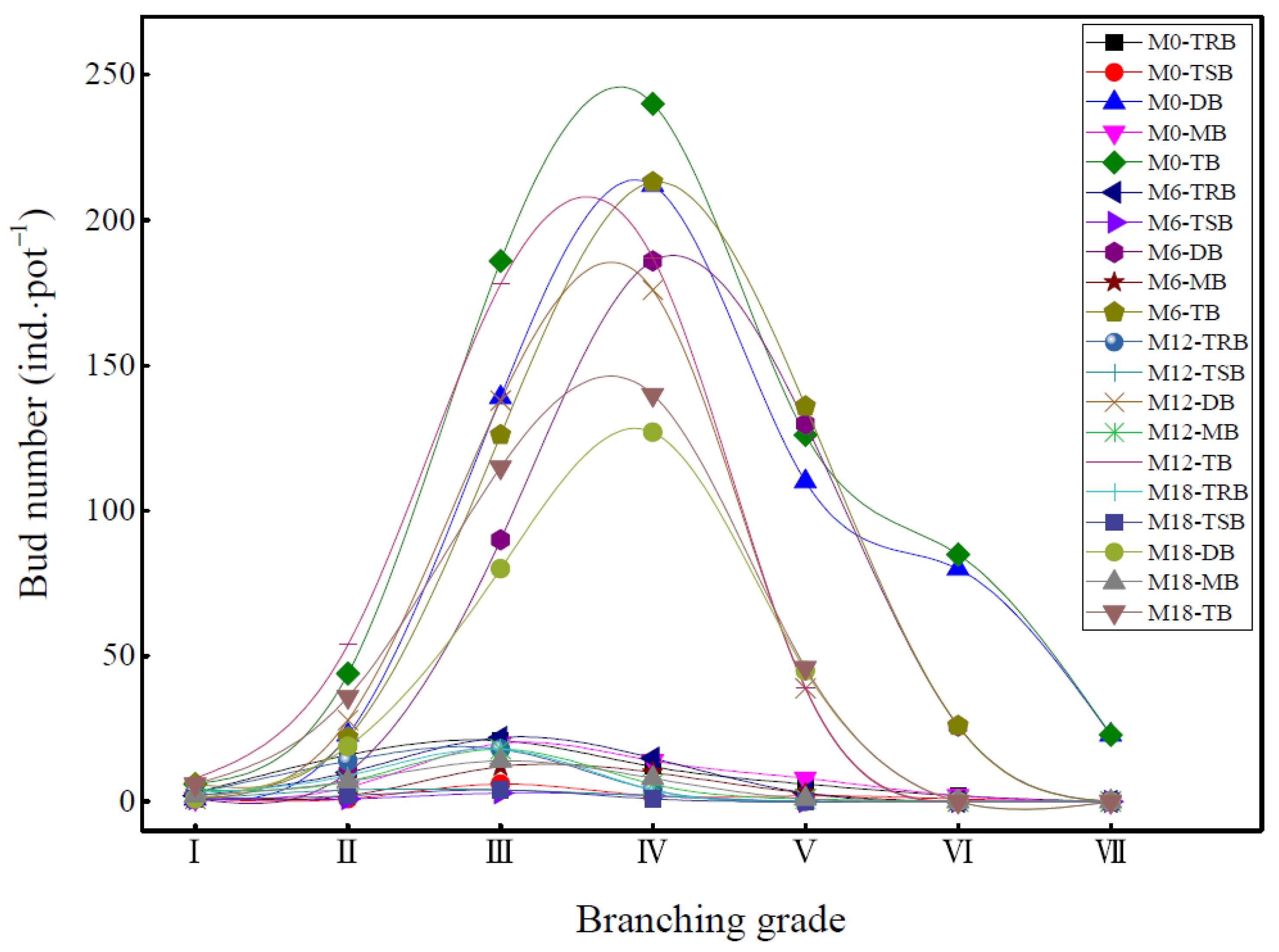
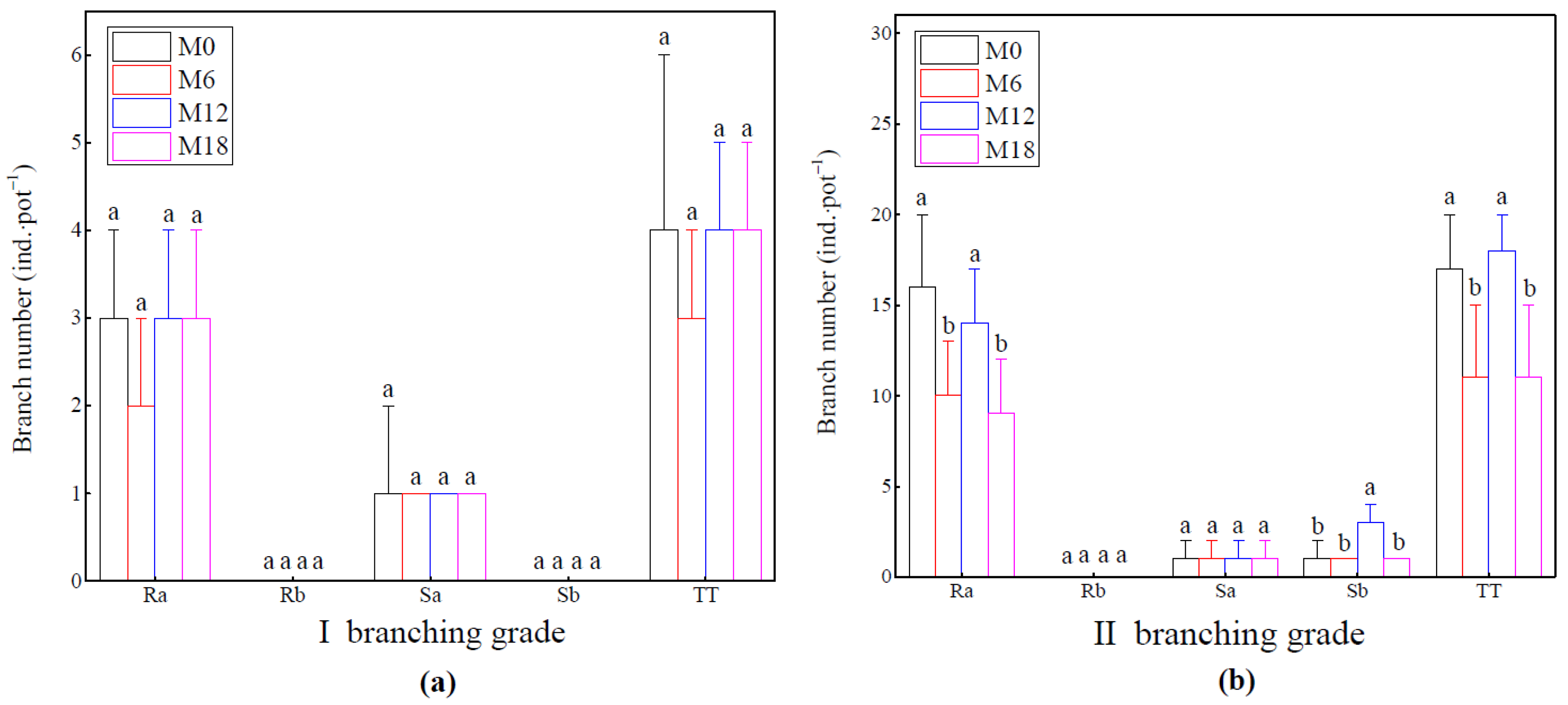
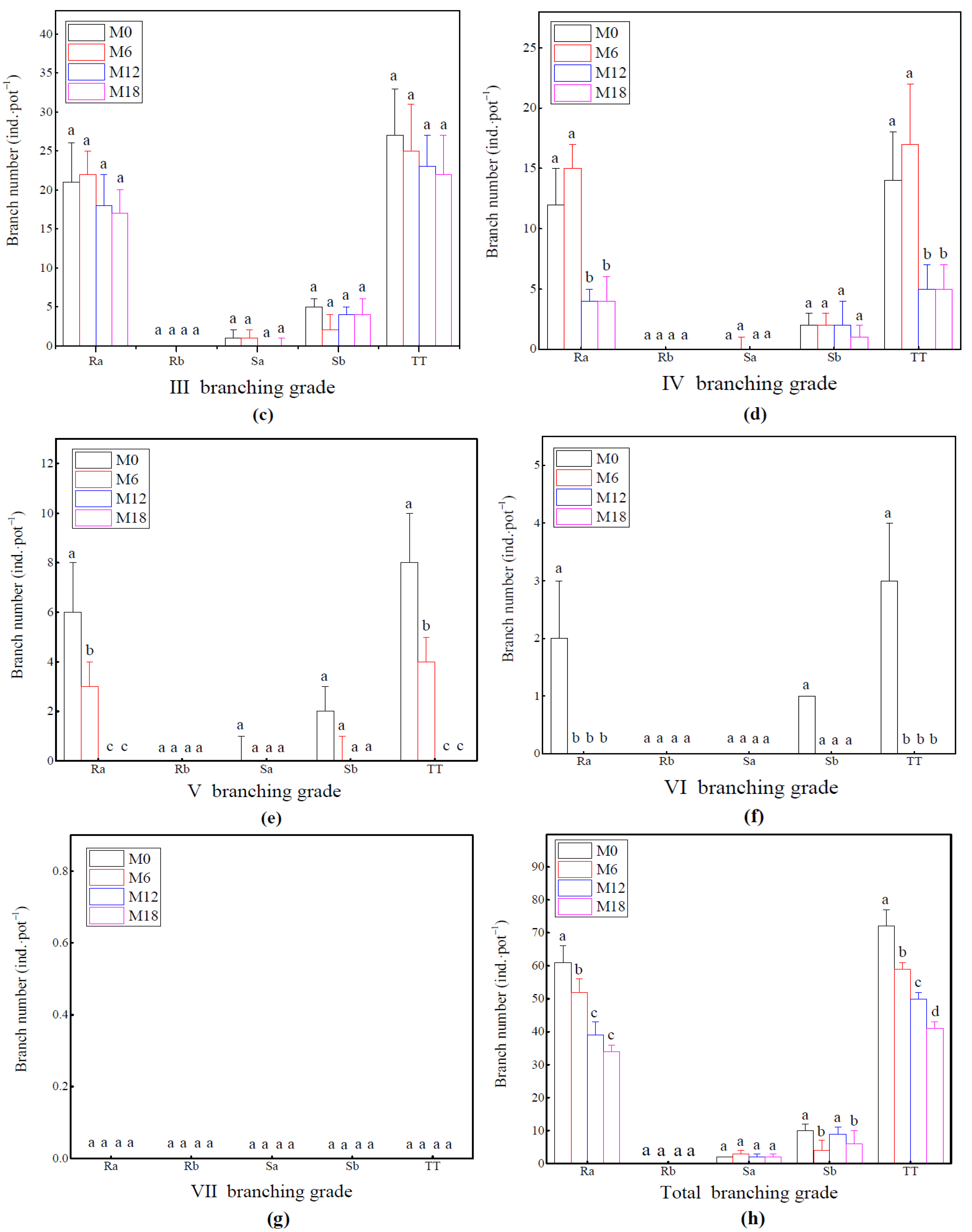
2.3.4. Branching Growth Investigation
2.3.5. Data Analysis
3. Results
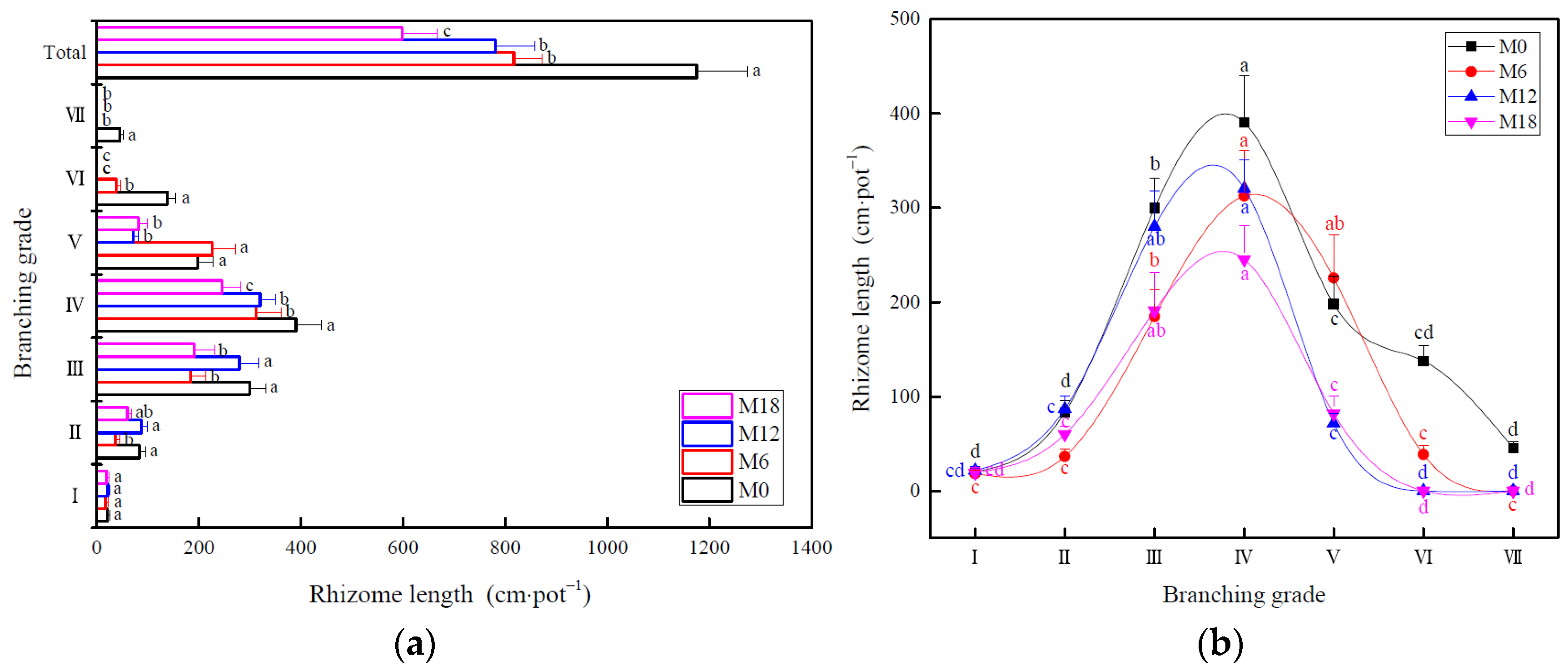
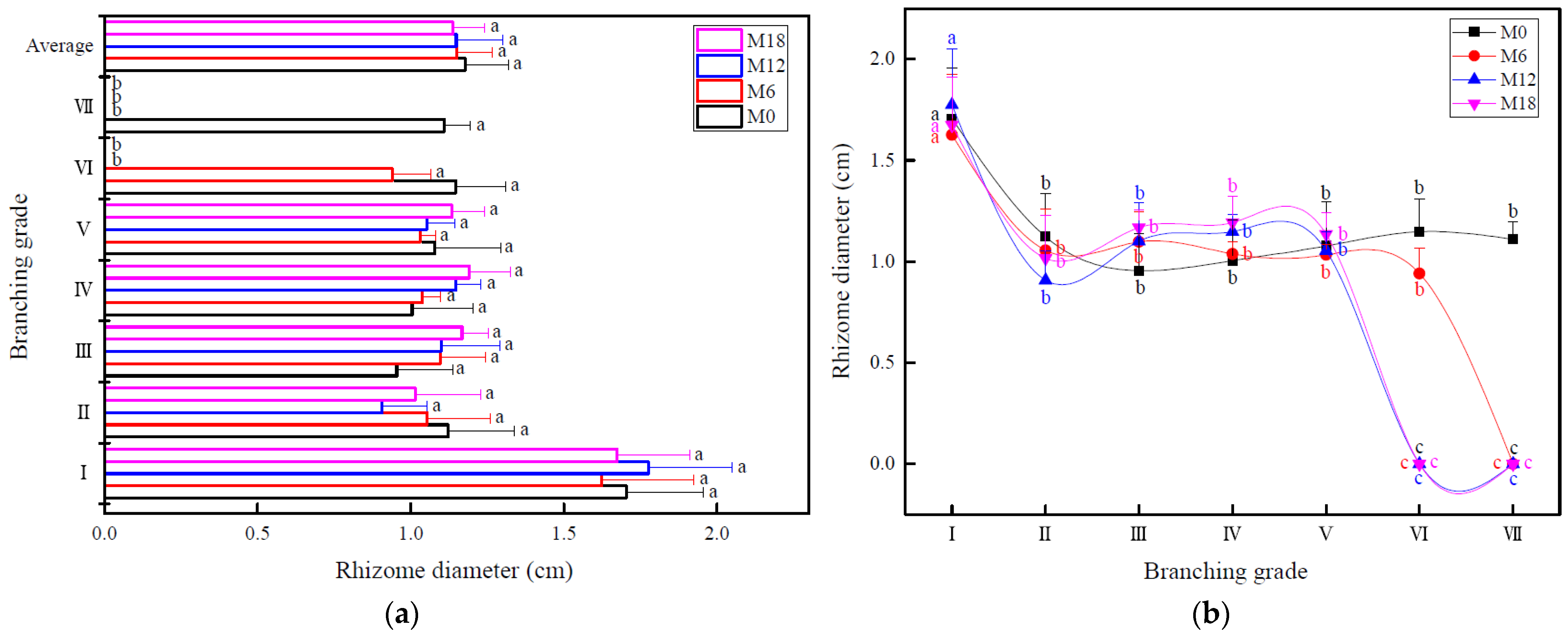
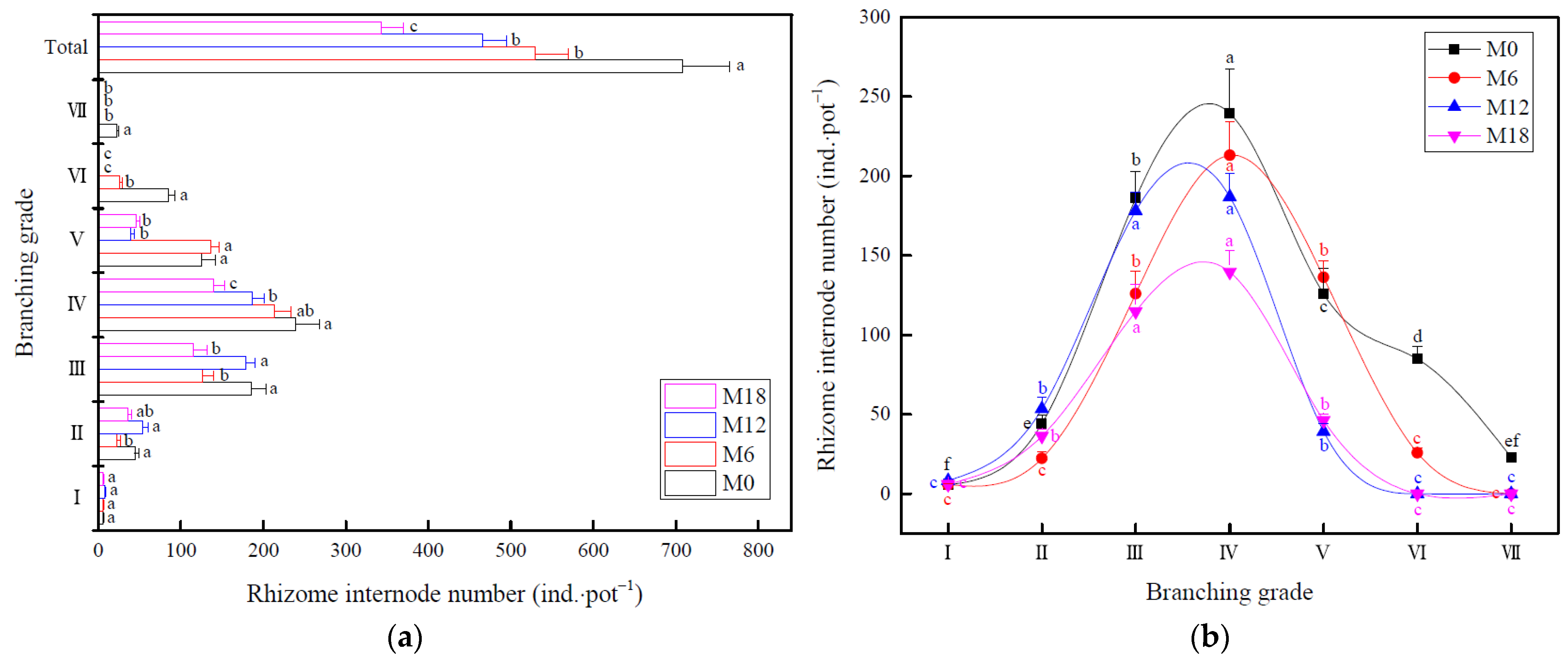
3.1. Changes in Rhizome Morphology
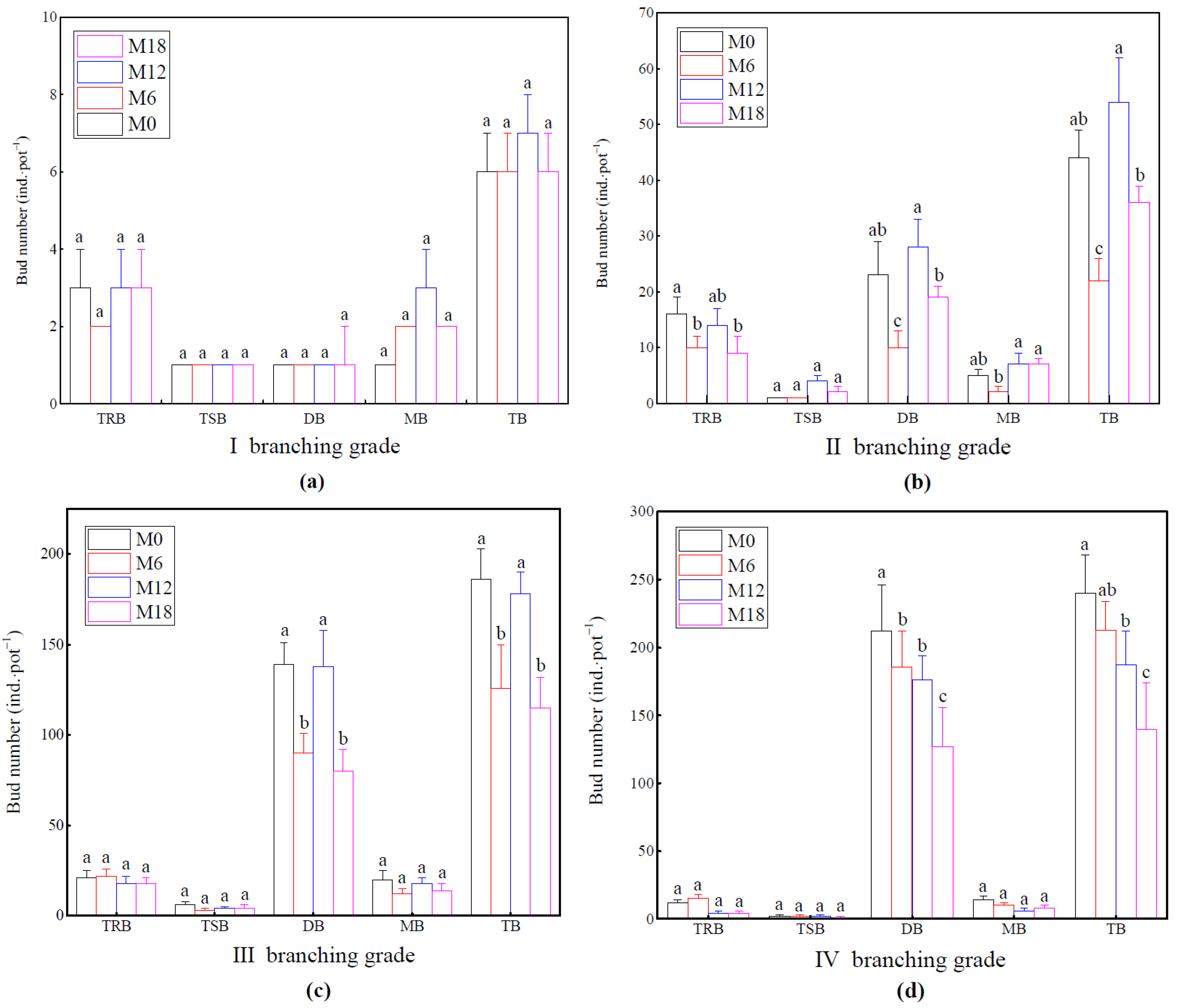
3.2. Response of Rhizome Bud Bank of Bamboo Ramet System
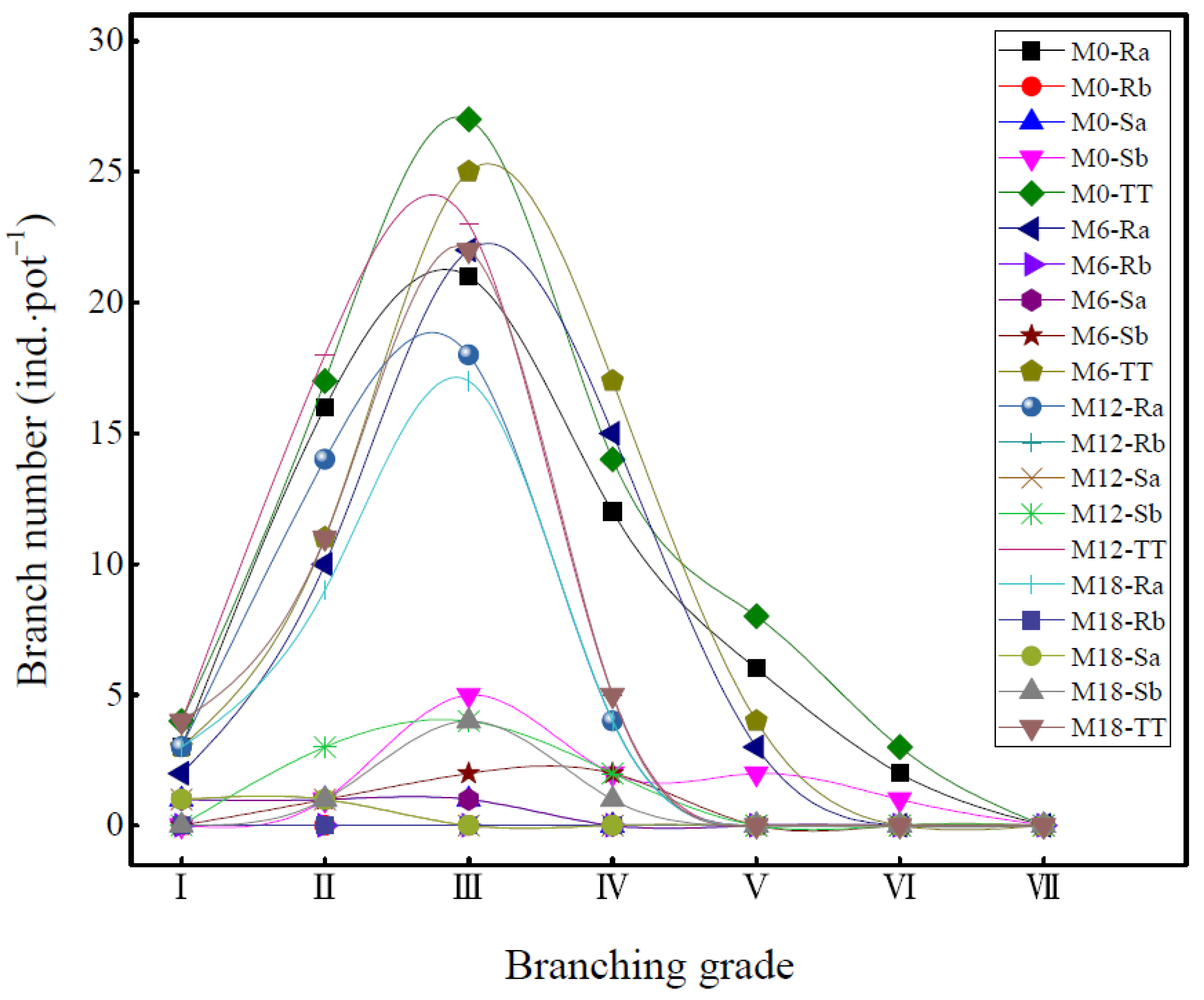
3.3. Branch Growth Difference of Bamboo Ramet System
4. Discussion
4.1. Effect of Intensive Management on Rhizome Morphology
4.2. Effect of Intensive Management on Rhizome Bud Bank
4.3. Effect of Intensive Management on Branch Growth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franklin, S.; Alpert, P.; Salguero-Gómez, R.; Janovský, Z.; Herben, T.; Klimešová, J.; Douhovnikoff, V. Next-gen plant clonal ecology. Perspect. Plant Ecol. 2021, 49, 125601. [Google Scholar] [CrossRef]
- Oborny, B. The plant body as a network of semi-autonomous agents: A review. Philos. Trans. R. Soc. B 2019, 374, 20180371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahru, T.; Kidane, B.; Mulatu, Y. Provenance variation on early survival rate and growth performance of Oxytenanthera abyssinica (A. Rich.) munro seedlings at green house: An indigenous lowland bamboo species in ethiopia. Int. J. For. Res. 2018, 2018, 5713456. [Google Scholar]
- Zhang, W.; Hu, T.; Chang, Y.; Fei, B.; Ma, Y.; Deng, Y.; Xia, M.; Fan, K.; Zhang, X.; Jiang, Z. Correlation between genetic characteristics, cell structure and material properties of Moso Bamboo (Phyllostachys edulis (Carriere) J. Houzeau) in different areas of China. Forests 2022, 13, 107. [Google Scholar] [CrossRef]
- Meena, R.K.; Bhandhari, M.S.; Barhwal, S.; Ginwal, H.S. Genetic diversity and structure of Dendrocalamus hamiltonii natural metapopulation: A commercially important bamboo species of northeast Himalayas. 3 Biotech 2019, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Du, H.; Mao, F.; Zhou, G.; Xing, L.; Liu, T.; Han, N.; Liu, E.; Ge, H.; Liu, Y.; et al. Mapping spatiotemporal decisions for sustainable productivity of bamboo forest land. Land Degrad. Dev. 2020, 31, 939–958. [Google Scholar] [CrossRef]
- Tambe, S.; Patnaik, S.; Upadhyay, A.P.; Edgaonkar, A.; Singhal, R.; Bisaria, J.; Srivastava, P.; Dahake, K.; Hiralal, M.H.; Tofa, D.; et al. Assessing the sustainability of bamboo management in central Indian forests. For. Trees Livelihoods 2021, 30, 28–46. [Google Scholar] [CrossRef]
- Zhou, Z.; Tang, Y.; Xu, H.; Wang, J.; Hu, L.; Xu, X. Dynamic changes in leaf biomass and the modeling of individual Moso Bamboo (Phyllostachys edulis (Carrière) J. Houz) under intensive management. Forests 2022, 13, 693. [Google Scholar] [CrossRef]
- Huy, B.; Thanh, G.T.; Poudel, K.P.; Temesgen, H. Individual plant allometric equations for estimating aboveground biomass and its components for a common bamboo species (Bambusa procera A. Chev. and A. Camus) in tropical forests. Forests 2019, 10, 316. [Google Scholar] [CrossRef] [Green Version]
- Ely, F.; Rada, F.; Fermin, G.; Clark, L.G. Ecophysiology and genetic diversity in species of the bamboo Chusquea in the high Andes, Venezuela. Plant Ecol. Divers. 2019, 12, 555–572. [Google Scholar] [CrossRef]
- Huang, L.; Xing, X.C.; Li, W.W.; Zhou, Y.; Zhang, Y.Q.; Xue, C.; Ren, Y.; Kang, J.Q. Population genetic structure of the giant panda staple food bamboo (Fargesia spathacea complex) and its taxonomic implications. J. Syst. Evol. 2021, 59, 1051–1064. [Google Scholar] [CrossRef]
- Boban, S.; Maurya, S.; Jha, Z. DNA fingerprinting: An overview on genetic diversity studies in the botanical taxa of Indian Bamboo. Genet. Resour. Crop Evol. 2022, 69, 469–498. [Google Scholar] [CrossRef]
- Dlamini, L.C.; Fakudze, S.; Makombe, G.G.; Muse, S.; Zhu, J. Bamboo as a valuable resource and its utilization in historical and modern-day China. BioResources 2022, 17, 1926–1938. [Google Scholar] [CrossRef]
- Qian, Z.; Zhuang, S.; Gao, J.; Tang, L. Can aeration improve bamboo soil fertility of soil below bamboo and fungal diversity under mulching conditions? Land Degrad. Dev. 2022, 33, 2353–2365. [Google Scholar] [CrossRef]
- Qian, Z.; Zhuang, S.; Gui, R.; Tang, L. Effect of soil aeration treatment on the physiological and biochemical characteristics of Phyllostachys praecox under the organic material mulching. Plant Soil 2021, 459, 357–369. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, S.; Jiang, H.; Peng, C.; Zhang, J.; Zhou, G. The impact of intensive management on net ecosystem productivity and net primary productivity of a Lei bamboo forest. Ecol. Model. 2020, 435, 109248. [Google Scholar] [CrossRef]
- Gui, R.; Wu, W.; Zhuang, S.; Zhong, Z. Intensive management increases soil acidification and phytotoxic Al content in Phyllostachys praecox stands in Southeast China. J. Sustain. For. 2018, 37, 46–55. [Google Scholar] [CrossRef]
- Chen, J.; Wu, Q.; Li, S.; Ge, J.; Liang, C.; Qin, H.; Xu, Q.; Fuhrmann, J.J. Diversity and function of soil bacterial communities in response to long-term intensive management in a subtropical bamboo forest. Geoderma 2019, 354, 113894. [Google Scholar] [CrossRef]
- Adomako, M.O.; Zhang, Q.; Yu, F.H. Genotypic differences in response to different patterns of clonal fragmentation in the aquatic macrophyte Pistia stratiotes. J. Plant Ecol. 2022, rtac035. [Google Scholar] [CrossRef]
- Adomako, M.O.; Alpert, P.; Du, D.L.; Yu, F.H. Effects of fragmentation of clones compound over vegetative generations in the floating plant Pistia stratiotes. Ann. Bot. 2021, 127, 123–133. [Google Scholar] [CrossRef]
- He, L.; Xiao, X.; Zhang, X.; Jin, Y.; Pu, Z.; Lei, N.; He, X.; Chen, J. Clonal fragments of stoloniferous invasive plants benefit more from stolon storage than their congeneric native species. Flora 2021, 281, 151877. [Google Scholar] [CrossRef]
- Gao, G.; Wu, Z.; Xing, W.; Zhang, X.; Zhong, H.; Pan, Y. Bud population characteristics of Phyllostachys praecox ‘prevernalis’ under different mulching cultivation periods. Bangl. J. Bot. 2018, 47, 969–974. [Google Scholar] [CrossRef]
- Wang, J.; Abdullah, I.; Xu, T.; Zhu, W.; Gao, Y.; Wang, L. Effects of mowing disturbance and competition on spatial expansion of the clonal plant Leymus chinensis into saline-alkali soil patches. Environ. Exp. Bot. 2019, 168, 103890. [Google Scholar] [CrossRef]
- Reijers, V.C.; Hoeks, S.; van Belzen, J.; Siteur, K.; de Rond, A.J.A.; van de Ven, C.N.; Lammers, C.; van de Koppel, J.; van der Heide, T. Sediment availability provokes a shift from Brownian to Lévy-like clonal expansion in a dune building grass. Ecol. Lett. 2021, 24, 258–268. [Google Scholar] [CrossRef]
- Guo, F.H.; Li, X.L.; Yin, J.J.; Jimoh, S.O.; Hou, X.Y. Grazing-induced legacy effects enhance plant adaption to drought by larger root allocation plasticity. J. Plant Ecol. 2021, 14, 1024–1029. [Google Scholar] [CrossRef]
- Pliszko, A.; Kostrakiewicz-Gierałt, K. The importance of sexual, asexual and mixed ramet clusters in production of descendant ramets in populations of Solidago × niederederi (Asteraceae). Biologia 2019, 74, 953–960. [Google Scholar] [CrossRef] [Green Version]
- Ott, J.P.; Klimešová, J.; Hartnett, D.C. The ecology and significance of below-ground bud banks in plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef]
- Zheng, X.; Gao, Y.; Wang, Y.; Xing, F.; Zhao, M.; Gao, Y. Optimal foraging strategies in varying nutrient heterogeneity: Responses of a stoloniferous clonal plant to patch pattern, size and quality. Écoscience 2022, 29, 221–232. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Ma, Q.; Liu, Z. Roles of aboveground vegetation, soil properties, and disturbance in determining belowground bud bank in sand dune ecosystems. Environ. Exp. Bot. 2020, 178, 104155. [Google Scholar] [CrossRef]
- Alhaj Hamoud, Y.; Shaghaleh, H.; Sheteiwy, M.; Guo, X.; Elshaikh, N.A.; Ullah Khan, N.; Oumarou, A.; Rahim, S.F. Impact of alternative wetting and soil drying and soil clay content on the morphological and physiological traits of rice roots and their relationships to yield and nutrient use-efficiency. Agric. Water Manag. 2019, 223, 105706. [Google Scholar] [CrossRef]
- Wielgusz, K.; Praczyk, M.; Irzykowska, L.; Świerk, D. Fertilization and soil pH affect seed and biomass yield, plant morphology, and cadmium uptake in hemp (Cannabis sativa L.). Ind. Crop. Prod. 2022, 175, 114245. [Google Scholar] [CrossRef]
- Yu, H.; Wang, L.; Liu, C.; Yu, D.; Qu, J. Effects of a spatially heterogeneous nutrient distribution on the growth of clonal wetland plants. BMC Ecol. 2020, 20, 59. [Google Scholar] [CrossRef]
- Zhang, L.M.; Alpert, P.; Yu, F.H. Nutrient foraging ability promotes intraspecific competitiveness in the clonal plant Hydrocotyle vulgaris. Ecol. Indic. 2022, 138, 108862. [Google Scholar] [CrossRef]
- Jiao, L.; Liu, X.; Wang, S.; Dong, X.; Li, F.; Xin, C. Spatial expansion strategy of the clonal modules for Phragmites australis and response to environmental factors in an inland river wetland. Pol. J. Environ. Stud. 2020, 29, 1185–1196. [Google Scholar] [CrossRef]
- Martin, F.M.; Dommanget, F.; Lavallée, F.; Evette, A. Clonal growth strategies of Reynoutria japonica in response to light, shade, and mowing, and perspectives for management. NeoBiota 2020, 56, 89–110. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potted Soil | pH | Conductivity (μs·cm−1) | Organic Matter (g·kg−1) | Hydrolytic Nitrogen (mg·kg−1) | Available Phosphorus (mg·kg−1) | Available Potassium (mg·kg−1) |
|---|---|---|---|---|---|---|
| M 0 | 6.48 | 101.6 | 33.07 | 42.56 | 45.45 | 465.84 |
| M 6 | 5.25 | 69.88 | 21.61 | 53.20 | 35.95 | 418.52 |
| M 12 | 4.79 | 54.57 | 33.17 | 107.50 | 43.97 | 539.04 |
| M 18 | 4.76 | 213.7 | 41.37 | 163.86 | 65.61 | 552.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, G.; Wen, X.; Wu, Z.; Zhong, H.; Zhang, X. Deciphering the Ramet System of a Bamboo Plant in Response to Intensive Management. Forests 2022, 13, 1968. https://doi.org/10.3390/f13111968
Gao G, Wen X, Wu Z, Zhong H, Zhang X. Deciphering the Ramet System of a Bamboo Plant in Response to Intensive Management. Forests. 2022; 13(11):1968. https://doi.org/10.3390/f13111968
Chicago/Turabian StyleGao, Guibin, Xing Wen, Zhizhuang Wu, Hao Zhong, and Xiaoping Zhang. 2022. "Deciphering the Ramet System of a Bamboo Plant in Response to Intensive Management" Forests 13, no. 11: 1968. https://doi.org/10.3390/f13111968