
Calibration of Thermal Dissipation Probes for Xylem Sap Flow in the Wood of a Diffuse-Porous and a Conifer Species under Cyclic Heating

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Measurement
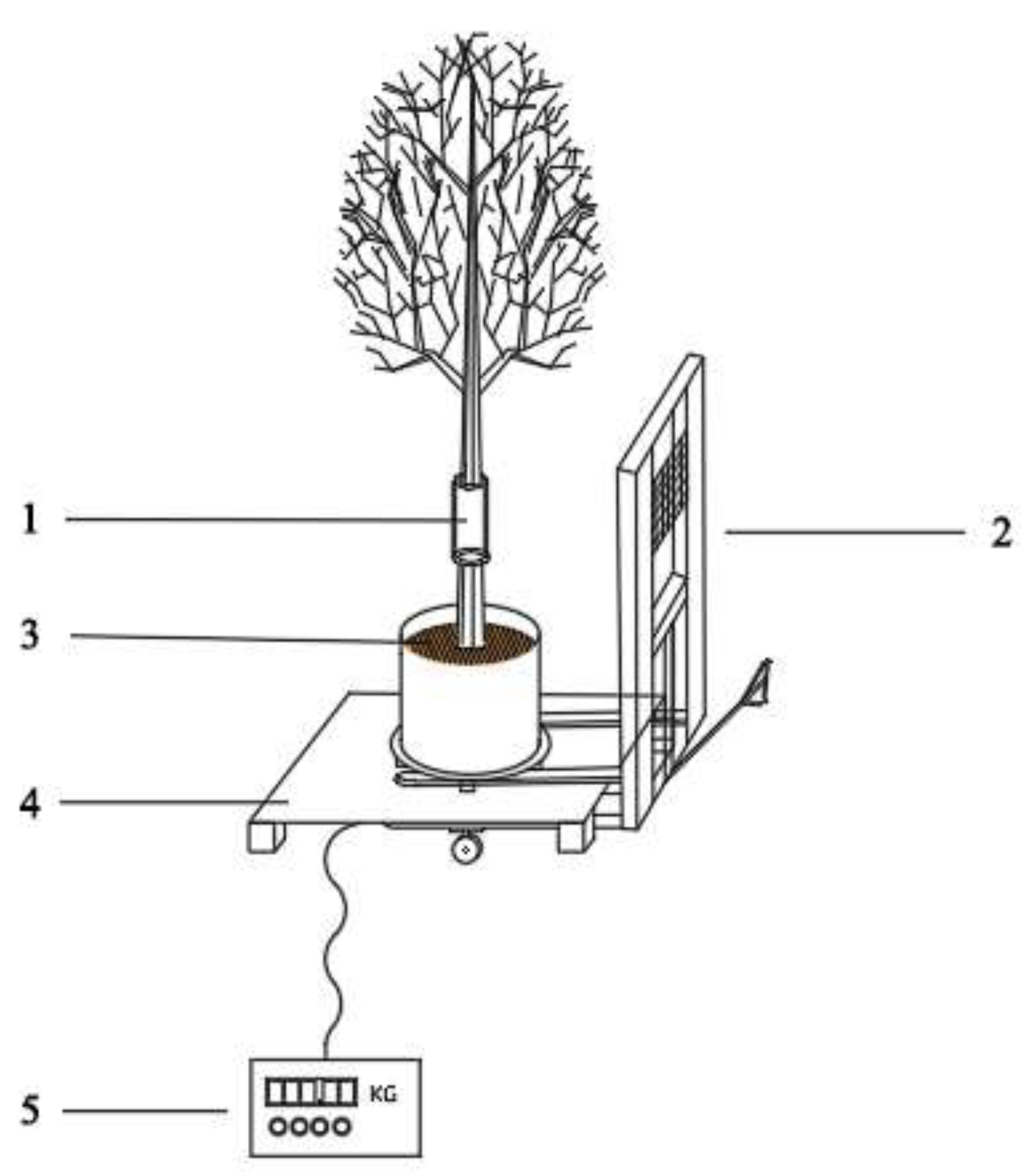
2.2.1. Sap Flow Measurement
2.2.2. Reference Flow Rate Measurement
2.3. Statistical Analysis
3. Results
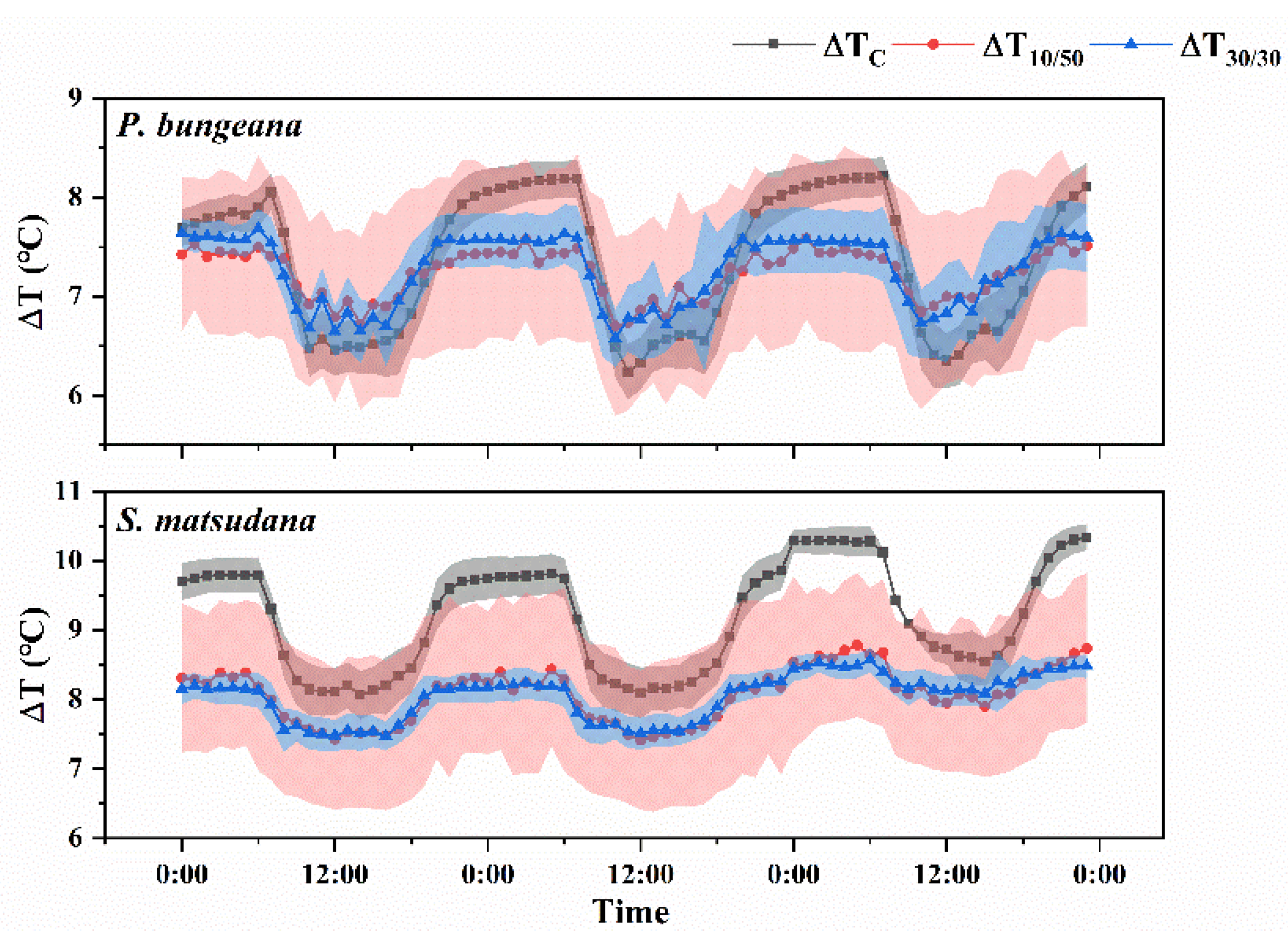
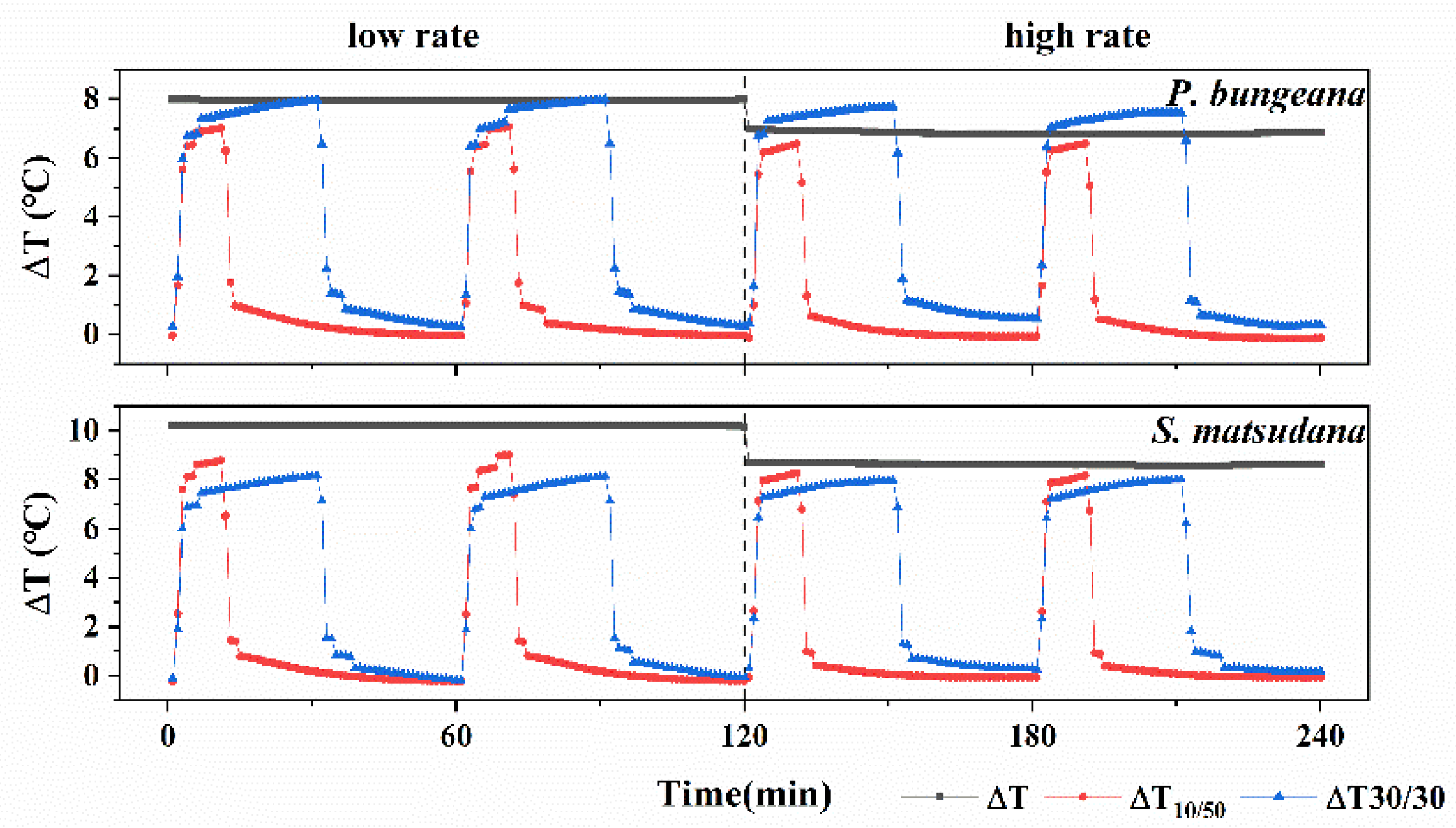
3.1. Tendency of Temperature Difference under Different Heating Modes
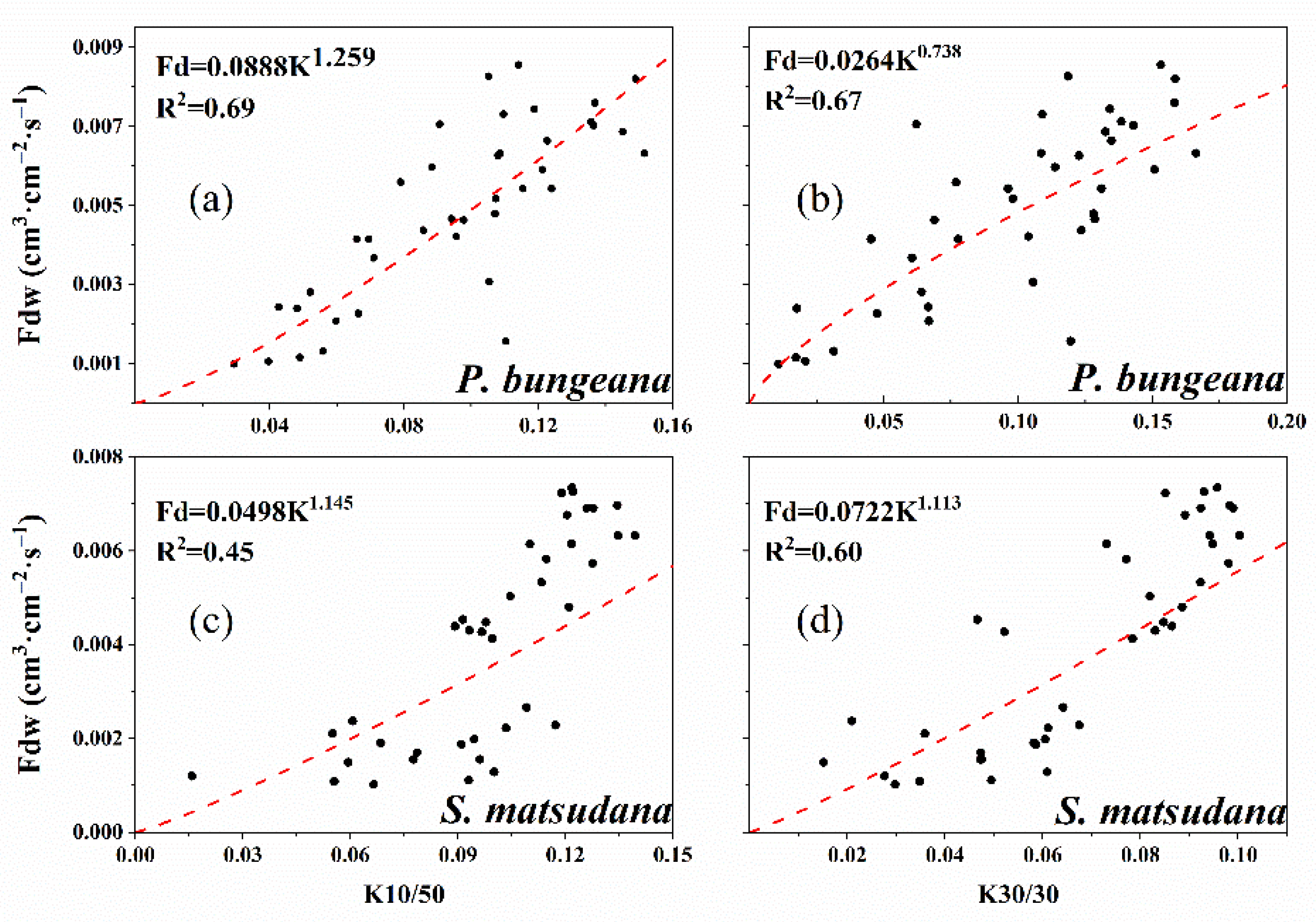
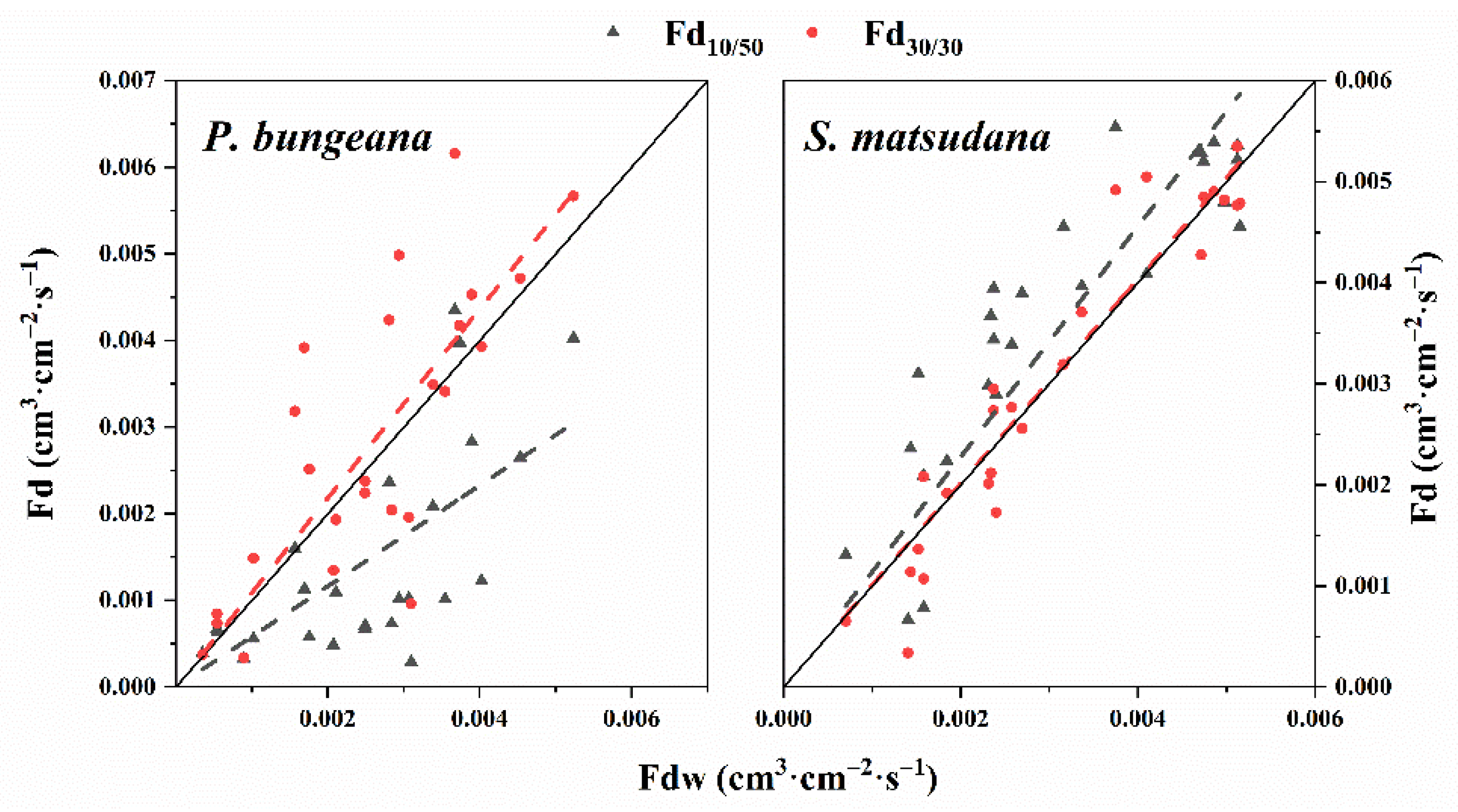
3.2. Calibration Equations
3.3. Validation of the Calibration Equations
4. Discussion
4.1. Accuracy of Sap Flow Density Estimates in the Field
4.2. Benefits and Limitations of the Cyclic Heating Mode
4.3. The Uncertainty of Outcome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green, S.; Clothier, B.; Jardine, B. Theory and practical application of heat pulse to measure sap flow. Agron. J. 2003, 95, 1371–1379. [Google Scholar] [CrossRef]
- Peramaki, M.; Vesala, T.; Nikinmaa, E. Analysing the applicability of the heat balance method for estimating sap flow in boreal forest conditions. Boreal Environ. Res. 2001, 6, 29–43. [Google Scholar]
- Granier, A. Une nouvelle méthode pour la mesure du flux de sève brute dans le tronc des arbres. Ann. Sci. For. 1985, 42, 193–200. [Google Scholar] [CrossRef]
- Flo, V.; Martinez-Vilalta, J.; Steppe, K.; Schuldt, B.; Poyatos, R. A synthesis of bias and uncertainty in sap flow methods. Agric. For. Meteorol. 2019, 271, 362–374. [Google Scholar] [CrossRef]
- Chen, Y.J.; Bongers, F.; Tomlinson, K.; Fan, Z.X.; Lin, H.; Zhang, S.B.; Zheng, Y.L.; Li, Y.P.; Cao, K.F.; Zhang, J.L. Time lags between crown and basal sap flows in tropical lianas and co-occurring trees. Tree Physiol. 2016, 36, 736–747. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.J.; Zhang, Y.J.; Goldstein, G.; Sun, M.; Ma, R.Y.; Cao, K.F. Determinants of water circulation in a woody bamboo species: Afternoon use and night-time recharge of culm water storage. Tree Physiol. 2015, 35, 964–974. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, N.; Kumagai, T.; Miyazawa, Y.; Matsumoto, K.; Tateishi, M.; Lim, T.K.; Mudd, R.G.; Ziegler, A.D.; Giambelluca, T.W.; Yin, S. Transpiration characteristics of a rubber plantation in central Cambodia. Tree Physiol. 2014, 34, 285–301. [Google Scholar] [CrossRef] [Green Version]
- Rabbel, I.; Bogena, H.; Neuwirth, B.; Diekkruger, B. Using Sap Flow Data to Parameterize the Feddes Water Stress Model for Norway Spruce. Water 2018, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Hoffmann, W.A.; Meinzer, F.C.; Franco, A.C.; Giambelluca, T.; Miralles-Wilhelm, F. Controls on stand transpiration and soil water utilization along a tree density gradient in a Neotropical savanna. Agric. For. Meteorol. 2008, 148, 839–849. [Google Scholar] [CrossRef]
- Jung, E.Y.; Otieno, D.; Lee, B.; Lim, J.H.; Kang, S.K.; Schmidt, M.W.T.; Tenhunen, J. Up-scaling to stand transpiration of an Asian temperate mixed-deciduous forest from single tree sapflow measurements. Plant Ecol. 2011, 212, 383–395. [Google Scholar] [CrossRef]
- Lu, P.; Urban, L.; Zhao, P. Granier’s thermal dissipation probe (TDP) method for measuring sap flow in trees: Theory and practice. Acta Bot. Sin. 2004, 46, 631–646. [Google Scholar]
- Goulden, M.L.; Field, C.B. 3 Methods for Monitoring the Gas-Exchange of Individual Tree Canopies—Ventilated-Chamber, Sap-Flow and Penman-Monteith Measurements on Evergreen Oaks. Funct. Ecol. 1994, 8, 125–135. [Google Scholar] [CrossRef]
- Taneda, H.; Sperry, J.S. A case-study of water transport in co-occurring ring- versus diffuse-porous trees: Contrasts in water-status, conducting capacity, cavitation and vessel refilling. Tree Physiol. 2008, 28, 1641–1651. [Google Scholar] [CrossRef] [Green Version]
- Bush, S.E.; Hultine, K.R.; Sperry, J.S.; Ehleringer, J.R. Calibration of thermal dissipation sap flow probes for ring- and diffuse-porous trees. Tree Physiol. 2010, 30, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Berdanier, A.B.; Miniat, C.F.; Clark, J.S. Predictive models for radial sap flux variation in coniferous, diffuse-porous and ring-porous temperate trees. Tree Physiol. 2016, 36, 932–941. [Google Scholar] [CrossRef] [Green Version]
- Baiamonte, G.; Motisi, A. Analytical approach extending the Granier method to radial sap flow patterns. Agric. Water Manag. 2020, 231, 105988. [Google Scholar] [CrossRef]
- Xie, J.; Wan, X.C. The accuracy of the thermal dissipation technique for estimating sap flow is affected by the radial distribution of conduit diameter and density. Acta Physiol. Plant. 2018, 40, 88. [Google Scholar] [CrossRef]
- Alizadeh, A.; Toudeshki, A.; Ehsani, R.; Migliaccio, K. Potential Sources of Errors in Estimating Plant Sap Flow Using Commercial Thermal Dissipation Probes. Appl. Eng. Agric. 2018, 34, 899–906. [Google Scholar] [CrossRef]
- Smith, D.M.; Allen, S.J. Measurement of sap flow in plant stems. J. Exp. Bot. 1996, 47, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Cermak, J.; Kucera, J. The Compensation of Natural Temperature-Gradient at the Measuring Point during the Sap Flow-Rate Determination in Trees. Biol. Plant. 1981, 23, 469–471. [Google Scholar] [CrossRef]
- Masmoudi, M.M.; Mahjoub, I.; Lhomme, J.P.; Ben Mechlia, N. Sap flow measurement by a single thermal dissipation probe in transient regime: Implementation of the method and test under field conditions. Ann. For. Sci. 2012, 69, 773–781. [Google Scholar] [CrossRef]
- Do, F.; Rocheteau, A. Influence of natural temperature gradients on measurements of xylem sap flow with thermal dissipation probes. 1. Field observations and possible remedies. Tree Physiol. 2002, 22, 641–648. [Google Scholar] [CrossRef]
- Do, F.; Rocheteau, A. Influence of natural temperature gradients on measurements of xylem sap flow with thermal dissipation probes. 2. Advantages and calibration of a noncontinuous heating system. Tree Physiol. 2002, 22, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Ayutthaya, S.I.N.; Do, F.C.; Pannengpetch, K.; Junjittakarn, J.; Maeght, J.L.; Rocheteau, A.; Cochard, H. Transient thermal dissipation method of xylem sap flow measurement: Multi-species calibration and field evaluation. Tree Physiol. 2010, 30, 139–148. [Google Scholar] [CrossRef]
- Lopez, J.G.; Licata, J.; Pypker, T.; Asbjornsen, H. Effects of heater wattage on sap flux density estimates using an improved tree-cut experiment. Tree Physiol. 2019, 39, 679–693. [Google Scholar] [CrossRef]
- Pasqualotto, G.; Carraro, V.; Menardi, R.; Anfodillo, T. Calibration of Granier-Type (TDP) Sap Flow Probes by a High Precision Electronic Potometer. Sensors 2019, 19, 2419. [Google Scholar] [CrossRef] [Green Version]
- Sperling, O.; Shapira, O.; Cohen, S.; Tripler, E.; Schwartz, A.; Lazarovitch, N. Estimating sap flux densities in date palm trees using the heat dissipation method and weighing lysimeters. Tree Physiol. 2012, 32, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Tfwala, C.M.; van Rensburg, L.D.; Bello, Z.A.; Green, S.R. Calibration of compensation heat pulse velocity technique for measuring transpiration of selected indigenous trees using weighing lysimeters. Agric. Water Manag. 2018, 200, 27–33. [Google Scholar] [CrossRef]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- McCulloh, K.A.; Winter, K.; Meinzer, F.C.; Garcia, M.; Aranda, J.; Lachenbruch, B. A, comparison of daily water use estimates derived from constant-heat sap-flow probe values and gravimetric measurements in pot-grown saplings. Tree Physiol. 2007, 27, 1355–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willmott, C.J. On the validation of models. Phys. Geogr. 1981, 2, 184–194. [Google Scholar] [CrossRef]
- Wang, Q.; Lintunen, A.; Zhao, P.; Shen, W.J.; Salmon, Y.; Chen, X.; Ouyang, L.; Zhu, L.W.; Ni, G.Y.; Sun, D.; et al. Assessing Environmental Control of Sap Flux of Three Tree Species Plantations in Degraded Hilly Lands in South China. Forests 2020, 11, 206. [Google Scholar] [CrossRef] [Green Version]
- Hayat, M.; Zha, T.S.; Jia, X.; Iqbal, S.; Qian, D.; Bourque, C.P.A.; Khan, A.; Tian, Y.; Bai, Y.J.; Liu, P.; et al. A multiple-temporal scale analysis of biophysical control of sap flow in Salix psammophila growing in a semiarid shrubland ecosystem of northwest China. Agric. For. Meteorol. 2020, 288, 107985. [Google Scholar] [CrossRef]
- Venturin, A.Z.; Guimaraes, C.M.; de Sousa, E.F.; Machado, J.A.; Rodrigues, W.P.; Serrazine, I.D.; Bressan-Smith, R.; Marciano, C.R.; Campostrini, E. Using a crop water stress index based on a sap flow method to estimate water status in conilon coffee plants. Agric. Water Manag. 2020, 241, 106343. [Google Scholar] [CrossRef]
- O’Keefe, K.; Bell, D.M.; McCulloh, K.A.; Nippert, J.B. Bridging the Flux Gap: Sap Flow Measurements Reveal Species-Specific Patterns of Water Use in a Tallgrass Prairie. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005446. [Google Scholar] [CrossRef]
- Sun, H.Z.; Aubrey, D.P.; Teskey, R.O. A simple calibration improved the accuracy of the thermal dissipation technique for sap flow measurements in juvenile trees of six species. Trees 2012, 26, 631–640. [Google Scholar] [CrossRef]
- Reyes-Acosta, J.L.; Vandegehuchte, M.W.; Steppe, K.; Lubczynski, M.W. Novel, cyclic heat dissipation method for the correction of natural temperature gradients in sap flow measurements. Part 2. Laboratory validation. Tree Physiol. 2012, 32, 913–929. [Google Scholar] [CrossRef] [Green Version]
- Lubczynski, M.W.; Chavarro-Rincon, D.; Roy, J. Novel, cyclic heat dissipation method for the correction of natural temperature gradients in sap flow measurements. Part 1. Theory and application. Tree Physiol. 2012, 32, 894–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nourtier, M.; Chanzy, A.; Granier, A.; Huc, R. Sap flow measurements by thermal dissipation method using cyclic heating: A processing method accounting for the non-stationary regime. Ann. For. Sci. 2011, 68, 1255–1264. [Google Scholar] [CrossRef] [Green Version]
- Do, F.C.; Ayutthaya, S.I.N.; Rocheteau, A. Transient thermal dissipation method for xylem sap flow measurement: Implementation with a single probe. Tree Physiol. 2011, 31, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Nhean, S.; Ayutthaya, S.I.N.; Rocheteau, A.; Do, F.C. Multi-species test and calibration of an improved transient thermal dissipation system of sap flow measurement with a single probe. Tree Physiol. 2019, 39, 1061–1070. [Google Scholar] [CrossRef]
- Ren, R.Q.; von der Crone, J.; Horton, R.; Liu, G.; Steppe, K. An improved single probe method for sap flow measurements using finite heating duration. Agric. For. Meteorol. 2020, 280, 107788. [Google Scholar] [CrossRef]
- Steppe, K.; De Pauw, D.J.W.; Doody, T.M.; Teskey, R.O. A comparison of sap flux density using thermal dissipation, heat pulse velocity and heat field deformation methods. Agric. For. Meteorol. 2010, 150, 1046–1056. [Google Scholar] [CrossRef]
- Rana, G.; De Lorenzi, F.; Palatella, L.; Martinelli, N.; Ferrara, R.M. Field scale recalibration of the sap flow thermal dissipation method in a Mediterranean vineyard. Agric. For. Meteorol. 2019, 269, 169–179. [Google Scholar] [CrossRef]
- James, S.A.; Clearwater, M.J.; Meinzer, F.C.; Goldstein, G. Heat dissipation sensors of variable length for the measurement of sap flow in trees with deep sapwood. Tree Physiol. 2002, 22, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Dichio, B.; Montanaro, G.; Sofo, A.; Xiloyannis, C. Stem and whole-plant hydraulics in olive (Olea europaea) and kiwifruit (Actinidia deliciosa). Trees 2013, 27, 183–191. [Google Scholar] [CrossRef]
- Maranon-Jimenez, S.; Van den Bulcke, J.; Piayda, A.; Van Acker, J.; Cuntz, M.; Rebmann, C.; Steppe, K. X-ray computed microtomography characterizes the wound effect that causes sap flow underestimation by thermal dissipation sensors. Tree Physiol. 2018, 38, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.D.; Childs, K.W.; King, A.W.; Hanson, P.J. A model of heat transfer in sapwood and implications for sap flux density measurements using thermal dissipation probes. Tree Physiol. 2011, 31, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Moore, G.W.; Bond, B.J.; Jones, J.A.; Meinzer, F.C. Thermal-dissipation sap flow sensors may not yield consistent sap-flux estimates over multiple years. Trees 2010, 24, 165–174. [Google Scholar] [CrossRef]
- Wiedemann, A.; Maranon-Jimenez, S.; Rebmann, C.; Herbst, M.; Cuntz, M. An empirical study of the wound effect on sap flux density measured with thermal dissipation probes. Tree Physiol. 2016, 36, 1471–1484. [Google Scholar] [CrossRef] [Green Version]
- Clearwater, M.J.; Meinzer, F.C.; Andrade, J.L.; Goldstein, G.; Holbrook, N.M. Potential errors in measurement of nonuniform sap flow using heat dissipation probes. Tree Physiol. 1999, 19, 681–687. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Height (m) | Trunk Diameter (cm) | Sapwood Area (cm2) | Sapwood Width (mm) | n |
|---|---|---|---|---|---|
| P. bungeana | 4.4 ± 0.22 | 11.5 ± 0.28 | 58.00 ± 2.54 | 19.3 ± 0.04 | 3 |
| S. matsudana | 5.4 ± 0.12 | 10.7 ± 0.31 | 63.27 ± 1.98 | 24.5 ± 0.06 | 3 |
| Species | Equation | MAE | MBE | RMSE | D | k | ER | R2 | n |
|---|---|---|---|---|---|---|---|---|---|
| P. bungeana | Original | 0.0017 | −0.0017 | 0.0020 | 0.56 | 0.33 | 67 | 0.85 | 25 |
| 10/50 (9) | 0.0012 | −0.0011 | 0.0015 | 0.71 | 0.58 | 42 | 0.78 | 25 | |
| 30/30 (11) | 0.0008 | 0.0003 | 0.0011 | 0.86 | 1.09 | 9 | 0.90 | 25 | |
| S. matsudana | Original | 0.0017 | −0.0017 | 0.0021 | 0.56 | 0.40 | 60 | 0.90 | 25 |
| 10/50 (10) | 0.0008 | 0.0006 | 0.0009 | 0.90 | 1.14 | 14 | 0.96 | 25 | |
| 30/30 (12) | 0.0004 | <0.00001 | 0.0005 | 0.97 | 1.008 | 0.8 | 0.98 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Ma, C.; Zhang, J.; Ma, Y.; Li, C.; Xiong, W. Calibration of Thermal Dissipation Probes for Xylem Sap Flow in the Wood of a Diffuse-Porous and a Conifer Species under Cyclic Heating. Forests 2022, 13, 1964. https://doi.org/10.3390/f13111964
Hu M, Ma C, Zhang J, Ma Y, Li C, Xiong W. Calibration of Thermal Dissipation Probes for Xylem Sap Flow in the Wood of a Diffuse-Porous and a Conifer Species under Cyclic Heating. Forests. 2022; 13(11):1964. https://doi.org/10.3390/f13111964
Chicago/Turabian StyleHu, Meijun, Changming Ma, Jinsong Zhang, Yujie Ma, Chunyou Li, and Wei Xiong. 2022. "Calibration of Thermal Dissipation Probes for Xylem Sap Flow in the Wood of a Diffuse-Porous and a Conifer Species under Cyclic Heating" Forests 13, no. 11: 1964. https://doi.org/10.3390/f13111964