

Genetic Diversity, Structure, and Differentiation of Pinus sylvestris L. Populations in the East European Plain and the Middle Urals
 , , and
, , and 
Abstract
:1. Introduction
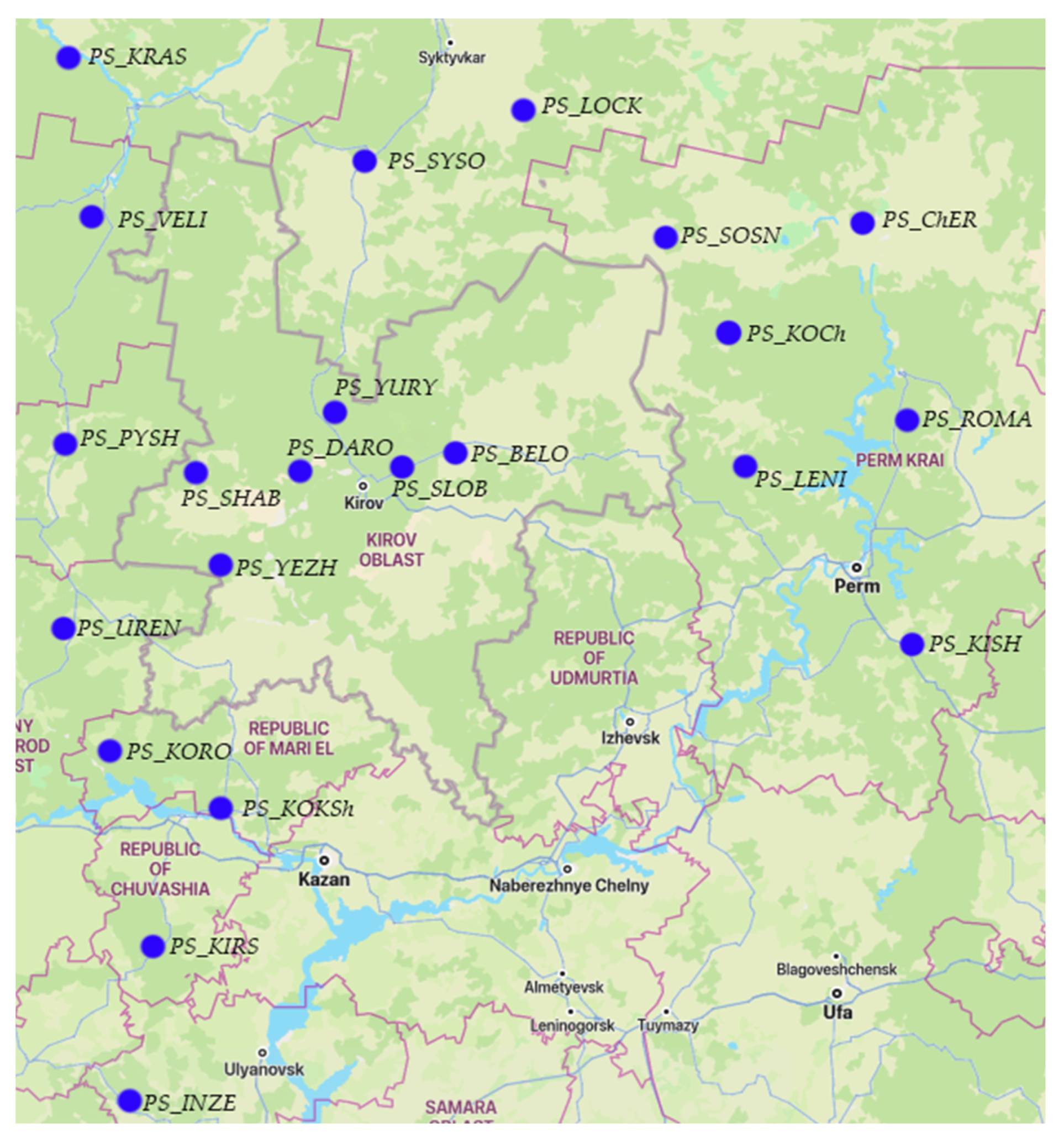
2. Materials and Methods
2.1. Materials
2.2. DNA Extraction and PCR
2.3. Data Analysis
3. Results
3.1. Genetic Diversity of P. sylvestris
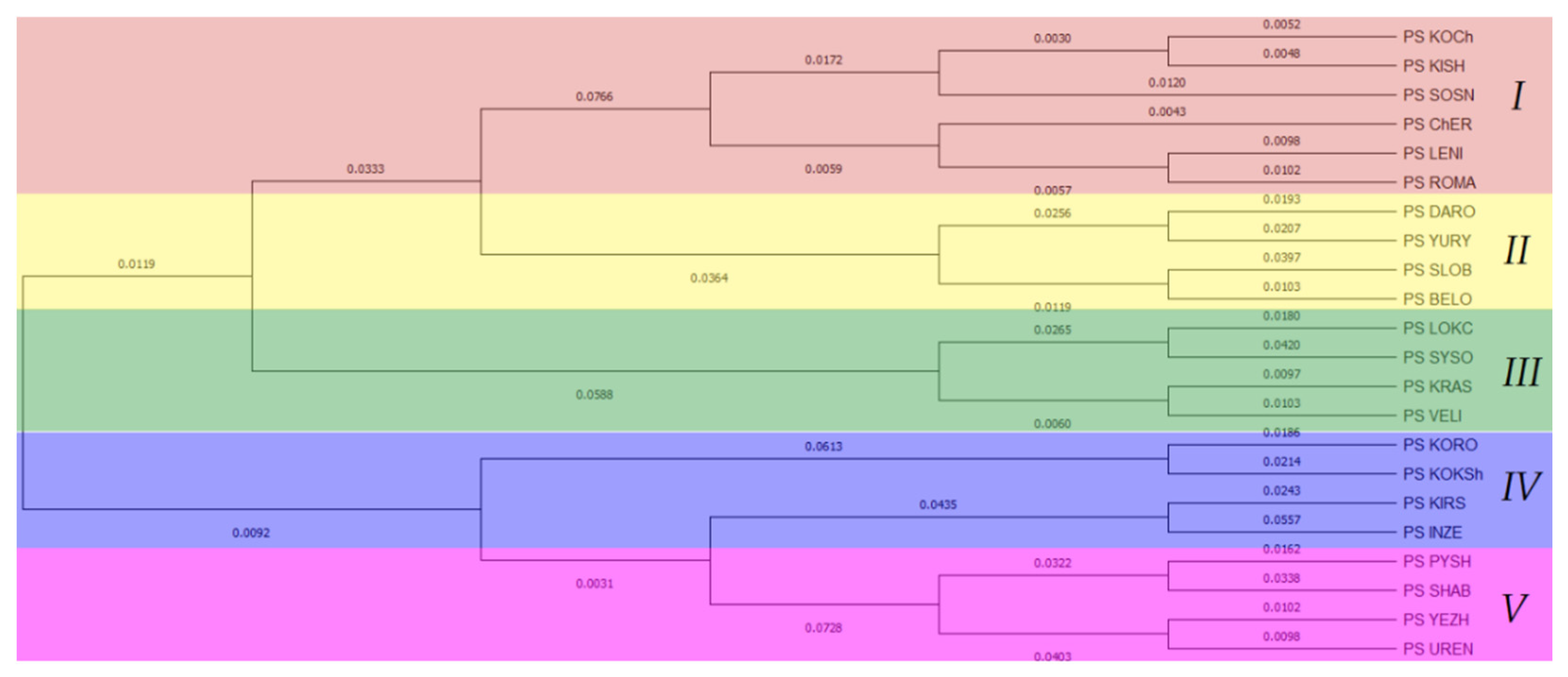
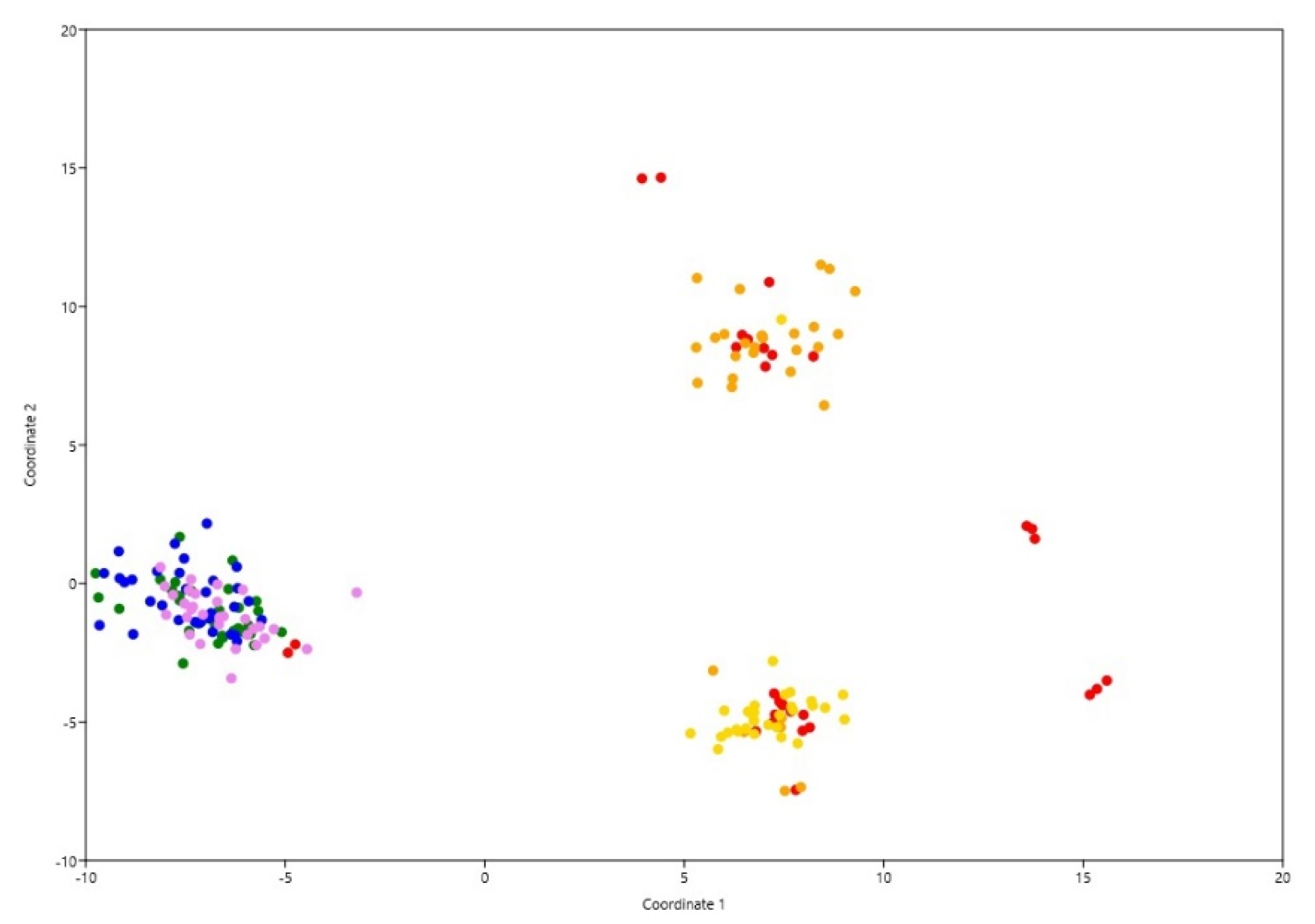
3.2. Population Genetic Structure of P. sylvestris
4. Discussion
4.1. Genetic Diversity of P. sylvestris
4.2. Population Genetic Structure of P. sylvestris
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Frankham, R. How closely does genetic diversity in finite populations conform to predictions of neutral theory? Large deficits in regions of low recombination. Heredity 2012, 108, 167–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spielman, D.; Brook, B.W.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Grady, J.J.; Brook, B.W.; Reed, D.H.; Ballou, J.D.; Tonkyn, D.W.; Frankham, R. Realistic levels of inbreeding depression strongly affect extinction risk in wild populations. Biol. Conserv. 2006, 133, 42–51. [Google Scholar] [CrossRef]
- Potokina, E.K.; Aleksandrova, T.G. Genetic singularity coefficients of common vetch Vicia sativa l. Accessions determined with molecular markers. Russ. J. Genet. 2009, 44, 1309–1316. [Google Scholar] [CrossRef]
- Hoelzel, A.R.; Bruford, M.W.; Fleischer, R.C. Conservation of adaptive potential and functional diversity. Conserv. Genet. 2019, 20, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Razgour, O.; Forester, B.; Taggart, J.B.; Bekaert, M.; Juste, J.; Ibanez, C.; Puechmaille, S.J.; Novella-Fernandez, R.; Alberdi, A.; Manel, S. Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proc. Natl. Acad. Sci. USA 2019, 116, 10418–10423. [Google Scholar] [CrossRef] [Green Version]
- Pazouki, L.; Shanjani, P.S.; Fields, P.D.; Martins, K.; Suhhorutšenko, M.; Viinalass, H.; Niinemets, Ü. Large within-population genetic diversity of the widespread conifer Pinus sylvestris at its soil fertility limit characterized by nuclear and chloroplast microsatellite markers. Eur. J. For. Res. 2015, 135, 161–177. [Google Scholar] [CrossRef]
- Hogberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.; Ekblad, A.; Hogberg, M.N.; Nyberg, G.; Ottosson-Lofvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef]
- Lindén, A.; Heinonsalo, J.; Buchmann, N.; Oinonen, M.; Sonninen, E.; Hilasvuori, E.; Pumpanen, J. Contrasting effects of increased carbon input on boreal som decomposition with and without presence of living root system of Pinus sylvestris L. Plant Soil 2013, 377, 145–158. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Liu, X.; Chen, W.; Liu, Q.; Dai, J. Abietic acid suppresses non-small-cell lung cancer cell growth via blocking ikkbeta/nf-kappab signaling. Onco. Targets Ther. 2019, 12, 4825–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dering, M.; Kosiński, P.; Wyka, T.P.; Pers-Kamczyc, E.; Boratyński, A.; Boratyńska, K.; Reich, P.B.; Romo, A.; Zadworny, M.; Żytkowiak, R.; et al. Tertiary remnants and Holocene colonizers: Genetic structure and phylogeography of Scots pine reveal higher genetic diversity in young boreal than in relict Mediterranean populations and a dual colonization of Fennoscandia. Divers. Distrib. 2017, 23, 540–555. [Google Scholar] [CrossRef] [Green Version]
- Floran, V.; Sestras, R.E.; GarcÍA Gil, M.R. Organelle genetic diversity and phylogeography of Scots pine (Pinus sylvestris L.). Not. Bot. Horti Agrobot. 2011, 39, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Hebda, A.; Wójkiewicz, B.; Wachowiak, W. Genetic characteristics of scots pine in Poland and reference populations based on nuclear and chloroplast microsatellite markers. Silva Fenn. 2017, 51, 1721. [Google Scholar] [CrossRef] [Green Version]
- Tóth, E.G.; Vendramin, G.G.; Bagnoli, F.; Cseke, K.; Höhn, M. High genetic diversity and distinct origin of recently fragmented scots pine (Pinus sylvestris L.) populations along the Carpathians and the Pannonian basin. Tree Genet. Genomes 2017, 13, 47. [Google Scholar] [CrossRef]
- Kavaliauskas, D.; Danusevičius, D.; Baliuckas, V. New insight into genetic structure and diversity of Scots pine (Pinus sylvestris L.) populations in Lithuania based on nuclear, chloroplast and mitochondrial DNA markers. Forests 2022, 13, 1179. [Google Scholar] [CrossRef]
- Chertov, N.; Nechaeva, Y.; Zhulanov, A.; Pystogova, N.; Danilova, M.; Boronnikova, S.; Kalendar, R. Genetic structure of Pinus populations in the Urals. Forests 2022, 13, 1278. [Google Scholar] [CrossRef]
- Vidyakin, A.I.; Semerikov, V.L.; Polezhaeva, M.A.; Dymshakova, O.S. Spread of mitochondrial DNA haplotypes in population of Scots pine (Pinus sylvestris L.) in northern European Russia. Russ. J. Genet. 2012, 48, 1267–1271. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Petrova, I.V. Phylogenogeography and genotaxonomy of Pinus sylvestris L. Populations. Russ. J. Ecol. 2012, 43, 273–280. [Google Scholar] [CrossRef]
- Bouzat, J.L. The population genetic structure of the greater rhea (Rhea americana) in an agricultural landscape. Biol. Conserv. 2001, 99, 277–284. [Google Scholar] [CrossRef]
- Grassi, F.; Imazio, S.; Failla, O.; Scienza, A.; Ocete Rubio, R.; Lopez, M.A.; Sala, F.; Labra, M. Genetic isolation and diffusion of wild grapevine Italian and Spanish populations as estimated by nuclear and chloroplast SSR analysis. Plant Biol. 2003, 5, 608–614. [Google Scholar] [CrossRef]
- Pautasso, M. Geographical genetics and the conservation of forest trees. Perspect. Plant Ecol. Evol. Syst. 2009, 11, 157–189. [Google Scholar] [CrossRef]
- Ribeiro, M.M.; Mariette, S.; Vendramin, G.G.; Szmidt, A.E.; Plomion, C.; Kremer, A. Comparison of genetic diversity estimates within and among populations of maritime pine using chloroplast simple-sequence repeat and amplified fragment length polymorphism data. Mol. Ecol. 2002, 11, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Kalendar, R.; Muterko, A.; Boronnikova, S. Retrotransposable elements: DNA fingerprinting and the assessment of genetic diversity. Methods Mol. Biol. 2021, 2222, 263–286. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Shustov, A.V.; Schulman, A.H. Palindromic sequence-targeted (pst) pcr, version 2: An advanced method for high-throughput targeted gene characterization and transposon display. Front. Plant Sci. 2021, 12, 691940. [Google Scholar] [CrossRef]
- Kalendar, R.; Shustov, A.V.; Seppanen, M.M.; Schulman, A.H.; Stoddard, F.L. Palindromic sequence-targeted (PST) PCR: A rapid and efficient method for high-throughput gene characterization and genome walking. Sci. Rep. 2019, 9, 17707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robledo-Arnuncio, J.J.; Gil, L. Patterns of pollen dispersal in a small population of Pinus sylvestris L. Revealed by total-exclusion paternity analysis. Heredity 2005, 94, 13–22. [Google Scholar] [CrossRef]
- Ganatsas, P.; Tsakaldimi, M.; Thanos, C. Seed and cone diversity and seed germination of Pinus pinea in Strofylia site of the natura 2000 network. Biodivers. Conserv. 2008, 17, 2427–2439. [Google Scholar] [CrossRef]
- Kalendar, R.; Boronnikova, S.; Seppanen, M. Isolation and purification of DNA from complicated biological samples. Methods Mol. Biol. 2021, 2222, 57–67. [Google Scholar] [CrossRef]
- Kalendar, R.; Schulman, A.H. IRAP and REMAP for retrotransposon-based genotyping and fingerprinting. Nat. Protoc. 2006, 1, 2478–2484. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R. A guide to using FASTPCR software for PCR, in silico PCR, and oligonucleotide analysis. Methods Mol. Biol. 2022, 2392, 223–243. [Google Scholar] [CrossRef] [PubMed]
- Vasilyeva, Y.; Chertov, N.; Nechaeva, Y.; Sboeva, Y.; Pystogova, N.; Boronnikova, S.; Kalendar, R. Genetic structure, differentiation and originality of Pinus sylvestris L. Populations in the east of the east European plain. Forests 2021, 12, 999. [Google Scholar] [CrossRef]
- Peakall, R.O.D.; Smouse, P.E. Genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Francis, Y.; Rongcai, Y.; Timothy, B. Popgene. Available online: https://sites.ualberta.ca/~fyeh/popgene_info.html (accessed on 10 October 2022).
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: Chichester, NY, USA, 1987. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega x: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Paul, D.R. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- McInnes, L.; Healy, J.; Saul, N.; Großberger, L. UMAP: Uniform manifold approximation and projection. J. Open Source Softw. 2018, 3, 861. [Google Scholar] [CrossRef]
- Khanova, E.; Konovalov, V.; Timeryanov, A.; Isyanyulova, R.; Rafikova, D. Genetic and selection assessment of the Scots pine (Pinus sylvestris L.) in forest seed orchards. Wood Res. 2020, 65, 283–292. [Google Scholar] [CrossRef]
- Lande, R. Genetics and demography in biological conservation. Science 1988, 241, 1455–1460. [Google Scholar] [CrossRef]
- Vucetich, J.A.; Waite, T.A. Spatial patterns of demography and genetic processes across the species’ range: Null hypotheses for landscape conservation genetics. Conserv. Genet. 2003, 4, 639–645. [Google Scholar] [CrossRef]
- Vidyakin, A.I.; Boronnikova, S.V.; Nechayeva, Y.S.; Pryshnivskaya, Y.V.; Boboshina, I.V. Genetic variation, population structure, and differentiation in Scots pine (Pinus sylvestris L.) from the northeast of the Russian plain as inferred from the molecular genetic analysis data. Russ. J. Genet. 2015, 51, 1213–1220. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Petrova, I.V.; Egorov, E.V.; Sannikova, N.S. Searching for and revealing the system of Pleistocene refugia for the species Pinus sylvestris L. Russ. J. Ecol. 2020, 51, 215–223. [Google Scholar] [CrossRef]
- Sannikov, S.N.; Petrova, I.V.; Egorov, E.V.; Sannikova, N.S. A system of Pleistocene refugia for Pinus sylvestris L. In the southern marginal part of the species range. Russ. J. Ecol. 2014, 45, 167–173. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer ID | Sequence 5′–3′ | Tm (°C) | Ta (°C) * | Total Bands | PIC * |
|---|---|---|---|---|---|
| ISSR-1 ((AC)8T) | ACACACACACACACACT | 55.0 | 56 | 31 | 0.196 |
| CR-212 ((CT)8TG) | CTCTCTCTCTCTCTCTTG | 55.9 | 56 | 43 | 0.260 |
| CR-215 ((CA)6GT) | CACACACACACAGT | 52.6 | 56 | 33 | 0.256 |
| M27 ((GA)8C) | GAGAGAGAGAGAGAGAC | 54.9 | 52 | 33 | 0.261 |
| X10 ((AGC)6C) | AGCAGCAGCAGCAGCAGCC | 72.4 | 64 | 42 | 0.224 |
| Populations | He | ne | I | Populations | He | ne | I |
|---|---|---|---|---|---|---|---|
| PS_KOCh | 0.131 (0.012) | 1.199 (0.020) | 0.213 (0.017) | PS_SHAB | 0.056 (0.010) | 1.092 (0.017) | 0.087 (0.015) |
| PS_SOSN | 0.119 (0.012) | 1.182 (0.020) | 0.194 (0.017) | PS_YEZH | 0.057 (0.010) | 1.092 (0.017) | 0.089 (0.015) |
| PS_KISH | 0.143 (0.013) | 1.228 (0.023) | 0.225 (0.019) | PS_UREN | 0.059 (0.010) | 1.092 (0.016) | 0.092 (0.014) |
| PS_LENI | 0.119 (0.011) | 1.181 (0.020) | 0.195 (0.017) | PS_KORO | 0.122 (0.014) | 1.207 (0.024) | 0.182 (0.020) |
| PS_ChER | 0.142 (0.012) | 1.223 (0.022) | 0.227 (0.018) | PS_KOKSh | 0.117 (0.014) | 1.204 (0.025) | 0.172 (0.020) |
| PS_ROMA | 0.125 (0.012) | 1.194 (0.021) | 0.202 (0.017) | PS_KIRS | 0.121 (0.014) | 1.206 (0.025) | 0.180 (0.020) |
| PS_LOKC | 0.112 (0.013) | 1.187 (0.023) | 0.169 (0.019) | PS_INZE | 0.104 (0.013) | 1.183 (0.025) | 0.152 (0.019) |
| PS_SYSO | 0.106 (0.013) | 1.175 (0.023) | 0.161 (0.019) | PS_DARO | 0.164 (0.014) | 1.268 (0.025) | 0.249 (0.020) |
| PS_KRAS | 0.129 (0.014) | 1.216 (0.024) | 0.195 (0.020) | PS_YURY | 0.176 (0.015) | 1.299 (0.027) | 0.263 (0.021) |
| PS_VELI | 0.118 (0.013) | 1.193 (0.023) | 0.180 (0.019) | PS_SLOB | 0.151 (0.014) | 1.249 (0.025) | 0.230 (0.020) |
| PS_PYSH | 0.063 (0.011) | 1.106 (0.019) | 0.096 (0.016) | PS_BELO | 0.177 (0.015) | 1.303 (0.027) | 0.264 (0.021) |
| Total | 0.119 (0.003) | 1.195 (0.005) | 0.183 (0.004) | Total | 0.119 (0.003) | 1.195 (0.005) | 0.183 (0.004) |
| Subdivision Indicator | df | SS | MS | Dispersion | % | p |
|---|---|---|---|---|---|---|
| Among groups | 1 | 3442.465 | 3442.465 | 10.041 | 28% | <0.001 |
| Among populations | 20 | 11379.929 | 568.996 | 13.168 | 37% | <0.001 |
| Within populations | 900 | 11373.549 | 12.637 | 12.637 | 35% | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sboeva, Y.; Chertov, N.; Nechaeva, Y.; Valeeva, A.; Boronnikova, S.; Kalendar, R. Genetic Diversity, Structure, and Differentiation of Pinus sylvestris L. Populations in the East European Plain and the Middle Urals. Forests 2022, 13, 1798. https://doi.org/10.3390/f13111798
Sboeva Y, Chertov N, Nechaeva Y, Valeeva A, Boronnikova S, Kalendar R. Genetic Diversity, Structure, and Differentiation of Pinus sylvestris L. Populations in the East European Plain and the Middle Urals. Forests. 2022; 13(11):1798. https://doi.org/10.3390/f13111798
Chicago/Turabian StyleSboeva, Yana, Nikita Chertov, Yulia Nechaeva, Alena Valeeva, Svetlana Boronnikova, and Ruslan Kalendar. 2022. "Genetic Diversity, Structure, and Differentiation of Pinus sylvestris L. Populations in the East European Plain and the Middle Urals" Forests 13, no. 11: 1798. https://doi.org/10.3390/f13111798