1. Introduction
The mechanisms of species coexistence have long been a research hotspot in community ecology. Classical ecological theory states that stable coexistence between competing species requires them to occupy different niches. Competing species will diverge, leading to a reduction in niche overlap. However, high levels of species diversity in tropical and sub-tropical forests cannot always be explained by variations in species’ characteristics, indicating that there may be niche differences between plants that have been overlooked [
1]. Coincidentally, characteristics related to patterns of aboveground vertical variation in species’ performances have been less frequently explored. Although several researchers have proposed that functional trait variation in the vertical dimension can help elucidate the mechanisms underlying species coexistence in communities [
2,
3,
4,
5], more research is needed to explore the significance of aboveground vertical differentiation to achieve a deeper understanding of community assembly rules. The vertical variation of key plant traits in response to limited resources should be especially considered.
Leaf functional traits play an important role in leaf and plant functioning and are associated with species’ strategies of resource acquisition and use [
6,
7]. Photosynthetic traits and related traits are key features, and their variation is important for the study of species coexistence [
8]. Shade tolerance is one of the most important traits for understanding community assembly and forest dynamics [
9]. The rate of leaf dark respiration (R
d) is supposed to be a good estimator of the shade tolerance of tree species [
10]. Chlorophyll (Chl) fluorescence acts as an indicator of energy conversion of photosynthetic organisms and has been a powerful tool to study photosynthesis performance [
11]. The maximum relative electron transport rate (
rETRmax), initial slope (
α), photoinhibition coefficient (
β), and half saturating irradiance (
Ik) are Chl fluorescence parameters derived from rapid light curves (RLCs; relative electron transport rate versus photosynthetic photon flux density (PPFD) curve), which reflect the expressed intrinsic capacity of photosynthesis [
12].
rETRmax determines the photosynthetic capacity, while
α reflects the light utilisation efficiency of the leaf. Assessments of
Ik represent the tolerance of the leaf to high light [
13]. Leaf dry mass (LDM) is the basic calculation index that represents the physiological activities of plant leaves and an important reference variable to study the competition pattern of forest plant species [
7]. Leaf area (LA) reflects the ability of leaves to capture light and perform photosynthesis and is also related to the energy and water balance of leaves [
14]. The specific leaf area (SLA) is associated with the leaf economic spectrum and the metabolic and growth rates of the species [
7]. Leaf dry matter content (LDMC) is correlated with the average density of the leaf tissue, potential RGR, and water balance of the leaf [
15]. Leaf thickness (LT) is associated with photosynthesis and growth rate [
16].
Tree crown architecture characteristics are important for the exploitation of the vertical height resource gradient. The crown depth (
Dc) of a tree has been associated with the capability for capture and utilisation of light since a deeper crown largely means that the crown covers a more complex light environment and consequently has a higher variability of traits in the vertical dimension, and vice versa. Crown breadth (
Bc) and crown volume (
Vc) can reflect the pattern of competition for space between a tree and its neighbours.
Bc and
Vc are also closely related to the total number of leaves in a tree and thereby the photosynthetic capacity of the tree, which in turn affects the growth level and competition status of the tree in the stand [
17,
18].
In a vertically well-developed, mature forest, species tend to develop different heights, which is an important manifestation of the spatial segregation of tree crowns along the vertical gradient of resource availability. Species with different heights should be characterised by different survival strategies, and variations in key traits should be found in the vertical direction of the community [
19,
20]. Moreover, photosynthetic performance plasticity within an individual crown in the vertical dimension is also believed to allow species to diverge along the vertical gradient of light availability within the canopy and contribute to a stable coexistence among neighbours [
3,
20].
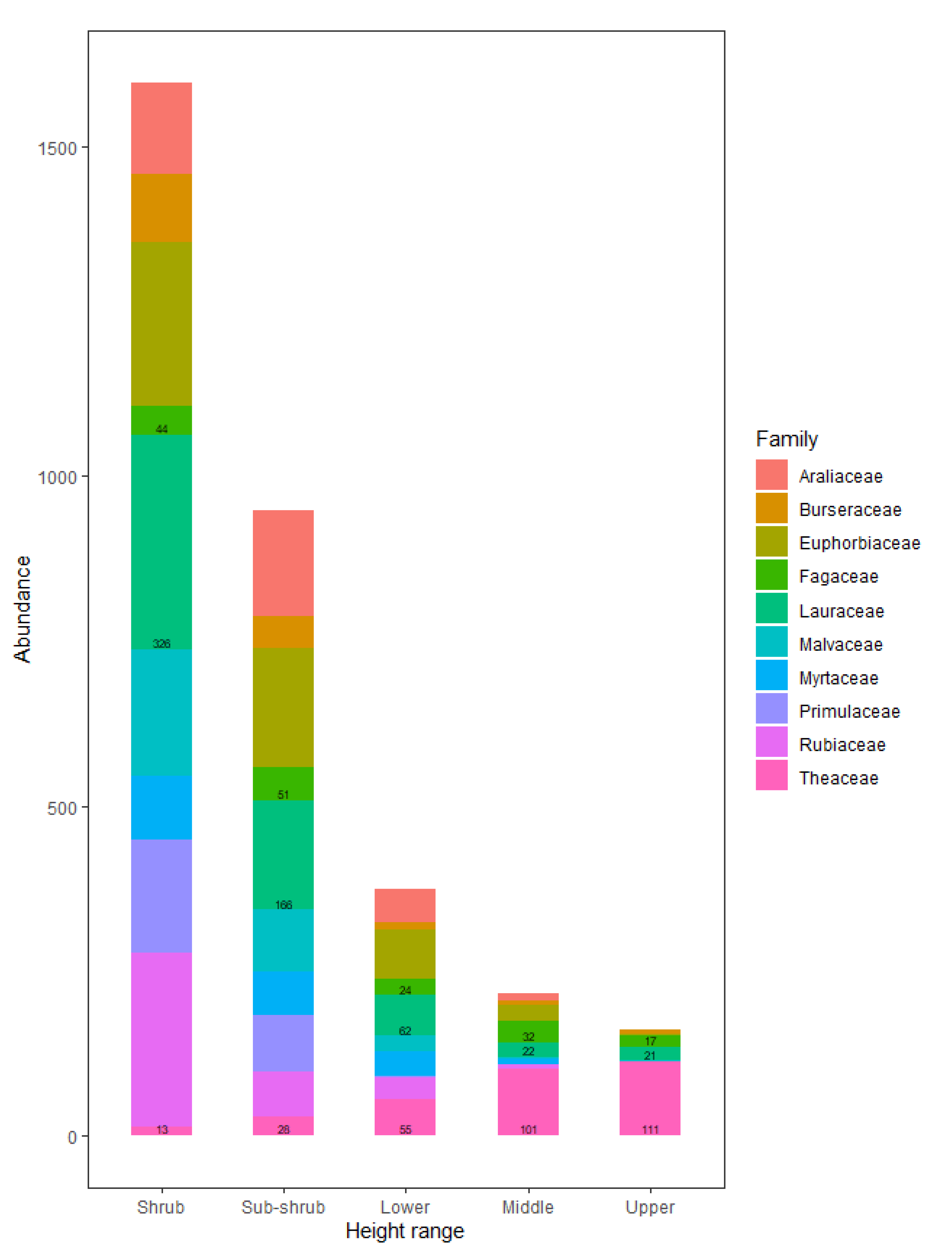
The divergence of comparative advantages between species pairs in different vertical spaces may be an important factor that leads to complementary resource utilisation and subsequently enhanced species coexistence. One species might be found to exhibit superior performance within a specific vertical range, producing a relative advantage over other species, while it might be average or even inferior in performance to other species within another vertical range. Furthermore, intuitive and persuasive evidence to verify the existence of vertical niche differentiation is that species with different strategies prefer different vertical spaces, which was revealed by differences in abundance across different height ranges [
21]. Numerous studies have found that the density of tree species differs across environmental gradients [
22,
23,
24]. Nevertheless, few studies have examined the role of vertical variation in the ability to utilise resources in the context of promoting plant coexistence.
To explore the patterns of species coexistence and enrich niche differentiation theory, we aimed to confirm the vertical niche differentiation of tree species in a natural subtropical forest by detecting variation in important leaf traits and crown structure characteristics within the vertical structure of trees. To this end, we examined vertical variation:
Between height groups (lower height group (LG), middle height group (MG), and higher height group (HG)) and within the same height range (shrub canopy (1.4–4 m), sub-shrub canopy (4–6.8 m), lower canopy (6.8–11.3 m), middle canopy (11.3–17.4 m), and upper canopy (17.4–27.1 m)) of different species;
In the same species across tree height;
At different parts of the tree crown (lower part (LP), middle part (MP), and upper part (UP)).
We also present the result of variation in different parts of the tree crown in the five height ranges and in each target species. The differences in traits and abundance of species across the five height ranges were also investigated to determine if there was complementarity among plants in the vertical dimension and if the existence of niche differentiation between species along the vertical gradient was substantiated.
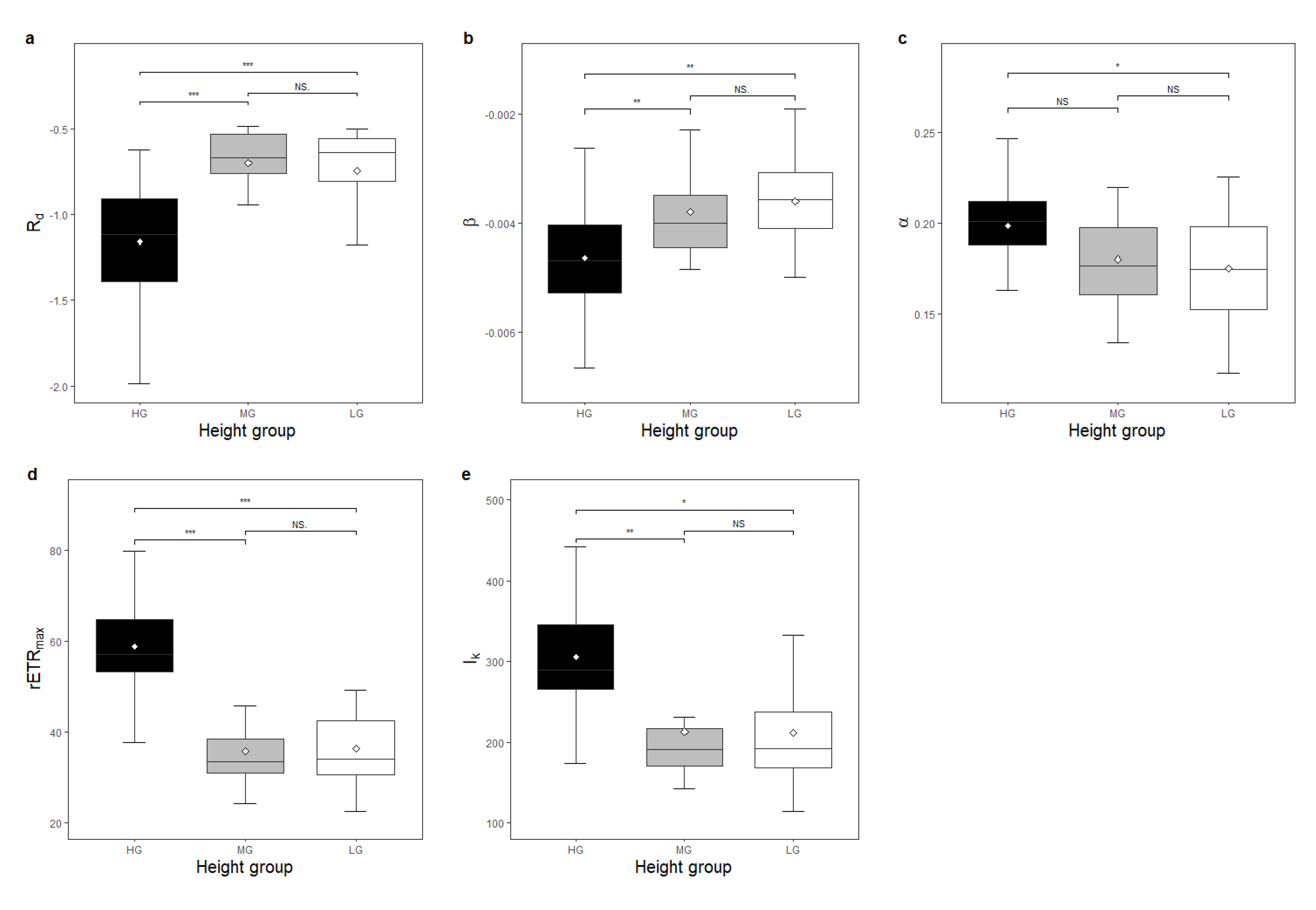
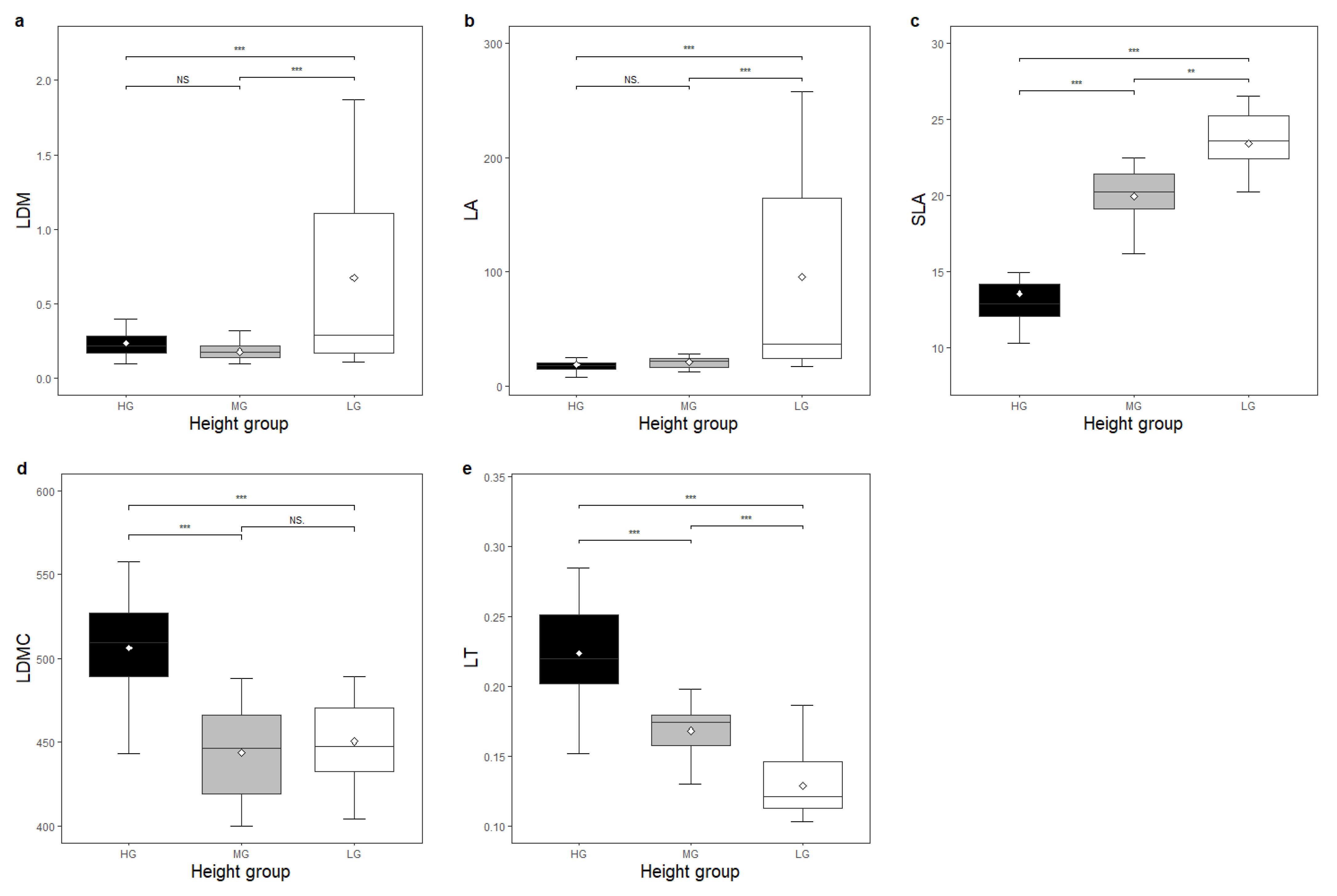
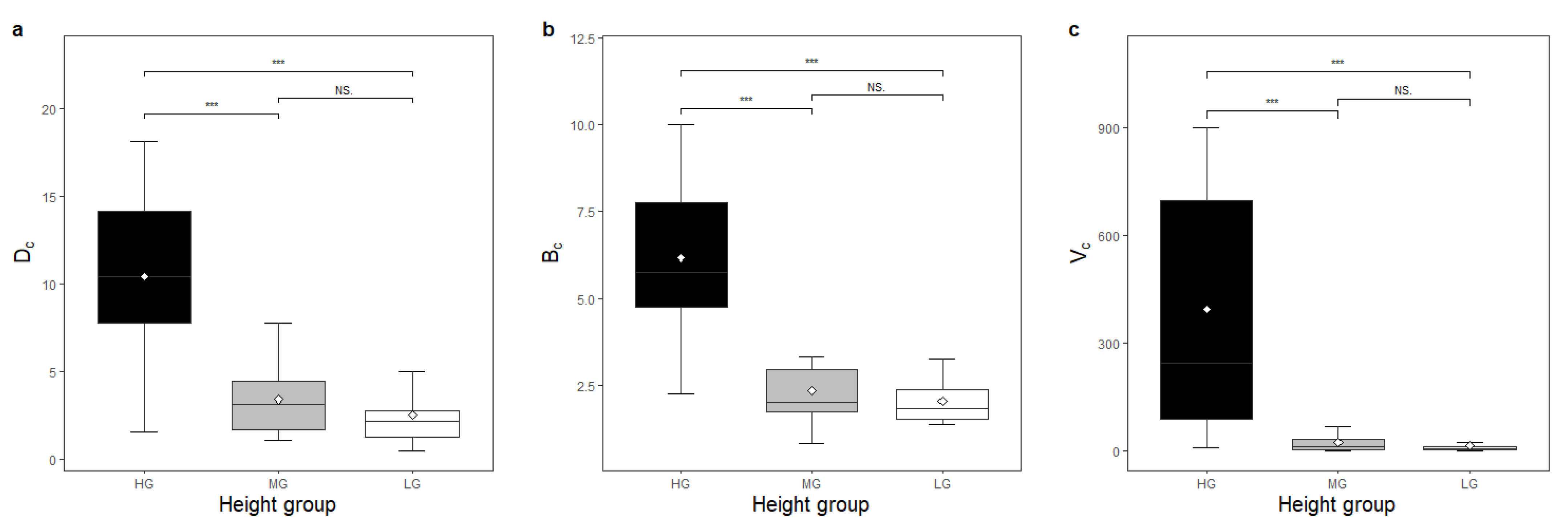
3. Results
Functional traits showed significant differences between species height groups (
Table A3,
Figure 2,
Figure 3 and
Figure 4). The R
d, LDM, LA, SLA, and
β of LG species was significantly higher than that of species in HG (
Figure 2a,b and
Figure 3a–c). The R
d, SLA, and
β of MG species was significantly higher than that of HG species (
Figure 2a,b and
Figure 3c). The LDM, LA, and SLA of LG species was significantly higher than that of MG species (
Figure 3a–c). The
α,
rETRmax,
Ik, LDMC, LT,
Dc,
Bc, and
Vc of LG species was lower than that of species in HG (
Figure 2c–e,
Figure 3d,e and
Figure 4). The
rETRmax,
Ik, LDMC, LT,
Dc,
Bc, and
Vc of MG species was lower than that of HG species (
Figure 2d,e,
Figure 3d,e and
Figure 4). The LT of LG species was lower than that of MG species (
Figure 3e). Tree height showed significant differences in each combination of height groups (
p < 0.05). R
d, LDM, LA, SLA, and
β were negatively related to tree height, while
rETRmax,
Ik, LDMC, LT,
Dc,
Bc, and
Vc were positively related to tree height (
p < 0.05).
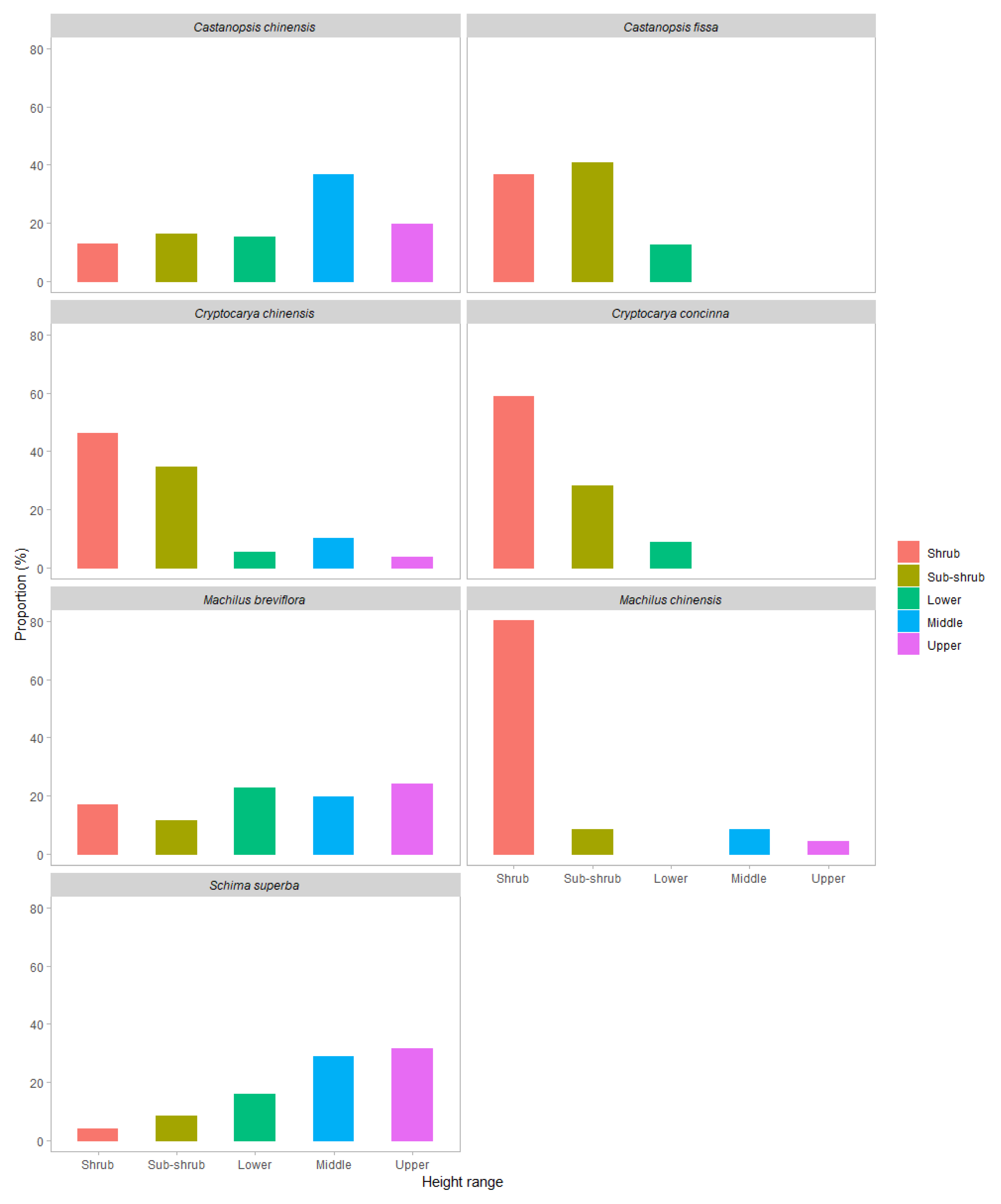
For species pairs co-existing in the same height range, species differences in traits were not consistent across different height ranges (
Table A3 and
Table A4). For instance, the SLA and
Dc of
Cr.
chinensis was only lower than that of Ca.
fissa in the shrub canopy, and the LT of
Cr.
chinensis was higher than that of Ca.
fissa in the shrub and sub-shrub canopies but not in the lower canopy. The LT of
Cr.
concinna was higher than that of Ca.
fissa in the sub-shrub canopy but not in the other height ranges.
Cryptocarya concinna had a significantly lower
β than
Cr.
chinensis in the lower canopy.
Machilus breviflora had a lower R
d and LDM than Ca.
chinensis in the upper canopy. The LDM and LT of
S.
superba was only lower than that of Ca.
chinensis in the upper canopy.
Schima superba had a significantly higher R
d, LDM, LA, and
α than that of
M.
breviflora in the upper canopy but not in the middle canopy. The
β of
M.
breviflora was significantly lower than that of
Cr.
chinensis in the lower canopy.
Machilus chinensis only had a higher SLA and lower LT than
Cr.
chinensis in the shrub canopy (
Table A4).
Intra-specific variabilities across tree height were found in our study (
Table A5). There were intra-specific variations in at least one trait for each species.
Castanopsis fissa,
Cr. chinensis and
M. chinensis showed intra-specific variations in relative more traits.
Species differences in
rETRmax,
Ik, LDMC, LT,
Dc,
Bc, and
Vc were positively related with differences in species abundance in higher height ranges (mainly in the middle and upper canopies) but inversely associated with differences in abundance in lower ranges (in the shrub and sub-shrub canopies) (
Table 3). Species differences in SLA and
β showed a positive correlation with species differences in abundance in lower height ranges and a negative correlation with species differences in abundance in higher ranges. Species differences in R
d were positively correlated with species differences in abundance in the shrub and sub-shrub canopies.
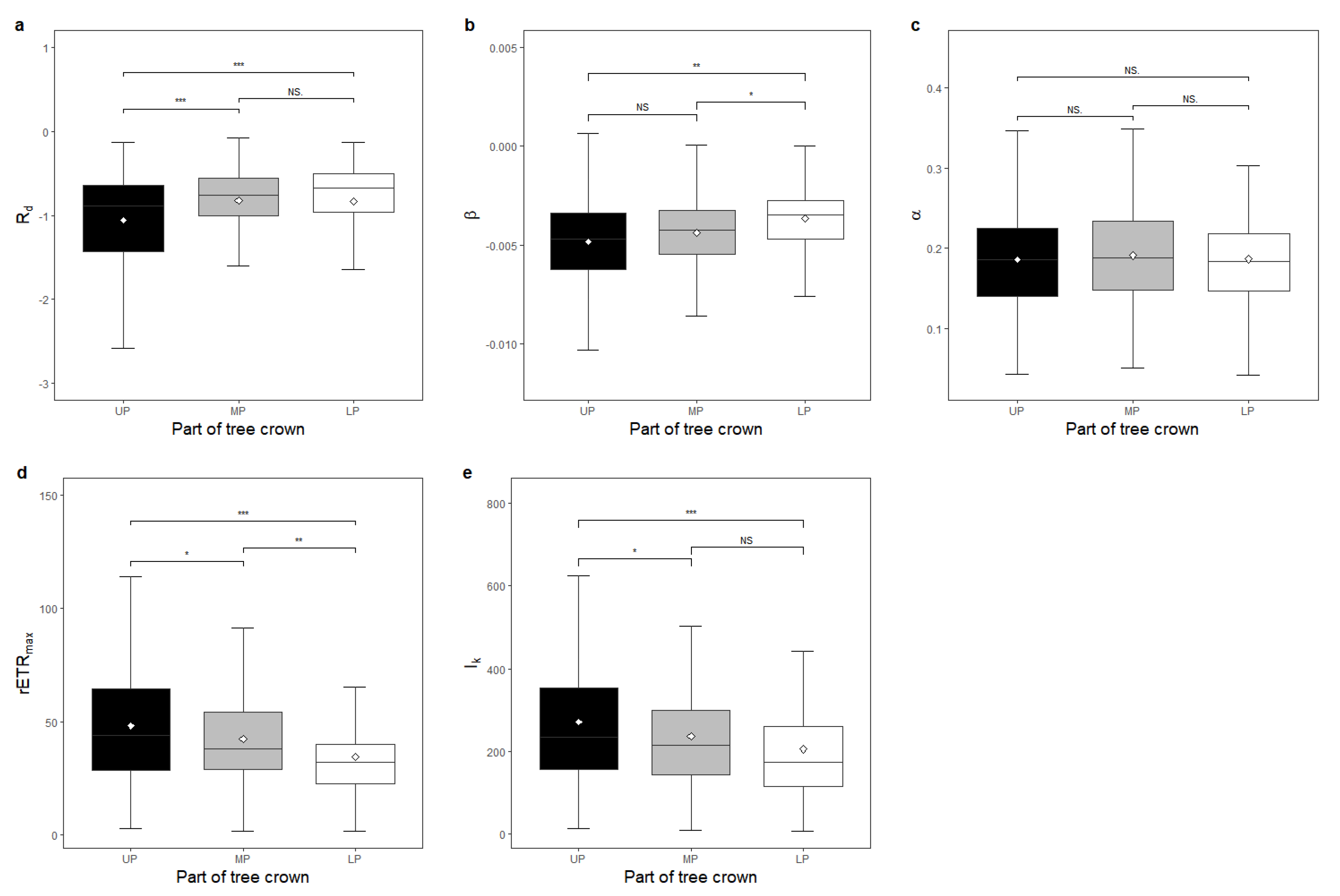
The results regarding different parts of the tree crown for all species are shown in
Table A6 and
Figure 5. The
β of LP was significantly higher than that of MP (
Figure 5b), while
rETRmax showed the opposite trend (
Figure 5d). The R
d and
β of LP was higher than that of UP (
Figure 5a,b), but
rETRmax and
Ik showed the opposite trend (
Figure 5d,e). The R
d of MP was higher than that of UP (
Figure 5a), but
rETRmax and
Ik showed the opposite trend (
Figure 5d,e).
With respect to individual species, significant differences of R
d between different parts of the tree crown were found in Ca.
chinensis,
Cr.
Concinna, and
S.
superba (
Table A6). Differences in
rETRmax were observed in Ca.
chinensis and
S.
superba. Differences in
β were found in Ca.
chinensis and
M.
chinensis. The difference in
α was found in
M.
breviflora, while differences in
Ik were found in Ca.
chinensis (
Table A6).
For each height range scenario, a difference in R
d between parts of the tree crown appeared in the sub-shrub, middle, and upper canopies of the forest. Differences in
β were observed in the lower and middle canopies. Differences in
rETRmax were found in the shrub, middle, and upper canopies. Differences in
Ik were found in the shrub and upper canopies (
Table A7). Intraspecific variabilities of photosynthetic traits were found among different parts of crown in all target species except Ca.
fissa and
Cr. Chinensis (
Table A8).
4. Discussion
The main results of the present study support the hypothesis that there is vertical niche differentiation among tree species in subtropical forests. Significant differences in functional traits between different height groups (
Table A3,
Figure 2) confirmed the existence of niche differentiation in the vertical dimension of the forest. In the present study, the R
d of HG species was found to be significantly lower than that of LG and MG species (
Figure 2a), demonstrating that there was significant vertical variation in shade tolerance among the species in these groups, as R
d is often used as a reliable surrogate measuring tree species’ shade tolerance [
10]. Species occupying the top layer of the forest canopy tended to be less shade tolerant compared to species at lower strata, which was consistent with previous studies [
5,
23]). The conclusion was further confirmed by the result that R
d was negatively related to tree height in the present study. Species gain competitive advantages through prior access to light by being taller than their neighbours, which requires more investment in non-productive structures and physiological activity that sustains essential consumption [
27], also resulting in a lower R
d.
The species in the lower height group had a higher LDM, LA, SLA, and
β than those in the higher group, but a lower
α,
rETRmax,
Ik, LDMC, LT,
Dc,
Bc, and
Vc (
Figure 2,
Figure 3 and
Figure 4). The relationship between these traits and tree height supported these results. This confirmed that leaves were larger in species occupying habitats with low light. Larger leaves are conducive for adapting to low-light conditions by increasing light interception. Moreover, the pattern that low-light species have larger leaves and high-light species have smaller leaves satisfied the demand for increasing the efficiency of water utilisation, which was the ratio of CO
2 uptake to water loss, ensuring the leaf energy and water balance in each light environment [
14,
28]. As a mass-based area of the leaf, SLA was more sensitive to showing differences between different height groups (
Figure 3c). Numerous studies have found that SLA decreases from the bottom of the canopy to its top [
29,
30,
31]. These variations are considered either an adaptation to a varying light environment, allowing the optimisation of carbon gains, or as a result of increased leaf water stress [
32]. Moreover, species at higher height ranges had thicker and denser leaves, regardless of the leaf area (
Figure 3e,d), which may also contribute to resisting low water availability and maintaining a higher modulus of elasticity and greater resistance to cell wall collapse under extreme water deficit conditions [
33].
Leaves in HG species had a higher intrinsic capacity of photosynthesis, capacity of high-light tolerance, photochemical reaction initiation rate, and light utilisation efficiency than in MG and LG species (
Figure 2c–e). These results were consistent with previous studies [
34,
35]. In contrast to our expectation, the
β of HG species was lower than that of LG and MG, demonstrating that HG species were more likely to decrease their photosynthetic potential when light intensity exceeded their saturation point than those at a lower height. One possible explanation for this paradox could be that in different light intensity ranges, the effect of the same amount of light intensity increases in the two groups of plants was not equal. Photoinhibition occurred at a higher light intensity level in HG species than in the other groups of species (
Figure 2e), and the light intensity that triggered photoinhibition in HG species was closer to the threshold for the plant to cease physiological activities. An increase in excess light of one unit in the high-light range is more potentially harmful to plants than that in the low-light range, and it will inevitably cause more intense adverse factors to the HG species. As a result, HG species have evolved more rapid responses to adapt to excess light and to protect plant tissues (or have simply suffered damage due to exposure to excess high light), resulting in a rapid decline in photosynthetic function.
HG species tend to have a larger crown size than LG and MG species, either in crown depth, breadth, or crown volume (
Figure 4). It is quite conceivable that tall trees have a larger respiration load, and they could compensate for this by making larger and deeper crowns with a larger amount of photosynthesising leaf area. Compared to a larger LA and SLA in shorter species, a larger tree crown in HG species is likely an alternative to ensure sufficient area to capture enough light energy since the size of the tree crown is closely related to the total number of leaves on a tree [
17,
18]. The photosynthetic capacity of the tree in HG and, consequently, the growth level and competition status of the tree in the stand can be ensured.
For species pairs in which tree crowns coexist in the same height range, trait differences between them were not consistent across different height groups (
Table A3 and
Table A4 ). Some significant differences in traits between species were only found in particular height ranges, even though those species grew together in multiple height ranges. For example, significant differences in SLA and
Dc between
Cr.
chinensis and Ca.
fissa were found only in the shrub canopy, while no significant difference between these species was found in the sub-shrub and lower canopies, although the crowns of both species were present in the shrub, sub-shrub, and lower canopies. Significant differences in R
d, LDM, LA, and
α between
S.
superba and
M.
breviflora were only shown in the upper canopy, although the crowns of both species occurred in the middle and upper canopies (
Table A4). These results demonstrated that some species performed better or worse than their competitors in specific height ranges in certain abilities, while showing no significant difference from their competitors in other height ranges, thereby demonstrating that there was variation in comparative advantages between species across different height ranges. For example, Ca.
fissa was superior in terms of enlarging the light-capturing area by increasing the area of each blade per mass and the total light-exposed area of the crown in the vertical dimension, which manifested as a larger SLA and
Dc, than
Cr.
chinensis in the shrub canopy. However, the SLA and
Dc between the two species were not different in the sub-shrub and lower canopies, which generally had a higher light level than the shrub canopy. This suggested that Ca.
fissa was more adaptive to a relative low-light environment and more competitive at a lower height range than
Cr.
chinensis. Although it has rarely been considered as the object of research regarding niche segregation, this kind of variation is an important factor that leads to complementary resource use, subsequently enhancing species coexistence. It also suggests that an imprecise or even misleading conclusion might be proposed without considering the vertical variation when comparing species differences.
Trait differences were also found in the same species across tree height (
Table A5). At least one trait was found with intra-specific variability for each species. It is understandable that as trees grow taller, the life history of species varies, and the environment to which they respond changes, and with that comes variation in traits. The results that R
d, LDM, LA, SLA, and
β were negatively related to tree height while
rETRmax,
Ik, LDMC, LT,
Dc,
Bc, and
Vc were positively related to tree height show the general trend of trait variation of all species as tree height varies. For certain species, however, it varies according to the ecological strategies of the species. For instance, intra-specific variances of crown breadth were not found in Ca.
chinensis, Ca.
fissa,
Cr. concinna, and
M.
breviflora in the study. It is interesting that there were more traits that showed intra-specific variations in Ca.
fissa,
Cr. chinensis, and
M. chinensis, which were all mainly distributed in the shrub and sub-shrub canopy in our study plot. This can be coupled with the fact that traits with intra-specific variability of these species were closely related to the acquisition of light resources. This may, therefore, hint at strong intraspecific competition for light resources in these species in the low-light environment. On the other hand, intraspecific variabilities of traits were also shown among different parts of crown (
Table A8), indicating that the vertical structure of a tree also contributes to the vertical niche differentiation of the plant.
Contrasting results between lower height ranges and higher ranges in the relationship between species’ differences in functional traits and species difference of abundance (
Table 3) demonstrated that there was obvious vertical niche separation between species in the community. Species with a stronger shade tolerance, leaf light-capturing efficiency, and a lower leaf photosynthetic capability, high-light tolerance, leaf dry matter content, leaf thickness, and crown size were more abundant in lower height ranges, and vice versa.
Variations in functional traits were also found in different positions within the crown of each tree in the vertical dimension (
Table A6). Shade tolerance in the top part of the tree was lower than that in the interior overall (
Figure 5a). This result revealed that the top layer of the crown featured a weaker ability to endure a low-light environment. Meanwhile, the UP had greater capacity to handle high-light conditions (
Figure 5e) and a higher photosynthetic potential (
Figure 5d). Leaves in LP had a higher
β than those of MP and UP, demonstrating a smaller rate of decline in the photosynthetic capacity when the light intensity exceeded its saturation point, which might be due to the increase by the same unit of light in the relatively lower light range being less destructive. Consistent with previous research [
36,
37], our results confirmed that photosynthetic performance in different parts of the crown differed across the vertical dimension.
A more detailed analysis showed that Ca.
chinensis and
S.
superba were more responsible for functional trait variations between parts of the tree crown in all species scenarios since most of the differences in shade tolerance and photosynthetic capacity between parts of the tree crown were found in these species (
Table A6 and
Table A8). In the study plot,
S.
superba, and Ca.
chinensis were common species, as the 4th and 17th most abundant of all species, respectively. The vertical variation in shade tolerance and photosynthetic potential within an individual crown may have contributed to the dominance of these species in the light-limited community. In each height range scenario, variations in photosynthetic characteristics mostly occurred in the middle and upper canopies of the forest (
Table A7). Differences in traits among parts of the tree crown preformed differently across height ranges, indicating that a change in perspective is required when interpreting niche differentiation patterns between species, and changes in canopy height layer should be considered.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




