
Differences in Transpiration Characteristics among Eucalyptus Plantations of Three Species on the Leizhou Peninsula, Southern China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plantation
2.2. Measurement of Environmental Factor Parameters
2.3. Leaf Area Index (LAI) Measurement
2.4. Sap Flow Measurements
2.5. Sapwood Area Determination
2.6. Estimation Methods for Individual Tree and Stand-Level Transpiration
2.7. Data Analysis and Statistics
3. Results
3.1. Stands Structure and the Relationship between Sapwood Area and DBH
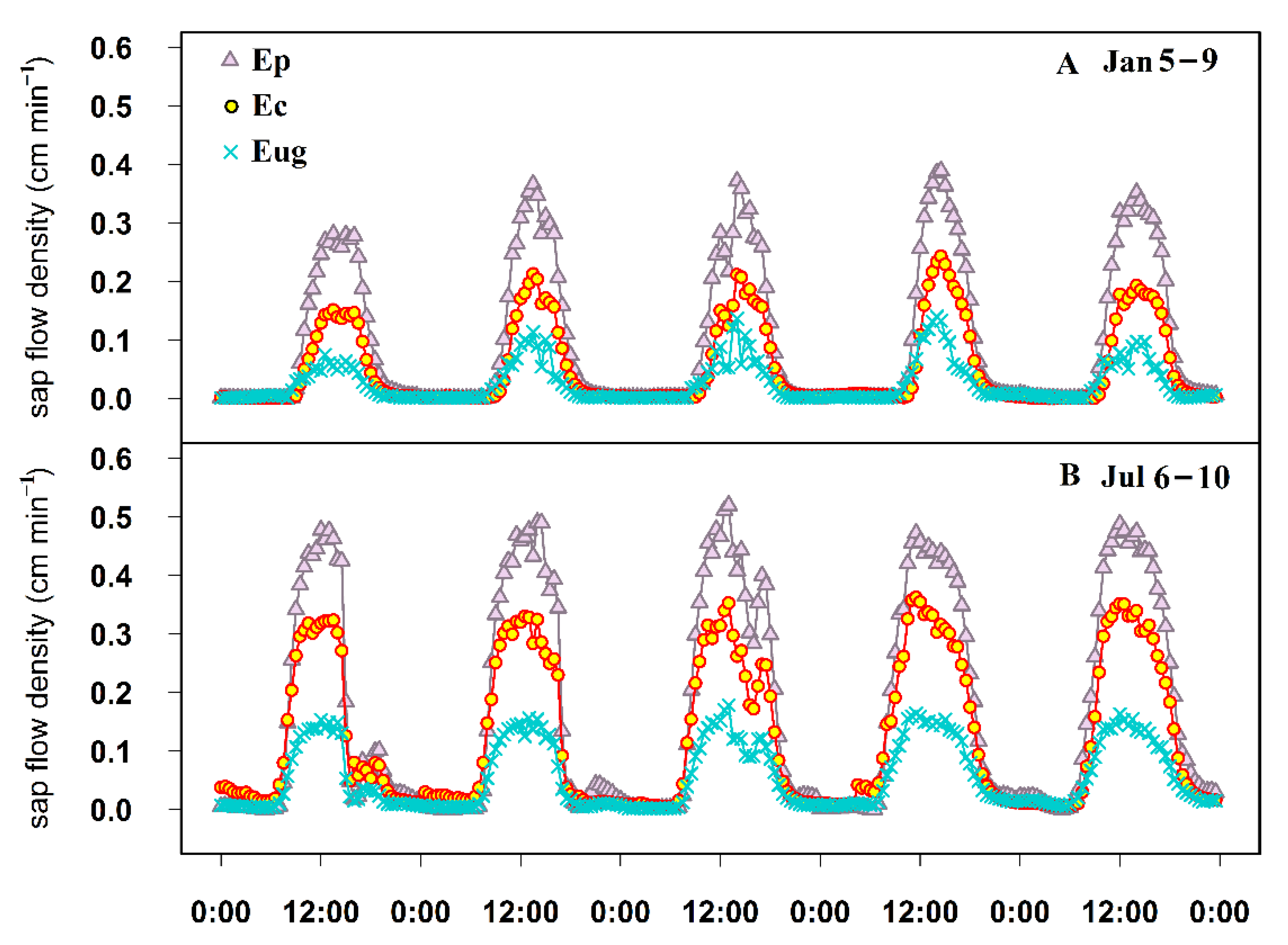
3.2. Characteristics of Sap Flow Density of the Three Eucalyptus Species
3.3. The Water Use Characteristics of the Three Species of Eucalyptus Stands
3.4. Factors Influencing Transpiration by the Three Eucalyptus Species
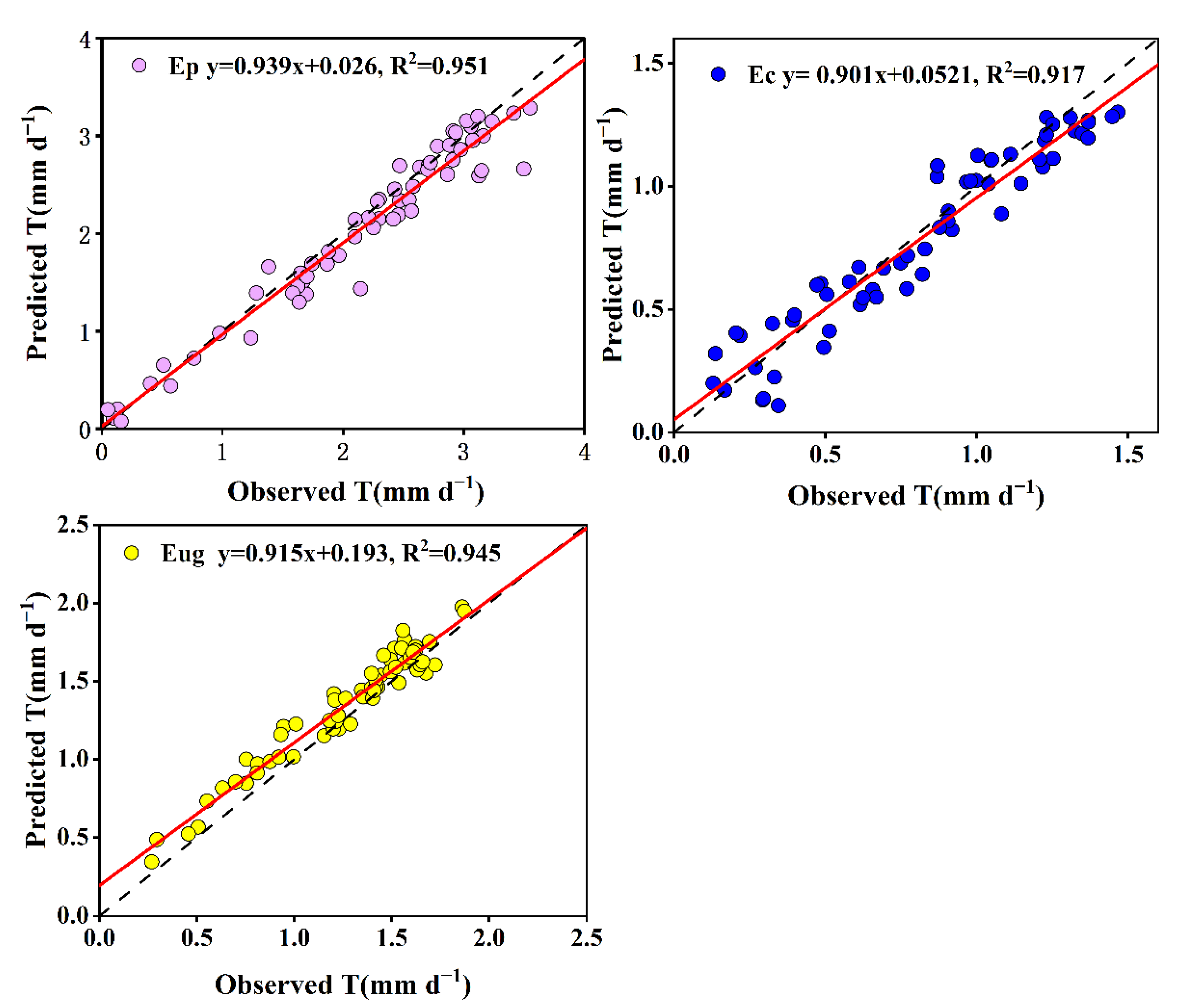
3.5. Model to Estimate the Daily T of Each Eucalyptus Species
4. Discussion
4.1. Sap Flow Density and Water Use of the Three Eucalyptus Species
4.2. Factors Influencing Water Use by the Three Eucalyptus Species
4.3. Daily Stand Transpiration (T) Predictive Models for the Three Eucalyptus Species
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Forestry and Grassland Administration. 2014–2018 China Forest Resources Report; China Forestry Publishing House: Beijing, China, 2019. [Google Scholar]
- Morris, J.; Ningnan, Z.; Zengjiang, Y.; Collopy, J.; Daping, X. Water use by fast-growing Eucalyptus urophylla plantations in southern China. Tree Physiol. 2004, 24, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Zhao, P.; Zhou, G.; Zhu, L.; Huang, Y.; Zhao, X.; Ni, G. Stand-scale transpiration of a Eucalyptus urophylla × Eucalyptus grandis plantation and its potential hydrological implication. Ecohydrology 2018, 11, e1938. [Google Scholar] [CrossRef]
- Cabral, O.M.R.; Rocha, H.R.; Gash, J.H.C.; Ligo, M.A.V.; Freitas, H.C.; Tatsch, J.D. The energy and water balance of a Eucalyptus plantation in southeast Brazil. J. Hydrol. 2010, 388, 208–216. [Google Scholar] [CrossRef]
- Jackson, R.B.; Jobbágy, E.G.; Avissar, R.; Roy, S.B.; Barrett, D.J.; Cook, C.W.; Farley, K.A.; Maitre, D.C.L.; McCarl, B.A.; Murray, B.C. Trading Water for Carbon with Biological Carbon Sequestrationy. Science 2005, 310, 1944–1947. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.M.; Hewlett, J.D. A review of catchment experiments to determine the effects of vegetation changes on water yield and evapotranspiration. J. Hydrol. 1982, 55, 3–23. [Google Scholar] [CrossRef]
- Scott, D.F.; Lesch, W. Streamflow responses to afforestation with Eucalyptus grandis and Pinus patula and to felling in the Mokobulaan experimental catchments, South Africa. J. Hydrol. 1997, 199, 360–377. [Google Scholar] [CrossRef]
- Buckley, T.N.; Turnbull, T.L.; Pfautsch, S.; Gharun, M.; Adams, M.A. Differences in water use between mature and post-fire regrowth stands of subalpine Eucalyptus delegatensis R. Baker. For. Ecol. Manag. 2012, 270, 1–10. [Google Scholar] [CrossRef]
- Lane, P.N.J.; Morris, J.; Zhang, N.; Zhou, G.; Zhou, G.; Xu, D. Water balance of tropical eucalypt plantations in south-eastern China. Agric. For. Meteorol. 2004, 124, 253–267. [Google Scholar] [CrossRef]
- Soares, J.V.; Almeida, A.C. Modeling the water balance and soil water fluxes in a fast growing Eucalyptus plantation in Brazil. J. Hydrol. 2001, 253, 130–147. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiao, K.; Zhao, Z.; Xiang, W.; Xu, C.; Lei, P.; Deng, X.; Li, J. Stand transpiration estimates from recalibrated parameters for the granier equation in a Chinese Fir (Cunninghamia lanceolata) plantation in southern China. Forests 2018, 9, 162. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cao, G.; Wang, Y.; Webb, A.A.; Yu, P.; Wang, X. Response of the daily transpiration of a larch plantation to variation in potential evaporation, leaf area index and soil moisture. Sci. Rep. 2019, 9, 4697. [Google Scholar] [CrossRef]
- Aranda, I.; Forner, A.; Cuestaa, B.; Valladares, F. Species-specific water use by forest tree species: From the tree to the stand. Agric. Water Manag. 2012, 114, 67–77. [Google Scholar] [CrossRef]
- Hu, Y.; Zhao, P.; Huang, Y.; Zhu, L.; Ni, G.; Zhao, X.; Huang, Z. Hydrologic balance, net primary productivity and water use efficiency of the introduced exotic Eucalyptus grandis × Eucalyptus urophylla plantation in south-western China. J. Plant Ecol. 2019, 12, 982–992. [Google Scholar] [CrossRef]
- Ouyang, L.; Wu, J.; Zhao, P.; Zhu, L.; Ni, G. Stand age rather than soil moisture gradient mainly regulates the compromise between plant growth and water use of Eucalyptus urophylla in hilly South China. Land Degrad. Dev. 2021, 32, 2423–2436. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Brockerhoff, E.G.; Wingfield, D.; Slippers, B. Planted forest health: The need for a global strategy. Science 2015, 349, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Ewers, B.E.; Mackay, D.S.; Gower, S.T.; Ahl, D.E.; Burrows, S.N.; Samanta, S.S. Tree species effects on stand transpiration in northern Wisconsin. Water Resour. Res. 2002, 38, 8-1–8-11. [Google Scholar] [CrossRef]
- Siddiq, Z.; Zhang, Y.; Zhu, S.; Cao, K. Canopy water status and photosynthesis of tropical trees are associated with trunk sapwood hydraulic properties. Plant Physiol. Biochem. 2019, 139, 724–730. [Google Scholar] [CrossRef]
- Siddiq, Z.; Cao, K.-F. Nocturnal transpiration in 18 broadleaf timber species under a tropical seasonal climate. For. Ecol. Manag. 2018, 418, 47–54. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Guo, J.; Wang, Y.; Ma, J.; Yu, S.; Yu, P.; Xu, L. Estimate canopy transpiration in larch plantations via the interactions among reference evapotranspiration, leaf area index, and soil moisture. For. Ecol. Manag. 2021, 481, 118749. [Google Scholar] [CrossRef]
- Xu, Z.; Man, X.; Cai, T.; Shang, Y. How Potential Evapotranspiration Regulates the Response of Canopy Transpiration to Soil Moisture and Leaf Area Index of the Boreal Larch Forest in China. Forests 2022, 13, 571. [Google Scholar] [CrossRef]
- Adil, B.; Hicham, F.; Ahmed, M.S.K.; Sanae, C.; Allal, S.; Mhamed, M.; Hassan, M. Study of Microclimate and Sapling Citrus Plant Transpiration in Tunnel Greenhouse Under Mediterranean Conditions. Acta Tech. Agric. 2022, 2, 61–66. [Google Scholar] [CrossRef]
- Abreu, M.C.; Soares, A.A.V.; de Freitas, C.H.; Martins, F.B. Transpiration and growth responses by Eucalyptus species to progressive soil drying. J. For. Res. 2022, 33, 1529–1543. [Google Scholar] [CrossRef]
- Lyu, J.; He, Q.; Chen, Q.; Cheng, R.; Li, G.; Otsuki, K.; Yamanaka, N.; Du, S. Distinct transpiration characteristics of black locust plantations acclimated to semiarid and subhumid sites in the Loess Plateau, China. Agric. Water Manag. 2022, 262, 107402. [Google Scholar] [CrossRef]
- Han, C.; Chen, N.; Zhang, C.; Liu, Y.; Khan, S.; Lu, K.; Li, Y.; Dong, X.; Zhao, C. Sap flow and responses to meteorological about the Larix principis-rupprechtii plantation in Gansu Xinlong mountain, northwestern China. For. Ecol. Manag. 2019, 451, 117519. [Google Scholar] [CrossRef]
- Tu, J.; Wei, X.; Huang, B.; Fan, H.; Jian, M.; Li, W. Improvement of sap flow estimation by including phenological index and time-lag effect in back-propagation neural network models. Agric. For. Meteorol. 2019, 276, 107608. [Google Scholar] [CrossRef]
- Oogathoo, S.; Houle, D.; Duchesne, L.; Kneeshaw, D. Vapour pressure deficit and solar radiation are the major drivers of transpiration of balsam fir and black spruce tree species in humid boreal regions, even during a short-term drought. Agric. For. Meteorol. 2020, 291, 108063. [Google Scholar] [CrossRef]
- Urban, J.; Rubtsov, A.V.; Urban, A.V.; Shashkin, A.V.; Vera, E.B. Canopy transpiration of a Larix sibirica and Pinus sylvestris forest in Central Siberia. Agric. For. Meteorol. 2019, 271, 64–72. [Google Scholar] [CrossRef]
- Heidi, H.; Michael, H.A.; Susan, C.; Michael, C.; Sjirk, G.; Coert, J.G.; Graham, H.; Brett, P.H.; Wayne, J.; Jan-Hendrik, K.; et al. Eucalyptus camaldulensis in South Africa—Past, present, future. Trans. R. Soc. South Afr. 2020, 75, 1–22. [Google Scholar] [CrossRef]
- Yang, G.; Lu, W.; Lin, Y.; Luo, J.; Wang, C.; Meder, R.; Warburton, P.M.; Arnold, R.J. Monitoring water potential and relative water content in eucalyptus camaldulensis using near infrared spectroscopy. J. Trop. For. Sci. 2017, 29, 121–128. [Google Scholar]
- Saravanan, T.; Roger, J.A.; Luo, J.; Bala, R.T. Genomic studies reveal substantial dominant effects and improved genomic predictions in an open-pollinated breeding population of Eucalyptus pellita. G3 Genes Genomes Genet. 2020, 10, 3751–3763. [Google Scholar] [CrossRef]
- Wirabuana, P.Y.A.P.; Sadono, R.; Juniarso, S. Fertilization Effects on Early Growth, Aboveground Biomass, Carbon Storage, and Leaf Characteristics of Eucalyptus pellita F. Muell. in South Sumatera. J. Man. Hut. Trop. 2019, 25, 154. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Du, A.; Wang, Z.; Zhu, W.; Li, C.; Wu, L. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2020, 456, 117683. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World reference base for soil resource 2006. In World Soil Resources Reports No. 103, 2nd ed.; FAO: Rome, Italy, 2006. [Google Scholar]
- Campbell, G.S.; Norman, J. An Introduction to Environmental Biophysics, 2nd ed.; Springer: New York, NY, USA, 1998. [Google Scholar]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef]
- Lu, P.; Urban, L.; Zhao, P. Granier’s Thermal Dissipation Probe (TDP) Method for Measuring Sap Flow in Trees: Theory and Practice. Acta Bot. Sin. 2004, 46, 631–646. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Benyon, R.G.; O’Sullivan, S.K.; Gribben, P.R. Relationships between stem diameter, sapwood area, leaf area and transpiration in a young mountain ash forest. Tree Physiol. 1995, 15, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Nadezhdina, N. Sap flow index as an indicator of plant water status. Tree Physiol. 1999, 19, 885–891. [Google Scholar] [CrossRef]
- Peng, X.; Jun, F.; Wang, Q.; Warrington, D. Discrepancy of sap flow in Salix matsudana grown under different soil textures in the water-wind erosion crisscross region on the Loess Plateau. Plant Soil 2015, 390, 383–399. [Google Scholar] [CrossRef]
- Ma, C.; Luo, Y.; Shao, M.; Li, X.; Sun, L.; Jia, X. Environmental controls on sap flow in black locust forest in Loess Plateau, China. Sci. Rep. 2017, 7, 13160. [Google Scholar] [CrossRef]
- Tie, Q.; Hu, H.; Tian, F.; Guan, H.; Lin, H. Environmental and physiological controls on sap flow in a subhumid mountainous catchment in North China. Agric. For. Meteorol. 2017, 240, 46–57. [Google Scholar] [CrossRef]
- Forrester, D.I.; Collopy, J.J.; Morris, J.D. Transpiration along an age series of Eucalyptus globulus plantations in southeastern Australia. For. Ecol. Manag. 2010, 259, 1754–1760. [Google Scholar] [CrossRef]
- Cheng, J.; Ou, Y.; Huang, D.; Liu, S.; Li, Y. Sap Flow Characteristics of Four Dominant Tree Species in a Mixed Conifer-broadleaf Forest in Dinghushan. Acta Ecol. Sin. 2015, 35, 4097–4104. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.J.; Benyon, R.G.; Lane, P.N.J. Responses of evapotranspiration at different topographic positions and catchment water balance following a pronounced drought in a mixed species eucalypt forest, Australia. J. Hydrol. 2012, 440, 62–74. [Google Scholar] [CrossRef]
- Macfarlane, C.; Bond, C.; White, D.A.; Grigg, A.H.; Ogden, G.N.; Silberstein, R. Transpiration and hydraulic traits of old and regrowth eucalypt forest in southwestern Australia. For. Ecol. Manag. 2010, 260, 96–105. [Google Scholar] [CrossRef]
- Almeida, A.C.; Soares, J.V.; Landsberg, J.J.; Rezende, G.D. Growth and water balance of Eucalyptus grandis hybrid plantations in Brazil during a rotation for pulp production. For. Ecol. Manag. 2007, 251, 10–21. [Google Scholar] [CrossRef]
- Hernandez-Santana, V.; Hernandez-Hernandez, A.; Vadeboncoeur, M.A.; Asbjornsen, H. Scaling from single-point sap velocity measurements to stand transpiration in a multispecies deciduous forest: Uncertainty sources, stand structure effect, and future scenarios. Can. J. For. Res. 2015, 45, 1489–1497. [Google Scholar] [CrossRef]
- Link, P.; Simonin, K.A.; Maness, H.; Oshun, J.; Dawson, T.; Fung, I. Species differences in the seasonality of evergreen tree transpiration in a Mediterranean climate: Analysis of multiyear, half- hourly sap flow observations. Water Resour. Res. 2014, 50, 1869–1894. [Google Scholar] [CrossRef]
- Yan, M.; Zhang, J.; He, Q.; Shi, W.; Otsuki, K.; Yamanaka, N.; Du, S. Sapflow-Based Stand Transpiration in a Semiarid Natural Oak Forest on China’s Loess Plateau. Forests 2016, 7, 227. [Google Scholar] [CrossRef]
- Horna, V.; Schuldt, B.; Brix, S.; Leuschner, C. Environment and tree size controlling stem sap flux in a perhumid tropical forest of Central Sulawesi, Indonesia. Ann. For. Sci. 2011, 68, 1027–1038. [Google Scholar] [CrossRef]
- Di, N.; Xi, B.; Clothier, B.; Wang, Y.; Li, G.; Jia, L. Diurnal and nocturnal transpiration behaviors and their responses to groundwater-table fluctuations and meteorological factors of Populus tomentosa in the North China Plain. For. Ecol. Manag. 2019, 448, 445–456. [Google Scholar] [CrossRef]
- Hatton, T.; Reece, P.; Taylor, P.; McEwan, K. Does leaf water efficiency vary among eucalypts in water-limited environments? Tree Physiol. 1998, 18, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, Y.; Zhu, W.; Du, A. The transpiration water consumption of two common fast-growing forests in Leizhou Peninsula. Acta Ecol. Sin. 2019, 39, 2147–2155. [Google Scholar] [CrossRef]
- Ma, L.; Lu, P.; Zhao, P.; Rao, X.; Cai, X.; Zeng, X. Diurnal, daily, seasonal and annual patterns of sap-flux-scaled transpiration from an Acacia mangium plantation in South China. Ann. For. Sci. 2008, 65, 402. [Google Scholar] [CrossRef]
- Tian, Q.; He, Z.; Xiao, S.; Du, J. Growing Season Stem Water Status Assessment of Qinghai Spruce through the Sap Flow and Stem Radial Variations in the Qilian Mountains of China. Forests 2018, 9, 2. [Google Scholar] [CrossRef]
- Ghimire, C.P.; Lubczynski, M.W.; Bruijnzeel, L.A.; Chavarro-Rincón, D. Transpiration and canopy conductance of two contrasting forest types in the Lesser Himalaya of Central Nepal. Agric. For. Meteorol. 2014, 197, 76–90. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Wilson, K.B.; Hanson, P.J. Environmental control of whole-plant transpiration, canopy conductance and estimates of the decoupling coefficient for large red maple trees. Agric. For. Meteorol. 2000, 104, 157–168. [Google Scholar] [CrossRef]
- White, D.A.; Beadle, C.L.; Worledge, D. Control of transpiration in an irrigated Eucalyptus globulus Labill. plantation. Plant Cell Environ. 2000, 23, 123–134. [Google Scholar] [CrossRef]
- Chang, X.; Zhao, W.; Liu, H.; Wei, X.; Liu, B.; He, Z. Qinghai spruce (Picea crassifolia) forest transpiration and canopy conductance in the upper Heihe River Basin of arid northwestern China. Agric. For. Meteorol. 2014, 198, 209–220. [Google Scholar] [CrossRef]
- Chen, D.; Wang, Y.; Liu, S.; Wei, X.; Wang, X. Response of relative sap flow to meteorological factors under different soil moisture conditions in rainfed jujube (Ziziphus jujuba Mill.) plantations in semiarid Northwest China. Agri. Water Manag. 2014, 136, 23–33. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Sun, G.; Ward, E.J.; Fu, B. Biophysical controls on canopy transpiration in a black locust (Robinia pseudoacacia) plantation on the semi-arid Loess Plateau, China. Ecohydrology 2016, 9, 1068–1081. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Oberbauer, S.F.; Clark, D.B. Whole tree xylem sap flow responses to multiple environmental variables in a wet tropical forest. Plant Cell Environ. 2004, 27, 551–567. [Google Scholar] [CrossRef]
- Asbjornsen, H.; Tomer, M.D.; Gomez-Cardenas, M.; Brudvig, L.A.; Greenan, C.M.; Schilling, K. Tree and stand transpiration in a Midwestern bur oak savanna after elm encroachment and restoration thinning. For. Ecol. Manag. 2007, 247, 209–219. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Goldstein, G.; Jackson, P.; Holbrook, N.M.; Guti6rrez, M.V.; Cavelier, J. Environmental and physiological regulation of transpiration in tropical forest gap species: The influence of boundary layer and hydraulic properties. Oecologia 1995, 101, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.B.; Baldocchi, D.D.; Misson, L.; Dawson, T.E.; Goldstein, A.H. What the towers don’t see at night: Nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California. Tree Physiol. 2007, 27, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Si, J.; Qi, F.; Yu, T.; Li, P. Effect of wind speed on transpiration rate of Populus euphratica in extreme arid deserts. J. Glaciol. Geocryol. 2015, 37, 1104–1111. [Google Scholar] [CrossRef]
- Campbell-Clause, J.M. Stomatal response of grapevines to wind. Aust. J. Exp. Agric. 1998, 38, 77–82. [Google Scholar] [CrossRef]
- Gutirrez, M.V.; Meinzer, F.C.; Grantz, D.A. Regulation of transpiration in coffee hedgerows: Covariation of environmental variables and apparent responses of stomata to wind and humidity. Plant Cell Environ. 1994, 17, 1305–1313. [Google Scholar] [CrossRef]
- Buckley, T.N.; Turnbull, T.L.; Pfautsch, S.; Adams, M.A. Nocturnal water loss in mature subalpine Eucalyptus delegatensis tall open forests and adjacent E. pauciflora woodlands. Ecol. Evol. 2011, 1, 435–450. [Google Scholar] [CrossRef]
- Galmés, J.; Flexas, J.; Savé, R.; Medrano, H. Water relations and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: Responses to water stress and recovery. Plant Soil 2007, 290, 139–155. [Google Scholar] [CrossRef]
- Oren, R.; Pataki, D.E. Transpiration in response to variation in microclimate and soil moisture in southeastern deciduous forests. Oecologia 2001, 127, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Bindi, M.; Bellesi, S.; Orlandini, S.; Fibbi, L.; Moriondo, M.; Sinclair, T. Influence of Water Deficit Stress on Leaf Area Development and Transpiration of Sangiovese Grapevines Grown in Pots. Am. J. Enol. Vitic. 2005, 56, 68–72. [Google Scholar] [CrossRef]
- Li, Z.; Yu, P.; Wang, Y.; Webb, A.A.; He, C.; Wang, Y.; Yang, L. A model coupling the effects of soil moisture and potential evaporation on the tree transpiration of a semi-arid larch plantation. Ecohydrology 2016, 10, e1764. [Google Scholar] [CrossRef]
- Ungar, E.D.; Rotenberg, E.; Raz-Yaseef, N.; Cohen, S.; Yakir, D.; Schiller, G. Transpiration and annual water balance of Aleppo pine in a semiarid region: Implications for forest management. For. Ecol. Manag. 2013, 298, 39–51. [Google Scholar] [CrossRef]
- Ayyoub, A.; Er-Raki, S.; Khabba, S.; Merlin, O.; Ezzahar, J.; Rodriguez, J.C.; Bahlaoui, A.; Chehbouni, A. A simple and alternative approach based on reference evapotranspiration and leaf area index for estimating tree transpiration in semi-arid regions. Agric. Water Manag. 2017, 188, 61–68. [Google Scholar] [CrossRef]
- Naithani, K.J.; Baldwin, D.C.; Gaines, K.P.; Lin, H.; Eissenstat, D.M. Spatial Distribution of Tree Species Governs the Spatio-Temporal Interaction of Leaf Area Index and Soil Moisture across a Forested Landscape. PLoS ONE 2013, 8, e58704. [Google Scholar] [CrossRef]
- Forrester, D.I.; Collopy, J.J.; Beadle, C.L.; Warren, C.R.; Baker, T.G. Effect of thinning, pruning and nitrogen fertiliser application on transpiration, photosynthesis and water-use efficiency in a young Eucalyptus nitens plantation. For. Ecol. Manag. 2012, 266, 286–300. [Google Scholar] [CrossRef]
- Di, N.; Wang, Y.; Clothier, B.; Liu, Y.; Jia, L.; Xi, B.; Shi, H. Modeling soil evaporation and the response of the crop coefficient to leaf area index in mature Populus tomentosa plantations growing under different soil water availabilities. Agric. For. Meteorol. 2019, 264, 125–137. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Hoffmann, W.A.; Meinzer, F.C.; Franco, A.C.; Giambelluca, T.; Miralles-Wilhelm, F. Controls on stand transpiration and soil water utilization along a tree density gradient in a Neotropical savanna. Agric. For. Meteorol. 2008, 148, 839–849. [Google Scholar] [CrossRef]
- Li, L.; Chen, S.; Yang, C.; Meng, F.; Sigrimis, N. Prediction of plant transpiration from environmental parameters and relative leaf area index using the random forest regression algorithm. J. Clean. Prod. 2020, 261, 121136. [Google Scholar] [CrossRef]
- Liu, N.; Buckley, T.; He, X.; Zhang, X.; Zhang, C.; Luo, Z.; Wang, H.; Sterling, N.; Guan, H. Improvement of a simplified process-based model for estimating transpiration under water-limited conditions. Hydrol. Process. 2019, 33, 1670–1685. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Fang, W.; Li, Z.; Wang, J.; Jin, Z. Determining the independent impact of soil water on forest transpiration: A case study of a black locust plantation in the Loess Plateau, China. J. Hydrol. 2019, 572, 671–681. [Google Scholar] [CrossRef]
- Granier, A.; Loustaub, D.; Bréda, N. A generic model of forest canopy conductance dependent on climate, soil water availability and leaf area index. Ann. For. Sci. 2000, 57, 755–765. [Google Scholar] [CrossRef]
- Sawano, S.; Hotta, N.; Tanaka, N.; Tsuboyama, Y.; Suzuki, M. Development of a simple forest evapotranspiration model using a process-oriented model as a reference to parameterize data from a wide range of environmental conditions. Ecol. Modell. 2015, 309, 93–109. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Ep | Ec | Eug |
|---|---|---|---|
| Age (year) | 12 | 12 | 12 |
| Mean DBH (cm) | 22.93 ± 1.78 a | 19.01 ± 1.65 b | 23.52 ± 1.19 a |
| Mean height (m) | 21.67 ± 1.99 b | 19.77 ± 0.93 b | 27.1 ± 0.32 a |
| Stand LAI | 2.43 ± 0.13 a | 1.87 ± 0.05 c | 2.08 ± 0.05 b |
| Stem density (plants/ha) | 950 | 1050 | 1175 |
| Mean sapwood area (cm2/plant) | 134.31 ± 18.15 b | 70.78 ± 7.85 c | 189.88 ± 12.02 a |
| Stand sapwood area (m2/ha) | 12.34 | 7.18 | 21.49 |
| Species | Ep | Ec | Eug | Precipitation (mm) | |||
|---|---|---|---|---|---|---|---|
| Time Scale | Monthly Water Use (mm) | Daily Mean Water Use (mm/d) | Monthly Water Use (mm) | Daily Mean Water Use (mm/d) | Monthly Water Use (mm) | Daily Mean Water Use (mm/d) | |
| Jan | 47.85 | 1.54 ± 0.07 cd/A | 16.09 | 0.52 ± 0.03 d/C | 35.73 | 1.15 ± 0.07 bc/B | 27.6 |
| Feb | 48.97 | 1.69 ± 0.08 cd/A | 18.84 | 0.65 ± 0.05 d/C | 30.14 | 1.04 ± 0.06 bc/B | 48.9 |
| Mar | 49.37 | 1.59 ± 0.11 cd/A | 18.90 | 0.61 ± 0.05 d/C | 32.81 | 1.06 ± 0.08 bc/B | 46.6 |
| Apr | 45.63 | 1.52 ± 0.16 cd/A | 21.13 | 0.70 ± 0.06 cd/B | 31.41 | 1.05 ± 0.10 bc/B | 112.3 |
| May | 85.00 | 2.74 ± 0.09 a/A | 33.59 | 1.08 ± 0.04 ab/C | 66.24 | 2.14 ± 0.07 a/B | 58.8 |
| Jun | 84.76 | 2.83 ± 0.11 a/A | 32.42 | 1.08 ± 0.04 ab/C | 58.18 | 1.94 ± 0.09 a/B | 121.6 |
| Jul | 91.92 | 2.97 ± 0.06 a/A | 38.55 | 1.24 ± 0.03 a/C | 56.29 | 1.82 ± 0.04 a/B | 121.3 |
| Aug | 68.08 | 2.20 ± 0.18 b/A | 28.66 | 0.92 ± 0.07 bc/B | 40.57 | 1.31 ± 0.10 b/B | 282.8 |
| Sep | 66.67 | 2.22 ± 0.11 b/A | 27.85 | 0.93 ± 0.05 b/C | 38.43 | 1.28 ± 0.07 b/B | 373.3 |
| Oct | 60.09 | 1.94 ± 0.12 bc/A | 30.93 | 1.00 ± 0.06 b/B | 38.49 | 1.24 ± 0.08 b/B | 260.7 |
| Nov | 55.26 | 1.84 ± 0.06 bc/A | 26.78 | 0.89 ± 0.03 bc/C | 38.23 | 1.27 ± 0.05 b/B | 6.9 |
| Dec | 39.81 | 1.28 ± 0.08 d/A | 17.78 | 0.57 ± 0.04 d/C | 27.06 | 0.87 ± 0.05 c/B | 1.5 |
| Monthly mean | 61.95 A | —— | 25.96 C | —— | 41.13 B | —— | 121.86 |
| Annual total | 743.41 | 2.03 ± 0.04 A | 311.52 | 0.85 ± 0.02 C | 493.58 | 1.35 ± 0.03 B | 1462.3 |
| Dry season total | 286.89 | 1.58 ± 0.04 A | 119.52 | 0.66 ± 0.02 C | 195.38 | 1.07 ± 0.03 B | 243.8 |
| Wet season total | 456.52 | 2.48 ± 0.05 A | 192.00 | 1.04 ± 0.02 C | 298.20 | 1.62 ± 0.04 B | 1218.5 |
| Eucalyptus Species | Predictive Model of T | R2 | n | Sig. |
|---|---|---|---|---|
| Ep | T = −1.9 × 10−5Rs2 + 0.0153Rs + 2.05 × 10−3Ta2 − 0.0586Ta − 8.57 × 10−4RH2 + 0.119 RH + 1.03× 107exp(−SWC/0.9655) + 0.161exp(−exp(−4.35(LAI − 2.044))) − 3.515 | 0.911 | 305 | 0.000 |
| Ec | T = −9.19 × 10−6Rs2 + 5.91 × 10−3Rs − 0.224VPD2 + 0.78VPD − 6.22 × 10−4Ta2 + 0.016 Ta + 1.97× 109exp(−SWC/0.6893) + 0.514 exp(−exp(−7.45(LAI − 1.575))) − 0.568 | 0.872 | 305 | 0.000 |
| Eug | T = −1.01 × 10−5Rs2 + 7.76 × 10−3Rs − 0.202VPD2 + 0.97VPD + 5.85 × 108exp(−SWC/0.7999) + 0.822exp(−exp(−0.858(LAI − 1.561))) − 0.541 | 0.863 | 305 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Liu, S.; Xu, Y.; Zhu, W.; Du, A. Differences in Transpiration Characteristics among Eucalyptus Plantations of Three Species on the Leizhou Peninsula, Southern China. Forests 2022, 13, 1544. https://doi.org/10.3390/f13101544
Wang Z, Liu S, Xu Y, Zhu W, Du A. Differences in Transpiration Characteristics among Eucalyptus Plantations of Three Species on the Leizhou Peninsula, Southern China. Forests. 2022; 13(10):1544. https://doi.org/10.3390/f13101544
Chicago/Turabian StyleWang, Zhichao, Siru Liu, Yuxing Xu, Wankuan Zhu, and Apeng Du. 2022. "Differences in Transpiration Characteristics among Eucalyptus Plantations of Three Species on the Leizhou Peninsula, Southern China" Forests 13, no. 10: 1544. https://doi.org/10.3390/f13101544