
Unraveling the Importance of Forest Structure and Composition Driving Soil Microbial and Enzymatic Responses in the Subtropical Forest Soils

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Forest Composition
2.2. Soil Sampling and Treatment
2.3. Chemical Analysis of Soil Samples
2.3.1. Soil pH and Enzymes Activity Determination
2.3.2. Soil Microbial Activity Analysis
2.4. Statistical Analysis
3. Results
3.1. Soil pH, DOC, and Enzymatic Responses
3.2. Soil Microbial Properties and Microbial Quotient
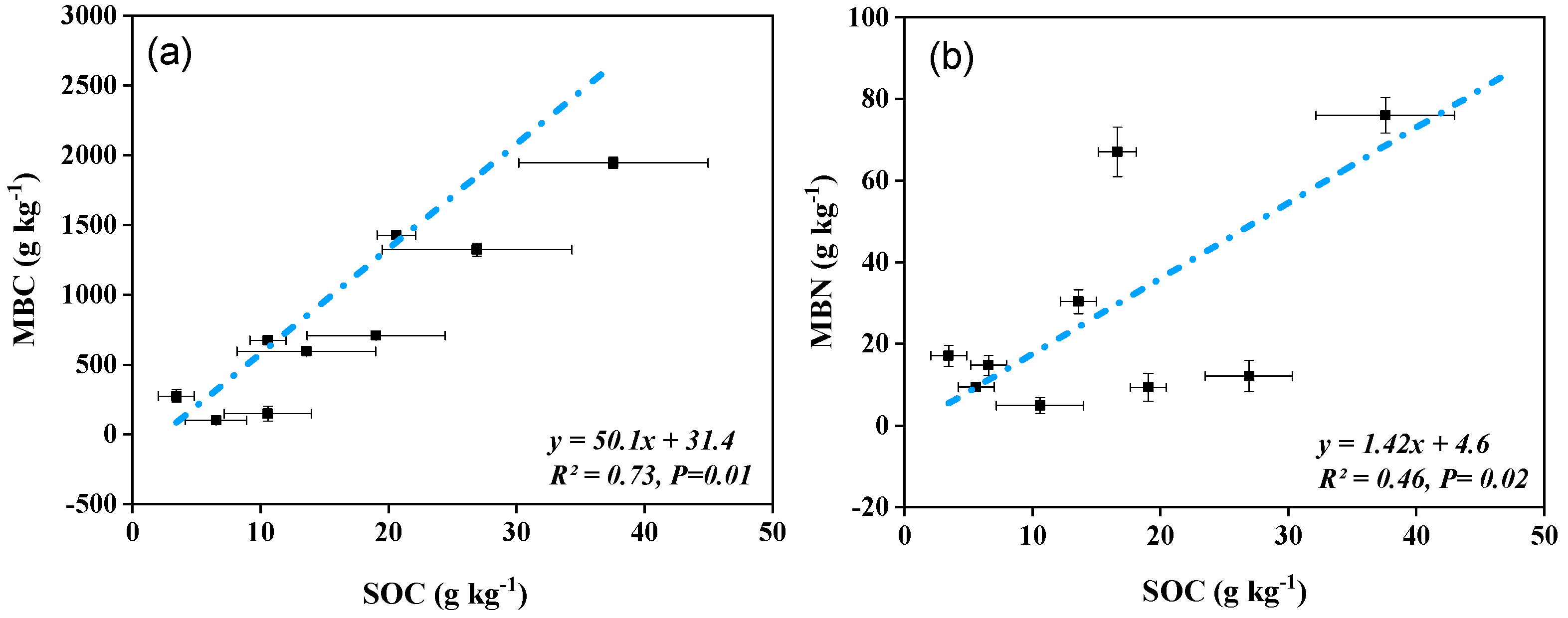
3.3. Correlations between Soil Enzymatic Responses, Chemical, and Microbial Properties
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pertile, M.; Antunes, J.E.L.; Araujo, F.F.; Mendes, L.W.; Van den Brink, P.J.; Araujo, A.S.F. Responses of soil microbial biomass and enzyme activity to herbicides imazethapyr and flumioxazin. Sci. Rep. 2020, 10, 7694. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.H.; Ma, X.; Rashid, M.H.U.; Wu, W.; Xu, J.; Tarin, M.W.K.; He, Z.; Wu, P. Impact of stand density on soil quality in Chinese Fir (Cunninghamia Lanceolata) monoculture. Appl. Ecol. Environ. Res. 2019, 17, 3553–3566. [Google Scholar] [CrossRef]
- Yan, Y.; Li, B.; Huang, Z.; Zhang, H.; Wu, X.; Farooq, T.H.; Wu, P.; Li, M.; Ma, X. Characteristics and driving factors of rhizosphere bacterial communities of Chinese Fir Provenances. Forests 2021, 12, 1362. [Google Scholar] [CrossRef]
- Durán, J.; Morse, J.L.; Groffman, P.M.; Campbell, J.L.; Christenson, L.M.; Driscoll, C.T.; Fahey, T.J.; Fisk, M.C.; Mitchell, M.J.; Templer, P.H. Winter climate change affects growing-season soil microbial biomass and activity in northern hardwood forests. Glob. Chang. Biol. 2014, 20, 3568–3577. [Google Scholar] [CrossRef] [PubMed]
- Kaschuk, G.; Alberton, O.; Hungria, M. Three decades of soil microbial biomass studies in Brazilian ecosystems: Lessons learned about soil quality and indications for improving sustainability. Soil Biol. Biochem. 2010, 42, 1–13. [Google Scholar] [CrossRef]
- McDaniel, M.D.; Grandy, A.S. Soil microbial biomass and function are altered by 12 years of crop rotation. Soil 2016, 2, 583–599. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Rinkes, Z.L.; Sinsabaugh, R.L.; Weintraub, M.N. Dynamic relationships between microbial biomass, respiration, inorganic nutrients and enzyme activities: Informing enzyme-based decomposition models. Front. Microbiol. 2013, 4, 223. [Google Scholar] [CrossRef]
- Mgelwa, A.S.; Hu, Y.-L.; Xu, W.-B.; Ge, Z.-Q.; Yu, T.-W. Soil carbon and nitrogen availability are key determinants of soil microbial biomass and respiration in forests along urbanized rivers of southern China. Urban For. Urban Green. 2019, 43, 126351. [Google Scholar] [CrossRef]
- Antisari, L.V.; Carbone, S.; Gatti, A.; Vianello, G.; Nannipieri, P. Toxicity of metal oxide (CeO2, Fe3O4, SnO2) engineered nanoparticles on soil microbial biomass and their distribution in soil. Soil Biol. Biochem. 2013, 60, 87–94. [Google Scholar] [CrossRef]
- Pose-Juan, E.; Igual, J.M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Influence of herbicide triasulfuron on soil microbial community in an unamended soil and a soil amended with organic residues. Front. Microbiol. 2017, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wang, Y.; Hui, D.; Jing, X.; Feng, W. Soil properties rather than climate and ecosystem type control the vertical variations of soil organic carbon, microbial carbon, and microbial quotient. Soil Biol. Biochem. 2020, 148, 107905. [Google Scholar] [CrossRef]
- Sahu, N.; Vasu, D.; Sahu, A.; Lal, N.; Singh, S.K. Strength of microbes in nutrient cycling: A key to soil health. In Agriculturally Important Microbes for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2017; pp. 69–86. [Google Scholar]
- Sherene, T. Role of soil enzymes in nutrient transformation: A review. Bio Bull. 2017, 3, 109–131. [Google Scholar]
- Farooq, T.H.; Kumar, U.; Mo, J.; Shakoor, A.; Wang, J.; Rashid, M.H.U.; Tufail, M.A.; Chen, X.; Yan, W. Intercropping of peanut–tea enhances soil enzymatic activity and soil nutrient status at different soil profiles in subtropical southern China. Plants 2021, 10, 881. [Google Scholar] [CrossRef]
- Decina, S.M.; Hutyra, L.R.; Gately, C.K.; Getson, J.M.; Reinmann, A.B.; Gianotti, A.G.S.; Templer, P.H. Soil respiration contributes substantially to urban carbon fluxes in the greater Boston area. Environ. Pollut. 2016, 212, 433–439. [Google Scholar] [CrossRef]
- Sarzhanov, D.A.; Vasenev, V.I.; Vasenev, I.I.; Sotnikova, Y.L.; Ryzhkov, O.V.; Morin, T. Carbon stocks and CO2 emissions of urban and natural soils in Central Chernozemic region of Russia. Catena 2017, 158, 131–140. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- DeForest, J.L.; Zak, D.R.; Pregitzer, K.S.; Burton, A.J. Atmospheric nitrate deposition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest. Soil Biol. Biochem. 2004, 36, 965–971. [Google Scholar] [CrossRef]
- Koerner, B.A.; Klopatek, J.M. Carbon fluxes and nitrogen availability along an urban–rural gradient in a desert landscape. Urban Ecosyst. 2010, 13, 1–21. [Google Scholar] [CrossRef]
- Lorenz, K.; Kandeler, E. Microbial biomass activities in urban soils in two consecutive years. J. Plant Nutr. Soil Sci. 2006, 169, 799–808. [Google Scholar] [CrossRef]
- Sushko, S.; Ananyeva, N.; Ivashchenko, K.; Vasenev, V.; Kudeyarov, V. Soil CO2 emission, microbial biomass, and microbial respiration of woody and grassy areas in Moscow (Russia). J. Soils Sediments 2019, 19, 3217–3225. [Google Scholar] [CrossRef]
- Farooq, T.H.; Yan, W.; Rashid, M.H.U.; Tigabu, M.; Gilani, M.M.; Zou, X.H.; Wu, P.F. Chinese fir (Cunninghamia Lanceolata) a green gold of China with continues decline in its productivity over the successive rotations: A review. Appl. Ecol. Environ. Res. 2019, 17, 11055–11067. [Google Scholar] [CrossRef]
- Farooq, T.H.; Shakoor, A.; Wu, X.; Li, Y.; Rashid, M.H.U.; Zhang, X.; Gilani, M.M.; Kumar, U.; Chen, X.; Yan, W. Perspectives of plantation forests in the sustainable forest development of China. iFor.-Biogeosci. For. 2021, 14, 166–174. [Google Scholar] [CrossRef]
- Farooq, T.H.; Wu, W.; Tigabu, M.; Ma, X.; He, Z.; Rashid, M.H.U.; Gilani, M.M.; Wu, P. Growth, biomass production and root development of Chinese fir in relation to initial planting density. Forests 2019, 10, 236. [Google Scholar] [CrossRef]
- He, Y.; Wang, Q.; Wang, S.; Yu, X. Characteristics of soil microbial biomass carbon and nitrogen and their relationships with soil nutrients in Cunninghamia lanceolata plantations. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2006, 17, 2292–2296. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J.; Yan, Q.L.; Sun, O.J. Changes in soil P chemistry as affected by conversion of natural secondary forests to larch plantations. For. Ecol. Manag. 2010, 260, 422–428. [Google Scholar] [CrossRef]
- Burton, J.; Chen, C.; Xu, Z.; Ghadiri, H. Gross nitrogen transformations in adjacent native and plantation forests of subtropical Australia. Soil Biol. Biochem. 2007, 39, 426–433. [Google Scholar] [CrossRef]
- Farooq, T.H.; Xincheng, X.; Shakoor, A.; Rashid, M.H.U.; Bashir, M.F.; Nawaz, M.F.; Kumar, U.; Shahzad, S.M.; Yan, W. Spatial distribution of carbon dynamics and nutrient enrichment capacity in different layers and tree tissues of Castanopsis eyeri natural forest ecosystem. Environ. Sci. Pollut. Res. 2022, 29, 10250–10262. [Google Scholar] [CrossRef]
- Farooq, T.H.; Chen, X.; Shakoor, A.; Li, Y.; Wang, J.; Rashid, M.H.U.; Kumar, U.; Yan, W. Unraveling the influence of land-use change on δ13C, δ15N, and soil nutritional status in coniferous, broadleaved, and mixed forests in southern china: A field investigation. Plants 2021, 10, 1499. [Google Scholar] [CrossRef]
- Paudel, B.R.; Udawatta, R.P.; Anderson, S.H. Agroforestry and grass buffer effects on soil quality parameters for grazed pasture and row-crop systems. Appl. Soil Ecol. 2011, 48, 125–132. [Google Scholar] [CrossRef]
- Hu, Z.; He, Z.; Huang, Z.; Fan, S.; Yu, Z.; Wang, M.; Zhou, X.; Fang, C. Effects of harvest residue management on soil carbon and nitrogen processes in a Chinese fir plantation. For. Ecol. Manag. 2014, 326, 163–170. [Google Scholar] [CrossRef]
- Yang, L.; Li, T.; Li, F.; Lemcoff, J.H.; Cohen, S. Fertilization regulates soil enzymatic activity and fertility dynamics in a cucumber field. Sci. Hortic. 2008, 116, 21–26. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Swift, M.J.; Heal, O.W.; Anderson, J.M.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979; Volume 5, ISBN 0520040015. [Google Scholar]
- Haynes, R.J. Labile organic matter fractions as centralcomponents of the quality of agricultural soils: Anoverview. Adv. Agron. 2005, 5, 221–268. [Google Scholar]
- Xiong, Y.; D’Atri, J.J.; Fu, S.; Xia, H.; Seastedt, T.R. Rapid soil organic matter loss from forest dieback in a subalpine coniferous ecosystem. Soil Biol. Biochem. 2011, 43, 2450–2456. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Allard, V.; Robin, C.; Newton, P.C.D.; Lieffering, M.; Soussana, J.F. Short and long-term effects of elevated CO2 on Lolium perenne rhizodeposition and its consequences on soil organic matter turnover and plant N yield. Soil Biol. Biochem. 2006, 38, 1178–1187. [Google Scholar] [CrossRef]
- Friedel, J.K.; Langer, T.; Siebe, C.; Stahr, K. Effects of long-term waste water irrigation on soil organic matter, soil microbial biomass and its activities in central Mexico. Biol. Fertil. Soils 2000, 31, 414–421. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought decreases soil enzyme activity in a Mediterranean Quercus ilex L. forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Hou-Long, J.; Zheng, L.; Shu-Duan, L.; An-Ding, X.; Chao, Y.; Yan, Z.; Guo-Shun, L.; Xie-Feng, Y. Effects of consecutive turnover of green manure and N fertilizer on soil microbial biomass and enzyme activity in tobacco-planted field. Indian J. Agric. Res. 2013, 47, 185–191. [Google Scholar]
- Zhang, Y.; Cui, D.; Yang, H.; Kasim, N. Differences of soil enzyme activities and its influencing factors under different flooding conditions in Ili Valley, Xinjiang. PeerJ 2020, 8, e8531. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, H.A.; Brussaard, L. Decomposition and nitrogen mineralization in natural and agroecosystems: The contribution of soil animals. Biogeochemistry 1990, 11, 175–211. [Google Scholar] [CrossRef]
- Wang, H.; Wu, J.; Li, G.; Yan, L. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau. Ecol. Evol. 2020, 10, 12211–12223. [Google Scholar] [CrossRef]
- Cheng, F.; Peng, X.; Zhao, P.; Yuan, J.; Zhong, C.; Cheng, Y.; Cui, C.; Zhang, S. Soil microbial biomass, basal respiration and enzyme activity of main forest types in the Qinling Mountains. PLoS ONE 2013, 8, e67353. [Google Scholar] [CrossRef]
- Fanin, N.; Moorhead, D.; Bertrand, I. Eco-enzymatic stoichiometry and enzymatic vectors reveal differential C, N, P dynamics in decaying litter along a land-use gradient. Biogeochemistry 2016, 129, 21–36. [Google Scholar] [CrossRef]
- Bradford, M.A.; Davies, C.A.; Frey, S.D.; Maddox, T.R.; Melillo, J.M.; Mohan, J.E.; Reynolds, J.F.; Treseder, K.K.; Wallenstein, M.D. Thermal adaptation of soil microbial respiration to elevated temperature. Ecol. Lett. 2008, 11, 1316–1327. [Google Scholar] [CrossRef]
- Yao, H.; He, Z.L.; Wilson, M.; Campbell, C.D. Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb. Ecol. 2000, 40, 223–237. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Xue, S.; Song, Z. Geoderma Rhizosphere soil microbial activity under different vegetation types on the Loess. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Zhu, L.; Xiao, Q.; Cheng, H.; Shi, B.; Shen, Y.; Li, S. Seasonal dynamics of soil microbial activity after biochar addition in a dryland maize field in North-Western China. Ecol. Eng. 2017, 104, 141–149. [Google Scholar] [CrossRef]
- Shi, W. Agricultural and ecological significance of soil enzymes: Soil carbon sequestration and nutrient cycling. In Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 43–60. [Google Scholar]
- Amin, B.A.Z.; Chabbert, B.; Moorhead, D.; Bertrand, I. Impact of fine litter chemistry on lignocellulolytic enzyme efficiency during decomposition of maize leaf and root in soil. Biogeochemistry 2014, 117, 169–183. [Google Scholar] [CrossRef]
- Yan, W.; Farooq, T.H.; Chen, Y.; Wang, W.; Shabbir, R.; Kumar, U.; Riaz, M.U.; Alotaibi, S.S.; Peng, Y.; Chen, X. Soil Nitrogen Transformation Process Influenced by Litterfall Manipulation in Two Subtropical Forest Types. Front. Plant Sci. 2022, 13, 923410. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.H.; Li, Z.; Yan, W.; Shakoor, A.; Kumar, U.; Shabbir, R.; Peng, Y.; Alotaibi, S.; Wróbel, J.; Kalaji, H.M. Variations in litterfall dynamics, C: N: P stoichiometry and associated nutrient return in pure and mixed stands of Camphor tree and Masson pine forests in Subtropical China. Front. Environ. Sci. 2022, 10, 903039. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil | Soil Depth | Forest Types | ||
|---|---|---|---|---|
| Attributes | (cm) | NBF a | Mixed | CPF |
| Soil pH | 0–10 | 4.21 (0.62) cB | 4.67 (0.15) bA | 5.17 (0.18) aA |
| 10–20 | 4.50 (0.24) bA | 4.54 (0.21) bB | 4.72 (0.32) aB | |
| 20–40 | 4.51 (0.29) bA | 4.58 (0.16) aB | 4.51 (0.35) bC | |
| SOC (g.kg−1) | 0–10 | 19.04 (0.36) bB | 47.56 (0.45) aA | 16.62 (0.04) cA |
| 10–20 | 26.91 (0.12) aA | 13.59 (0.07) bB | 5.59 (0.09) cB | |
| 20–40 | 10.58 (0.17) aC | 6.52 (0.17) bC | 3.45 (0.04) cC | |
| DOC (mg.kg−1) | 0–10 | 445.12 (7.12) aA | 273.46 (7.21) bA | 154.9 (2.21) cA |
| 10–20 | 367.74 (6.36) aB | 182.21 (5.49) cB | 136.5 (1.58) cB | |
| 20–40 | 199.33 (7.62) aC | 92.29 (0.18) cC | 97.65 (1.15) cC | |
| Soil Variables | Forest Type | Soil Depth | Forest Type × Soil Depth |
|---|---|---|---|
| Soil DOC | 297.8 (<0.001) | 169.5 (<0.001) | 237.6 (<0.001) |
| Soil SOC | 125.1 (<0.001) | 178.2 (<0.001) | 165.6 (<0.001) |
| Soil urease | 119.1 (<0.001) | 33.1 (<0.001) | 81.5 (<0.001) |
| Soil sucrase | 113.1 (<0.001) | 282.2 (<0.001) | 207.6 (<0.001) |
| Soil protease | 11.5 (0.001) | 79.4 (0.07) | 97.4 (<0.001) |
| Soil catalase | 72.28 (0.001) | 51.8 (0.001) | 46.7 (0.06) |
| MBC | 98.4 (<0.001) | 789.3 (<0.001) | 689.1 (<0.001) |
| MBN | 335.4 (<0.001) | 560.4 (<0.001) | 146.3 (<0.001) |
| MBC/SOC | 536.2 (0.01) | 116.1 (0.01) | 2.4 (0.001) |
| MBN/TN | 568.3 (0.01) | 84.1 (0.01) | 118.3 (0.06) |
| Soil Enzymes | Depth (cm) | Forest Types | ||
|---|---|---|---|---|
| NBF a | Mixed | CPF | ||
| Urease | 0–10 | 249.83 (4.07) aA | 247.26 (3.11) aB | 232.86 (1.54) bB |
| (U/g) | 10–20 | 221.56 (3.02) bB | 261.92 (2.35) aA | 216.16 (2.91) cC |
| 20–40 | 230.55 (2.03) bC | 247.26 (2.47) aB | 250.34 (3.88) aA | |
| Sucrase | 0–10 | 78.06 (1.87) cC | 80.32 (3.93) bA | 95.06 (2.68) aA |
| (U/g) | 10–20 | 82.72 (3.23) aB | 70.56 (1.91) bC | 62.92 (2.19) cC |
| 20–40 | 90.68 (2.70) aA | 75.51 (3.78) cB | 80.32 (1.37) bB | |
| Protease | 0–10 | 36.31 (2.30) aA | 32.97 (1.40) bC | 35.46 (0.33) aA |
| (U/g) | 10–20 | 36.09 (1.29) bA | 37.96 (0.49) aA | 35.06 (1.49) bA |
| 20–40 | 31.23 (1.27) bB | 35.28 (1.43) aB | 35.52 (30.56) aA | |
| Catalase | 0–10 | 26.89 (1.52) bB | 28.55 (1.58) aA | 26.19 (0.35) bA |
| (U/g) | 10–20 | 29.69 (0.39) aA | 24.04 (2.75) bB | 23.72 (1.50) cB |
| 20–40 | 29.53 (2.56) aA | 27.01 (1.32) bAB | 27.97 (1.87) bA | |
| pH | C/N | DOC | SOC | AP | AK | MBC | MBN | MBC/ SOC | MBN/ TN | Urease | Sucrase | Protease | Catalase | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | 1 | |||||||||||||
| a C/N | 0.290 | 1 | ||||||||||||
| DOC | 0.042 | 0.798 ** | 1 | |||||||||||
| SOC | 0.129 | 0.249 * | 0.412 * | 1 | ||||||||||
| AP | 0.256 | 0.277 | 0.553 ** | 0.509 | 1 | |||||||||
| AK | 0.365 | 0.946 ** | 0.657 ** | 0.215 * | 0.132 | 1 | ||||||||
| MBC | 0.108 | 0.791 ** | 0.684 ** | 0.731 ** | 0.150 | 0.842 ** | 1 | |||||||
| MBN | 0.341 | 0.570 ** | 0.723 ** | 0.467 ** | 0.295 | 0.572 ** | 0.846 ** | 1 | ||||||
| MBC/ SOC | 0.483 * | 0.209 | 0.176 | 0.414 ** | 0.034 | 0.094 | 0.336 | 0.248 | 1 | |||||
| MBN/ TN | 0.668 ** | 0.052 | 0.209 | 0.349 | 0.247 | 0.057 | 0.371 | 0.698 ** | 0.477 * | 1 | ||||
| Urease | 0.306 | 0.121 | 0.506 ** | 0.471 | 0.437 * | 0.084 | 0.001 | 0.423 * | 0.344 | 0.444 * | 1 | |||
| Sucrase | 0.331 | 0.368 | 0.361 | 0.124 | 0.072 | 0.168 | 0.212 | 0.093 | 0.147 | 0.002 | 0.091 | 1 | ||
| Protease | 0.093 | 0.168 | 0.256 | 0.348 | 0.445 * | 0.085 | 0.099 | 0.141 | 0.018 | 0.103 | 0.315 | 0.405 * | 1 | |
| Catalase | 0.245 | 0.510 ** | 0.344 | 0.356 | 0.294 | 0.340 | 0.099 * | 0.078 * | 0.360 | 0.139 | 0.116 | 0.625 ** | 0.549 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, T.H.; Chen, X.; Shakoor, A.; Rashid, M.H.U.; Kumar, U.; Alhomrani, M.; Alamri, A.S.; Ravindran, B.; Yan, W. Unraveling the Importance of Forest Structure and Composition Driving Soil Microbial and Enzymatic Responses in the Subtropical Forest Soils. Forests 2022, 13, 1535. https://doi.org/10.3390/f13101535
Farooq TH, Chen X, Shakoor A, Rashid MHU, Kumar U, Alhomrani M, Alamri AS, Ravindran B, Yan W. Unraveling the Importance of Forest Structure and Composition Driving Soil Microbial and Enzymatic Responses in the Subtropical Forest Soils. Forests. 2022; 13(10):1535. https://doi.org/10.3390/f13101535
Chicago/Turabian StyleFarooq, Taimoor Hassan, Xiaoyong Chen, Awais Shakoor, Muhammad Haroon U. Rashid, Uttam Kumar, Majid Alhomrani, Abdulhakeem S. Alamri, Balasubramani Ravindran, and Wende Yan. 2022. "Unraveling the Importance of Forest Structure and Composition Driving Soil Microbial and Enzymatic Responses in the Subtropical Forest Soils" Forests 13, no. 10: 1535. https://doi.org/10.3390/f13101535