
Response of Temperate Forest Ecosystems under Decreased Nitrogen Deposition: Research Challenges and Opportunities


Abstract
:1. Introduction
2. Research at Fernow Experimental Forest
2.1. Background
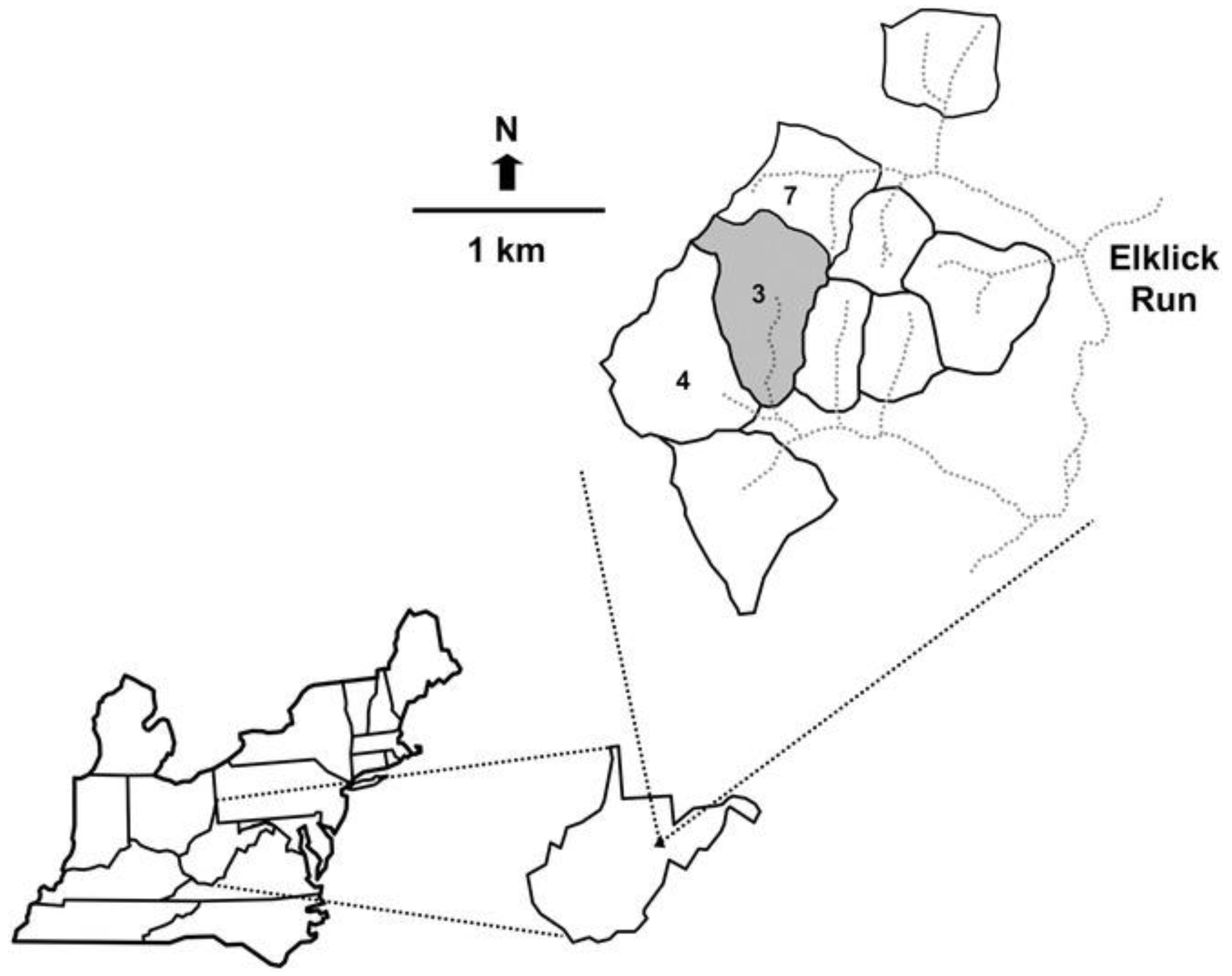
2.2. Site Description
2.3. Field Design
2.4. Findings
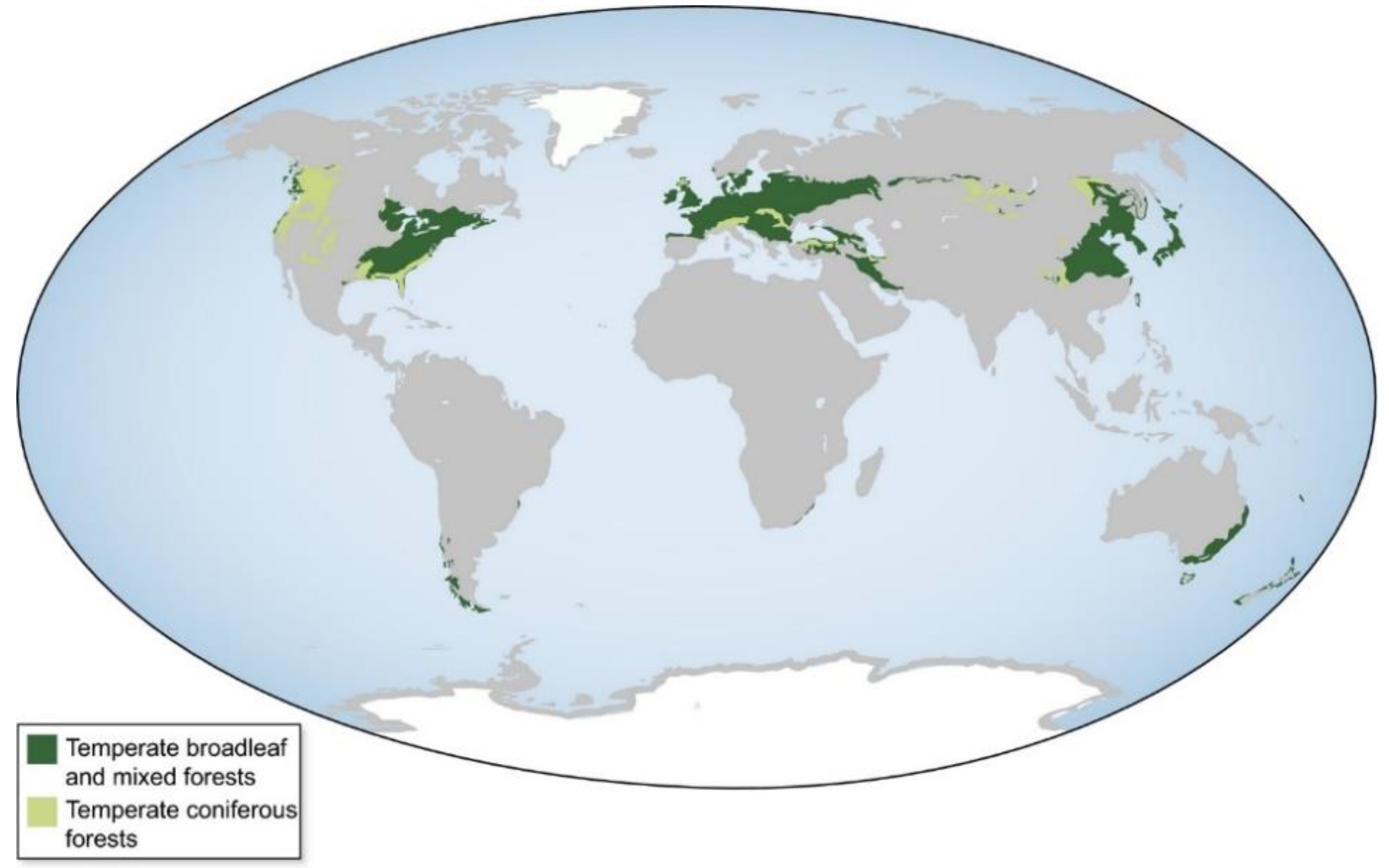
3. Response of Terrestrial Ecosystems to Decreased N Deposition
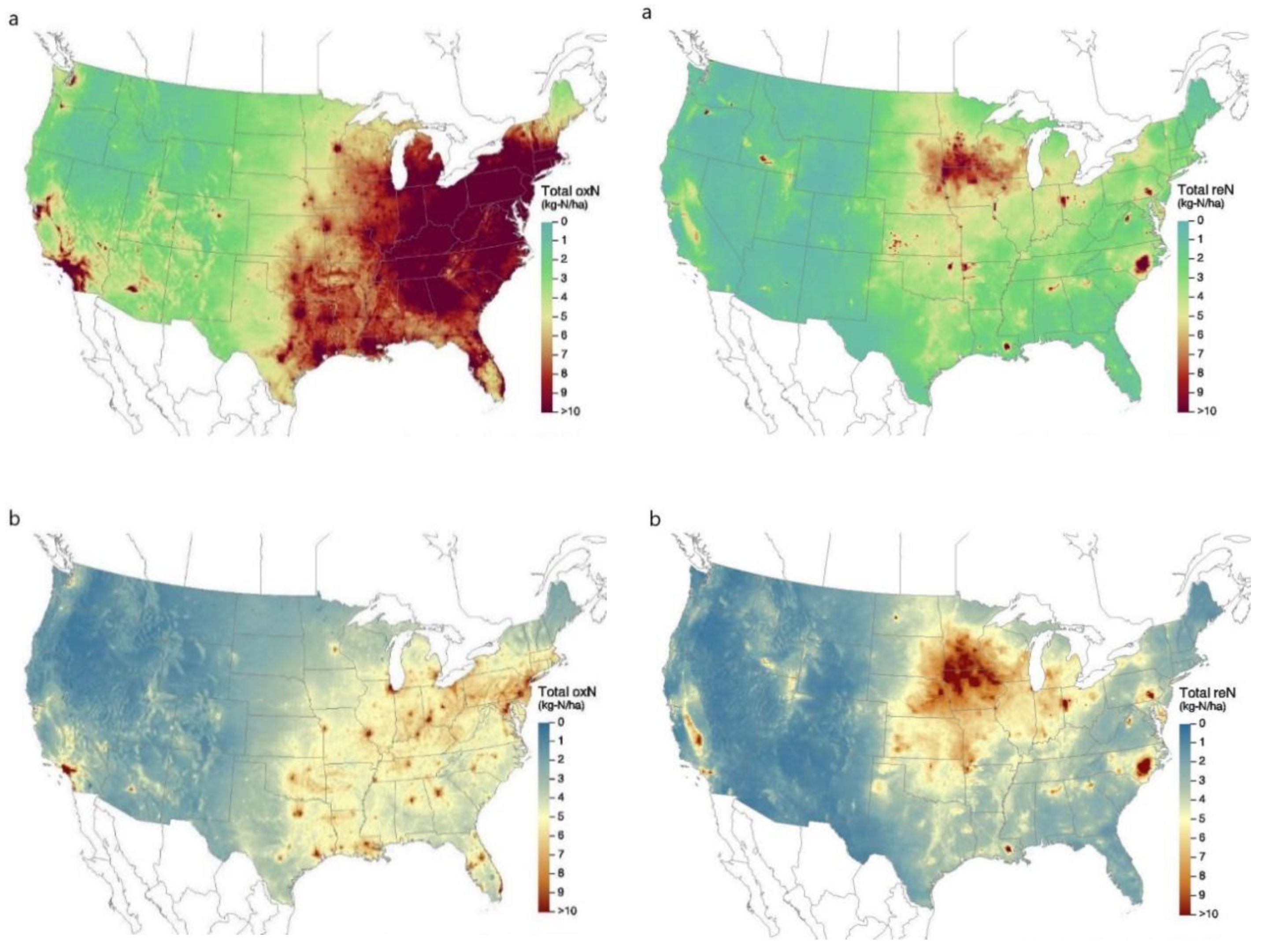
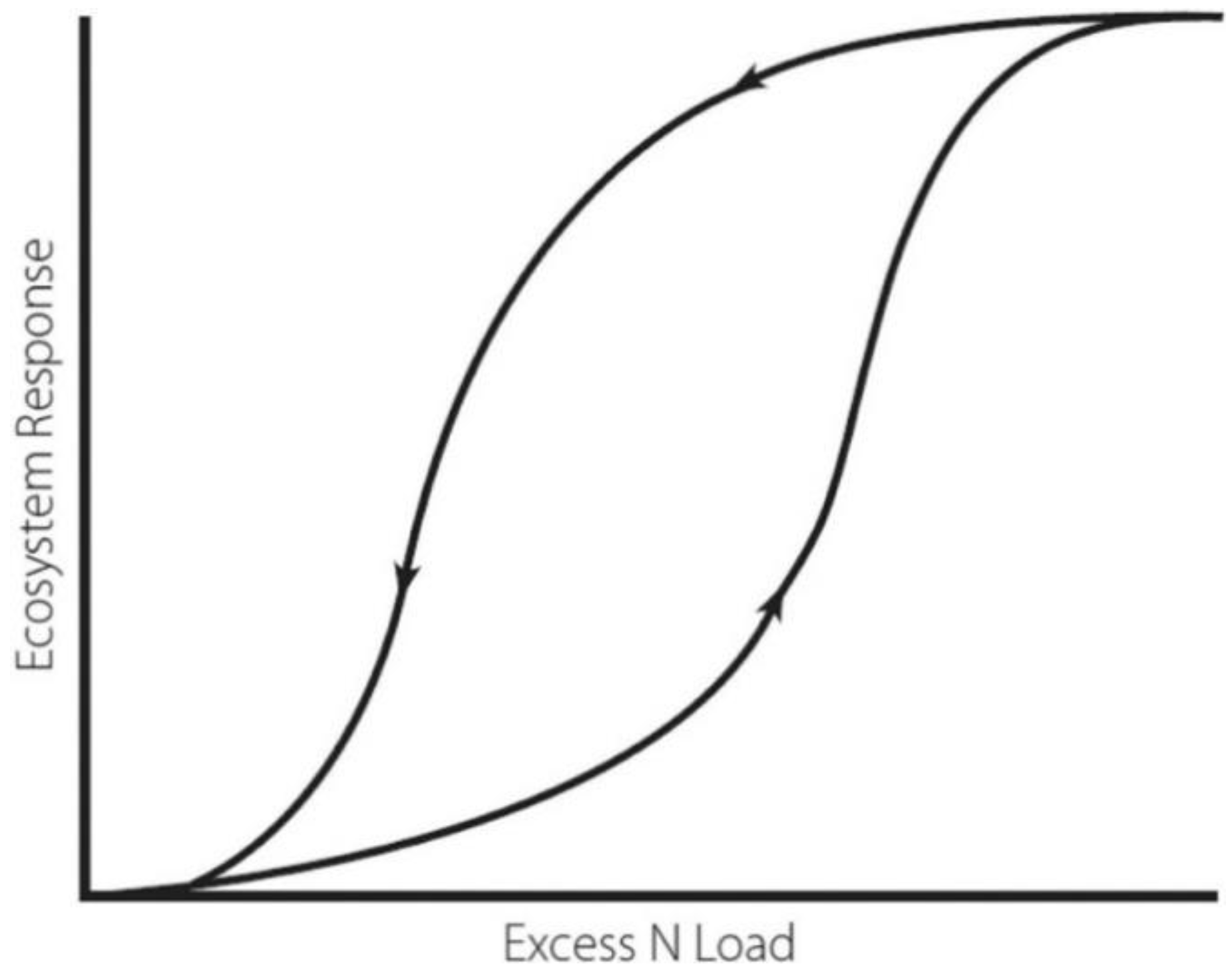
4. Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horii, C.V.; Munger, J.W.; Wofsy, S.C.; Zahniser, M.; Nelson, D.; McManus, J.B. Atmospheric reactive nitrogen concentration and flux budgets at a Northeastern U.S. forest site. Agric. For. Meteorol. 2005, 133, 210–225. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity effects of terrestrial ecosystems: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvertown, J.; Poulton, P.; Johnson, A.E.; Edwards, G.; Heard, M.; Biss, P.M. The Park Grass Experiment 1865-2006: Its contribution to ecology. J. Ecol. 2006, 94, 801–814. [Google Scholar] [CrossRef]
- De Schrijver, A.; de Frenne, P.; Ampoorter, E.; van Nevel, L.; Demey, A.; Wuyts, K.; Verheyen, K. Cumulative nitrogen inputs drives species loss in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2011, 20, 803–816. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Römermann, C.; Pillar, V.D.; Kudernatsch, T.; Fischer, A. High functional diversity is related to high nitrogen availability in a deciduous forest—Evidence from a functional trait approach. Folia Geobot. 2010, 45, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt-Römermann, M.; Baeten, L.; Craven, D.; De Frenne, P.; Hédl, R.; Lenoir, J.; Bert, D.; Brunet, J.; Chudomelová, M.; Decocq, G.; et al. Drivers of temporal changes in temperate forest plant diversity vary across spatial scales. Glob. Chang. Biol. 2015, 21, 3726–3737. [Google Scholar] [CrossRef]
- Verheyen, K.; Baeten, L.; De Frenne, P.; Bernhardt-Römermann, M.; Brunet, J.; Cornelis, J.; Decocq, G.; Dierschke, H.; Eriksson, O.; Hédl, R.; et al. Driving factors behind the eutrophication signal in understorey plant communities of deciduous temperate forests. J. Ecol. 2012, 100, 352–365. [Google Scholar] [CrossRef]
- Dirnböck, T.; Grandin, U.; Bernhardt-Römermann, M.; Beudert, B.; Canullo, R.; Forsius, M.; Grabner, M.-T.; Holmberg, M.; Kleemola, S.; Lundin, L.; et al. Forest floor vegetation response to nitrogen deposition in Europe. Glob. Chang. Biol. 2014, 20, 429–440. [Google Scholar] [CrossRef]
- Ferretti, M.; Marchetto, A.; Arisci, S.; Bussotti, F.; Calderisi, M.; Carnicelli, S.; Cecchini, G.; Fabbio, G.; Bertini, G.; Matteucci, G.; et al. On the tracks of Nitrogen deposition effects on temperate forests at their southern European range–an observational study from Italy. Glob. Chang. Biol. 2014, 20, 3423–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilliam, F.S.; Welch, N.T.; Phillips, A.H.; Billmyer, J.H.; Peterjohn, W.T.; Fowler, Z.K.; Walter, C.A.; Burnham, M.B.; May, J.D.; Adams, M.B. Twenty-five year response of the herbaceous layer of a temperate hardwood forest to elevated nitrogen deposition. Ecosphere 2016, 7, e01250. [Google Scholar] [CrossRef] [Green Version]
- Eastman, B.A.; Adams, M.B.; Brzotek, E.R.; Burnham, M.B.; Carrara, J.E.; Kelly, C.; McNeil, B.E.; Walter, C.A.; Peterjohn, W.T. Altered plant carbon partitioning enhanced forest ecosystem carbon storage after 25 years of nitrogen additions. New Phytol. 2021, 230, 1435–1448. [Google Scholar] [CrossRef]
- Patel, K.F.; Fernandez, I.J.; Nelson, S.J.; Malcom, J.; Norton, S.A. Contrasting stream nitrate and sulfate response to recovery from experimental watershed acidification. Biogeochemistry 2020, 151, 127–138. [Google Scholar] [CrossRef]
- Ågren, G.I.; Bosatta, E. Nitrogen saturation of terrestrial ecosystems. Environ. Pollut. 1988, 54, 185–197. [Google Scholar] [CrossRef]
- Skeffington, R.A.; Wilson, E.J. Excess nitrogen deposition: Issues for consideration. Environ. Pollut. 1988, 54, 159–184. [Google Scholar] [CrossRef]
- Aber, J.D.; McDowell, W.H.; Nadelhoffer, K.J.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.G.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen saturation in temperate forest ecosystems: Hypotheses revisited. BioScience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Aber, J.D.; Nadelhoffer, K.J.; Steudler, P.; Melillo, J.M. Nitrogen saturation in northern forest ecosystems—Hypotheses and implications. BioScience 1998, 39, 378–386. [Google Scholar] [CrossRef]
- Gilliam, F.S. Forest ecosystems of temperate climatic regions: From ancient use to climate change. New Phytol. 2016, 212, 871–887. [Google Scholar] [CrossRef]
- Lovett, G.M.; Goodale, C.L. A new conceptual model of nitrogen saturation based on experimental nitrogen addition to an oak forest. Ecosystems 2011, 14, 615–631. [Google Scholar] [CrossRef]
- Peterjohn, W.T.; Adams, M.B.; Gilliam, F.S. Symptoms of nitrogen saturation in two central Appalachian hardwood forests. Biogeochemistry 1996, 35, 507–522. [Google Scholar] [CrossRef]
- Gilliam, F.S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.A.M.; Anthony, M.A.; Pec, G.J.; Trocha, L.K.; Trzebny, A.; Geyer, K.M.; van Diepen, L.T.A.; Frey, S.D. Fungal community structure and function shifts with atmospheric nitrogen deposition. Glob. Chang. Biol. 2021, 27, 1349–1364. [Google Scholar] [CrossRef]
- Thomas, R.Q.; Canham, C.D.; Weathers, K.C.; Goodale, C.L. Increased tree carbon storage in response to nitrogen deposition in the US. Nat. Geosci. 2010, 3, 13–17. [Google Scholar] [CrossRef]
- Peel, J.L.; Haeuber, R.; Garcia, V.; Russell, A.G.; Neas, L. Impact of nitrogen and climate change interactions on ambient air pollution and human health. Biogeochemistry 2013, 114, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Lloret, J.; Valiela, I. Unprecedented decrease in deposition of nitrogen oxides over North America: The relative effects of emission controls and prevailing air-mass trajectories. Biogeochemistry 2016, 129, 165–180. [Google Scholar] [CrossRef]
- Du, E. Rise and fall of nitrogen deposition in the United States. Proc. Natl. Acad. Sci. USA 2016, 113, E3594–E3595. [Google Scholar] [CrossRef] [Green Version]
- Warner, J.X.; Dickerson, R.R.; Wei, Z.; Strow, L.L.; Wang, Y.; Liang, Q. Increased atmospheric ammonia over the world’s major agricultural areas detected from space. Geophys. Res. Lett. 2017, 44, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, F.S.; Burns, D.A.; Driscoll, C.T.; Frey, S.D.; Lovett, G.M.; Watmough, S.A. Decreased atmospheric nitrogen deposition in eastern North America: Predicted responses of forest ecosystems. Environ. Pollut. 2019, 244, 560–574. [Google Scholar] [CrossRef]
- Högberg, P.; Johannisson, C.; Yarwood, S.; Callesen, I.; Näsholm, T.; Myrold, D.D.; Högberg, M.N. Recovery of ectomycorrhiza after ‘nitrogen saturation’ of a conifer forest. New Phytol. 2011, 189, 515–525. [Google Scholar] [CrossRef]
- Rowe, E.C.; Jones, L.; Dise, N.B.; Evans, C.D.; Mills, G.; Hall, J.; Stevens, C.J.; Mitchell, R.J.; Field, C.; Caporn, S.J.M.; et al. Metrics for evaluation the ecological benefits of decreased nitrogen deposition. Biol. Conserv. 2014, 212, 454–463. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.J. How long do ecosystems take to recover from atmospheric nitrogen deposition? Biol. Conserv. 2016, 200, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Groffman, P.M.; Driscoll, C.T.; Durán, J.; Campbell, J.L.; Christenson, L.M.; Fahey, T.J.; Fisk, M.C.; Fuss, C.; Likens, G.E.; Lovett, G.; et al. Nitrogen oligotrophication in northern hardwood forests. Biogeochemistry 2018, 141, 523–539. [Google Scholar] [CrossRef]
- Schmitz, A.; Sanders, T.G.M.; Bolte, A.; Bussotti, F.; Dirnböck, T.; Johnson, J.; Peñuelas, J.; Pollastrini, M.; Prescher, A.-K.; Sardans, J.; et al. Responses of forest ecosystems in Europe to decreasing nitrogen deposition. Environ. Pollut. 2019, 244, 980–994. [Google Scholar] [CrossRef]
- Adams, M.B.; DeWalle, D.R.; Hom, J. The Fernow Watershed Acidification Study; Environmental Pollution Series 11; Springer: New York, NY, USA, 2006. [Google Scholar]
- Weitzman, S. Five Years of Research on the Fernow Experimental Forest; Station Paper NE-61; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Upper Darby, PA, USA, 1953.
- Gilliam, F.S.; Hockenberry, A.W.; Adams, M.B. Effects of atmospheric nitrogen deposition on the herbaceous layer of a central Appalachian hardwood forest. J. Torrey Bot. Soc. 2006, 133, 240–254. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, F.S.; Roberts, M.R. The Herbaceous Layer in Forests of Eastern North America; Oxford University Press, Inc.: New York, NY, USA, 2003. [Google Scholar]
- Gilliam, F.S. The Herbaceous Layer in Forests of Eastern North America, 2nd ed.; Oxford University Press, Inc.: New York, NY, USA, 2014. [Google Scholar]
- Gilliam, F.S.; Turrill, N.L.; Aulick, S.D.; Evans, D.K.; Adams, M.B. Herbaceous layer and soil response to experimental acidification in a Central Appalachian hardwood forest. J. Environ. Qual. 1994, 23, 835–844. [Google Scholar] [CrossRef] [Green Version]
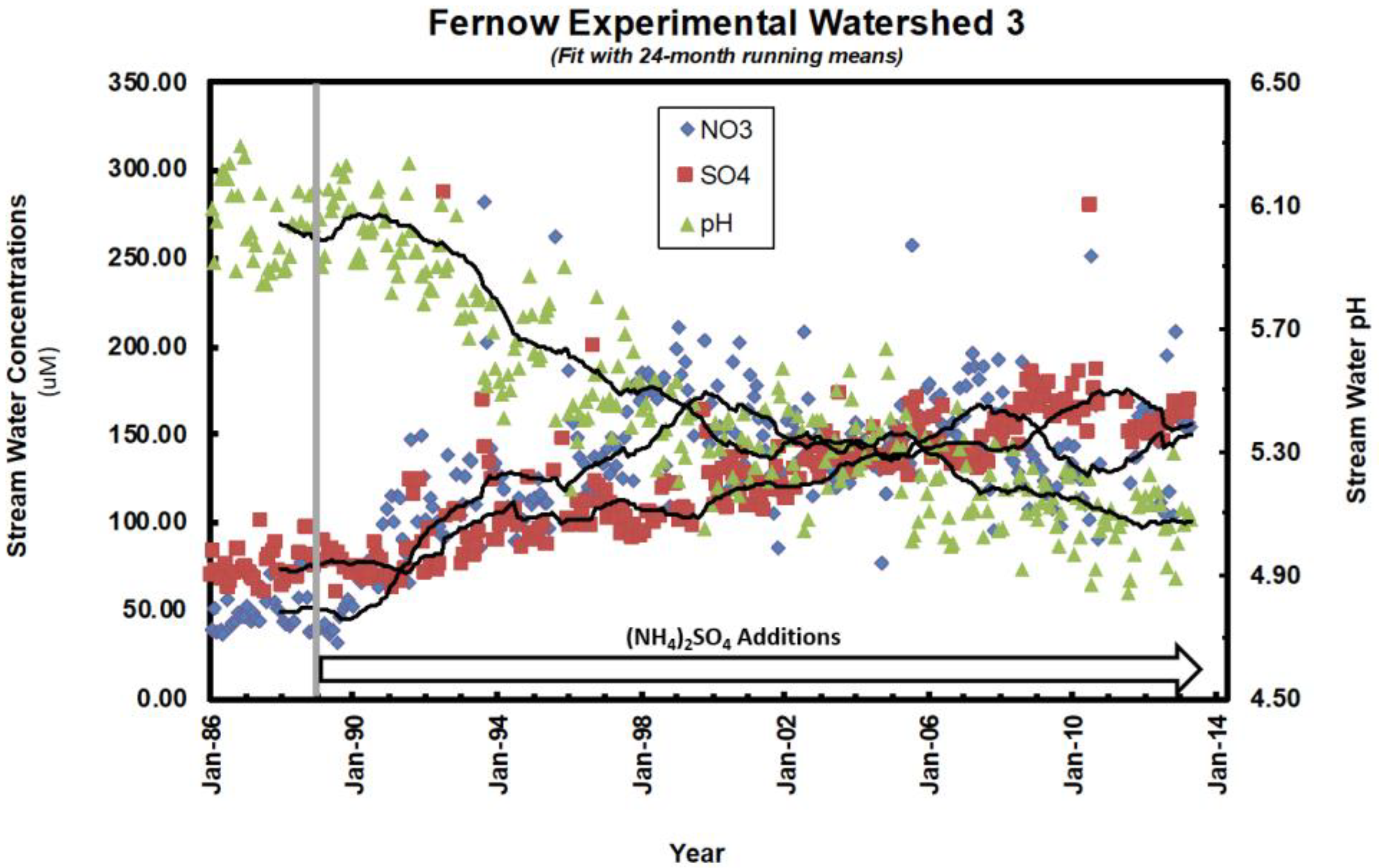
- Gilliam, F.S.; Adams, M.B.; Yurish, B.M. Ecosystem nutrient response to chronic nitrogen inputs at Fernow Experimental Forest, West Virginia. Can. J. For. Res. 1996, 26, 196–205. [Google Scholar] [CrossRef]
- Edwards, P.J.; Kochenderfer, J.N.; Coble, D.W.; Adams, M.B. Soil leachate responses during 10 years of induced whole-watershed acidification. Water Air Soil Pollut. 2002, 140, 99–118. [Google Scholar] [CrossRef]
- Edwards, P.J.; Wood, F.; Kochenderfer, J.N. Baseflow and peakflow chemical responses to experimental applications of ammonium sulphate to forested watersheds in north-central West Virginia, USA. Hydrol. Process. 2002, 16, 2287–2310. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Walter, C.A.; Adams, M.B.; Peterjohn, W.T. Nitrogen (N) dynamics in the mineral soil of a Central Appalachian hardwood forest during a quarter century of whole-watershed N additions. Ecosystems 2018, 21, 1489–1504. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Adams, M.B.; Peterjohn, W.T. Response of soil fertility to 25 years of experimental acidification in a temperate hardwood forest. J. Environ. Qual. 2020, 49, 961–972. [Google Scholar] [CrossRef]
- Adams, M.B.; Angradi, T.R. Decomposition and nutrient dynamics of hardwood leaf litter in the Fernow Whole-Watershed Acidification Experiment. For. Ecol. Manag. 1996, 83, 61–69. [Google Scholar] [CrossRef]
- Carrara, J.E.; Walter, C.A.; Hawkins, J.S.; Peterjohn, W.T.; Averill, C.; Brzostek, E.R. Interactions among plants, bacteria, and fungi reduce extracellular enzyme activities under long-term N fertilization. Glob. Chang. Biol. 2017, 24, 2721–2734. [Google Scholar] [CrossRef] [PubMed]
- Burnham, M.B.; Cumming, J.R.; Adams, M.B.; Peterjohn, W.T. Soluble soil aluminum alters the relative uptake of mineral nitrogen forms by six mature temperate broadleaf tree species: Possible implications for watershed nitrate retention. Oecologia 2017, 185, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Venterea, R.T.; Groffman, P.M.; Castro, M.S.; Verchot, L.V.; Fernandez, I.J.; Adams, M.B. Soil emissions of nitric oxide in two forest watersheds subjected to elevated N inputs. For. Ecol. Manag. 2004, 196, 335–349. [Google Scholar] [CrossRef]
- Peterjohn, W.T.; McGervey, R.J.; Sexstone, A.J.; Christ, M.J.; Foster, C.J.; Adams, M.B. Nitrous oxide production in two forested watersheds exhibiting symptoms of nitrogen saturation. Can. J. For. Res. 1998, 28, 1723–1732. [Google Scholar] [CrossRef]
- Gress, S.E.; Nichols, T.D.; Northcraft, C.C.; Peterjohn, W.T. Nutrient limitation in soils exhibiting differing nitrogen availabilities: What lies beyond nitrogen saturation. Ecology 2007, 88, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, F.S.; Billmyer, J.H.; Walter, C.A.; Peterjohn, W.T. Effects of excess nitrogen on biogeochemistry of a temperate hardwood forest: Evidence of nutrient redistribution by a forest understory species. Atmos. Environ. 2016, 146, 261–270. [Google Scholar] [CrossRef] [Green Version]
- DeWalle, D.R.; Tepp, J.S.; Swistock, B.R.; Sharpe, W.E.; Edwards, P.J. Tree-ring cation response to experimental watershed acidification in West Virginia and Maine. J. Environ. Qual. 1999, 28, 299–309. [Google Scholar] [CrossRef]
- May, J.D.; Burdette, S.B.; Gilliam, F.S.; Adams, M.B. Interspecific divergence in foliar nutrient dynamics and stem growth in a temperate forest in response to chronic nitrogen inputs. Can. J. For. Res. 2005, 35, 1023–1030. [Google Scholar] [CrossRef]
- Malcomb, J.D.; Scanlon, T.M.; Epstein, H.E.; Druckenbrod, D.L.; Vadeboncoeur, M.A.; Lanning, M.; Adams, M.B.; Wang, L. Assessing temperate forest growth and climate sensitivity in response to a long-term whole-watershed acidification experiment. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005560. [Google Scholar] [CrossRef]
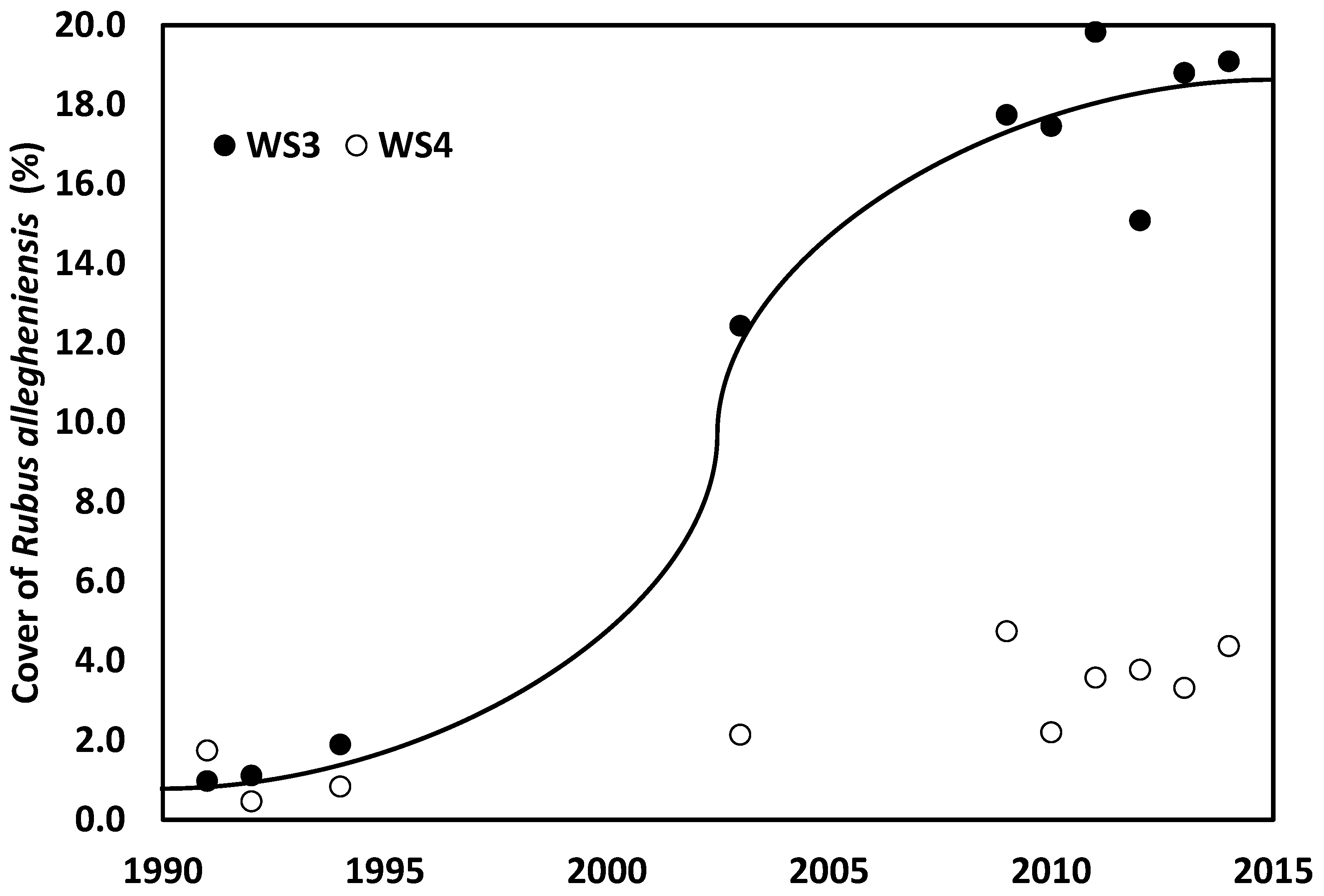
- Walter, C.A.; Raiff, D.T.; Burnham, M.B.; Gilliam, F.S.; Adams, M.B.; Peterjohn, W.T. Nitrogen fertilization interacts with light to increase Rubus spp. cover in a temperate forest. Plant Ecol. 2016, 217, 421–430. [Google Scholar] [CrossRef]
- Boxman, A.W.; van der Ven, P.J.M.; Roelofs, J.G.M. Ecosystem recovery after a decrease in nitrogen input to a Scots pine stand at Ysselsteyn, the Netherlands. For. Ecol. Manag. 1998, 101, 155–163. [Google Scholar] [CrossRef]
- Strengbom, J.; Nordin, A.; Nasholm, T.; Ericson, L. Slow recovery of boreal forest ecosystem following decreased nitrogen input. Funct. Ecol. 2001, 15, 451–457. [Google Scholar] [CrossRef]
- Booth, M.S.; Stark, J.M.; Rastetter, E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol. Monogr. 2005, 75, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, F.S.; May, J.D.; Adams, M.B. Response of foliar nutrients of Rubus allegheniensis to nutrient amendments in a central Appalachian hardwood forest. For. Ecol. Mang. 2018, 411, 101–107. [Google Scholar] [CrossRef]
- Wen, Z.; Xu, W.; Li, Q.; Han, M.; Tang, A.; Zhang, Y.; Luo, X.; Shen, J.; Wang, W.; Li, K.; et al. Changes of nitrogen deposition in China from 1980 to 2018. Environ. Int. 2020, 144, 106022. [Google Scholar] [CrossRef]
- Nieland, M.A.; Moley, P.; Hanschu, J.; Zeglin, L.H. Differential resilience of soil microbes and ecosystem functions following cessation of long-term fertilization. Ecosystems 2021. [Google Scholar] [CrossRef]
- Newcomer, M.E.; Bouskill, N.J.; Wainwright, H.; Maavara, T.; Arora, B.; Siirila-Woodburn, E.R.; Dwivedi, D.; Williams, K.H.; Steefel, C.; Hubbard, S.S. Hysteresis patterns of watershed nitrogen retention and loss over the past 50 years in United States hydrological basins. Glob. Biogeochem. Cycles 2021, 35, e2020GB006777. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Yr | Focus | Findings |
|---|---|---|---|
| Gilliam et al. (1994) [39] | 3 | herb layer/soil nutrients | no significant differences between WS3, WS4, and WS7 |
| Gilliam et al. (1996) [40] | 4 | N mineralization, foliar nutrients | no differences for N mineralization |
| Peterjohn et al. (1996) [20] | 4 | symptoms of N saturation | clear evidence of N saturation on untreated WS4 |
| Gilliam et al. (2006) [36] | 5 | herb layer communities | no differences in composition and diversity |
| Edwards et al. (2002) [41] | 8 | soil solution chemistry | higher NO3− and cation concentrations on WS3 |
| Edwards et al. (2002) [42] | 8 | stream chemistry | increases in NO3−, cations, and acidity, following ~2 yr lag |
| Gilliam et al. (2016) [11] | 25 | herb layer composition/diversity | N alters herb layer composition, decreases species diversity |
| Gilliam et al. (2018) [43] | 25 | soil N mineralization/nitrification | no watershed differences, increased homogeneity on WS3 |
| Gilliam et al. (2020) [44] | 25 | soil fertility | significant decreases in base cations and soil pH |
| Eastman et al. (2021) [12] | 30 | long-term carbon and N budgets | added N leads to greater C storage in vegetation and soil |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilliam, F.S. Response of Temperate Forest Ecosystems under Decreased Nitrogen Deposition: Research Challenges and Opportunities. Forests 2021, 12, 509. https://doi.org/10.3390/f12040509
Gilliam FS. Response of Temperate Forest Ecosystems under Decreased Nitrogen Deposition: Research Challenges and Opportunities. Forests. 2021; 12(4):509. https://doi.org/10.3390/f12040509
Chicago/Turabian StyleGilliam, Frank S. 2021. "Response of Temperate Forest Ecosystems under Decreased Nitrogen Deposition: Research Challenges and Opportunities" Forests 12, no. 4: 509. https://doi.org/10.3390/f12040509