
Cambial Activity and Phenology in Three Understory Species along an Altitude Gradient in Mexico


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Environmental Factors
2.3. Phenology, Leaf Area and Growth
2.4. Cambial Activity
2.5. Statistical Analyses
3. Results
3.1. Cambial Activity
3.2. Phenology
3.3. Effect of Environmental Variables on Vascular Cambium and Phenology
4. Discussion
4.1. Seasonality
4.2. Phenology
4.3. Vascular Cambium
4.4. Vascular Cambium, Phenology and Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Schmitt, U.; Möller, R.; Eckstein, D. Seasonal wood formation dynamics of beech (Fagus sylvatica L.) and black locust (Robinia pseudoacacia L.) as determined by the ‘‘pinning’’ technique. J. Appl. Bot. 2000, 74, 10–16. [Google Scholar]
- Čufar, K.; Prislan, P.; De Luis, M.; Gričar, J. Tree-ring variation, wood formation and phenology of beech (Fagus sylvatica) from a representative site in Slovenia, SE Central Europe. Trees 2008, 22, 749–758. [Google Scholar] [CrossRef]
- Rao, S.K.; Rajput, K.S. Relationship between seasonal cambial activity, development of xylem and phenology in Azadirachta indica growing in different forests of Gujarat State. Ann. For. Sci. 2001, 58, 691–698. [Google Scholar] [CrossRef] [Green Version]
- Marcati, C.R.; Angyalossy, V.; Evert, R.F. Seasonal variation in wood formation of Cedrela fissilis (Meliaceae). IAWA J. 2006, 27, 199–211. [Google Scholar] [CrossRef]
- Yañéz-Espinosa, L.; Terrazas, T.; López-Mata, L. Phenology and radial stem growth periodicity in evergreen subtropical rainforest trees. IAWA J. 2010, 31, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.-G.; Delauriers, A.; Rossi, S. Xylem formation can be modeled statistically as a function of primary growth and cambium activity. New Phytol. 2014, 203, 831–841. [Google Scholar] [CrossRef]
- Morin, X.; Augspurger, C.; Chuine, I. Process-based modeling of species’ distribution: What limits temperate tree species’ range boundaries? Ecology 2007, 88, 2280–2291. [Google Scholar] [CrossRef] [PubMed]
- Borchert, R.; Renner, S.S.; Calle, Z.; Navarrete, D.; Tye, A.; Gautier, L.; Spichiger, R.; von Hildebrand, P. Photoperiodic induction of synchronous flowering near the Equator. Nature 2005, 433, 627–629. [Google Scholar] [CrossRef]
- Worbes, M. How to measure growth dynamics in tropical trees. A review. IAWA J. 1995, 16, 337–351. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Begin, Y. Cellular phenology of annual ring formation of Abies balsamea in the Quebec boreal forest (Canada). Can. J. Forest Res. 2003, 33, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Cuny, H.E.; Rathgeber, C.B.K.; Lebourgeois, F.; Fortin, M.; Fournier, M. Life strategies in intra-annual dynamics of wood formation: Example of three conifer species in a temperate forest in north-east France. Tree Physiol. 2012, 32, 612–625. [Google Scholar] [CrossRef] [Green Version]
- Yañéz-Espinosa, L.; Terrazas, T.; López-Mata, L. Integrated analysis of tropical trees growth: A multivariate approach. Ann. Bot. 2006, 98, 637–645. [Google Scholar] [CrossRef]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Yamane, K.; Islam, M.D.A.; Oribe, Y.; Ko, J.H.; Jin, H.O.; Funada, R. A rapid decrease in temperature induces latewood formation in artificially reactivated cambium of conifer stems. Ann. Bot. 2012, 110, 875–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deslauriers, A.; Rossi, S.; Anfodillo, T.; Saracino, A. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Eckstein, D.; Shao, X. Seasonal cambial activity of relict Chinese pine at the northern limit of its natural distribution in North China, exploratory results. IAWA J. 2009, 30, 371–378. [Google Scholar] [CrossRef]
- Cornelius, C.; Estrella, N.; Franz, H.; Menzel, A. Linking altitudinal gradients and temperature responses of plant phenology in the Bavarian Alps. Plant Biol. 2012, 15, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gou, X.; Pederson, N.; Zhang, F.; Niu, H.; Zhao, S.; Wang, F. Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol. 2018, 38, 840–852. [Google Scholar] [CrossRef]
- Moser, L.; Fonti, P.; Büntgen, U.; Esper, J.; Luterbacher, J.; Franzen, J.; Frank, D. Timing and duration of European larch growing season along altitudinal gradients in the Swiss Alps. Tree Physiol. 2010, 30, 225–233. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gričar, J.; Gruber, A.; King, G.M.; et al. A meta-analysis of cambium phenology and growth: Linear and non-linear patterns in conifers of the northern hemisphere. Ann. Bot. 2013, 112, 1921–1930. [Google Scholar] [CrossRef]
- Aljaro, M.E.; Ávila, G.; Hoffman, A.; Kummerow, J. The annual rhythm of cambial activity in two woody species of the Chilean “matorral”. Amer. J. Bot. 1972, 59, 879–885. [Google Scholar] [CrossRef]
- Avila, G.; Aljaro, M.E.; Araya, S.; Montenegro, G.; Kummerow, J. The seasonal cambium activity of Chilean and Californian shrubs. Amer. J. Bot. 1975, 62, 473–478. [Google Scholar] [CrossRef]
- Li, X.; Rossi, S.; Liang, E.; Camarero, J.J. Temperature thresholds for the onset of xylogenesis in alpine shrubs on the Tibetan plateau. Trees 2016, 30, 2091–2099. [Google Scholar] [CrossRef]
- Lara, N.O.T.; Marcati, C.R. Cambial dormancy lasts 9 months in a tropical evergreen species. Trees 2016, 30, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Gilliam, F.S. The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Muller, R.N. Nutrient relations of the herbaceous layer in deciduous forest ecosystems. In The Herbaceous Layer in Forests of Eastern North America; Gilliam, F.S., Roberts, M.R., Eds.; Oxford University Press: New York, NY, USA, 2003; pp. 15–37. [Google Scholar]
- Donnelly, A.; Yu, R. Temperate deciduous shrub phenology: The overlooked forest layer. Int. J. Biometeorol. 2021, 65, 343–355. [Google Scholar] [CrossRef]
- Su, X.; Wang, M.; Huang, Z.; Fu, S.; Chen, H.Y.H. Forest understorey vegetation: Colonization and the availability and heterogeneity of resources. Forests 2019, 10, 944. [Google Scholar] [CrossRef] [Green Version]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; University Press: Oxford, UK, 1934. [Google Scholar]
- Yazdanshenas, H.; Kalagar, M.; Toularoud, M.M. Understory plant species diversity of Asalem’s forests, northern Iran. For. Res. Eng. Int. J. 2019, 3, 56–62. [Google Scholar]
- Cortés-Flores, J.; Cornejo-Tenorio, G.; Ibarra-Manríquez, G. Flowering phenology and pollination syndromes in species with different growth forms in a Neotropical temperate forest of Mexico. Botany 2015, 93, 361–367. [Google Scholar] [CrossRef]
- Bonilla-Valencia, L.; Martínez-Orea, Y.; Castillo-Argüero, S.; Barajas-Guzmán, G.; Romero-Romero, M.A.; Díaz-López, E.T. Reproductive phenology of understory species in an Abies religiosa (Pinaceae) forest in the Magdalena River Basin, Mexico City. J. Torrey Bot. Soc. 2017, 144, 313–327. [Google Scholar] [CrossRef]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen, 2nd ed.; Instituto de Geografía, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2004; Serie Libros No. 6. [Google Scholar]
- Morgado-González, G.; Gómez-Guerrero, A.; Villanueva-Díaz, J.; Terrazas, T.; Ramírez-Herrera, C.; Hernández de la Rosa, P. Densidad de la madera de Pinus hartwegii Lind. en dos niveles altitudinales y de exposición. Agrociencia 2019, 53, 645–660. [Google Scholar]
- Sánchez-González, A.; López-Mata, L. Plant species richness and diversity along an altitudinal gradient in the Sierra Nevada, Mexico. Divers. Distrib. 2005, 11, 567–575. [Google Scholar] [CrossRef]
- Jiménez-Noriega, M.S.; Terrazas, T.; López-Mata, L.; Sánchez-González, A.; Vibrans, H. Anatomical variation of five plant species along an elevation gradient in Mexico City basin within the Trans-Mexican Volcanic Belt, Mexico. J. Mt. Sci. 2017, 14, 218–2199. [Google Scholar] [CrossRef]
- Rich, P.M. Characterizing plant canopies with hemispherical photographs. Remote Sens. Rev. 1990, 5, 13–29. [Google Scholar] [CrossRef]
- Schleppi, P.; Paquette, A. Solar radiation in forests: Theory for hemispherical photography. In Hemispherical Photography in Forest Science: Theory, Methods, Applications. Managing Forest Ecoystems; Fournier, R.A., Hall, R.J., Eds.; Springer Science+Business Media: Berlin/Heidelberg, Germany, 2017; Volume 28. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants: BBCH Monograph; Open Agrar Repositorium: Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- Fournier, L.A. Un método cuantitativo para la medición de características fenológicas en árboles. Turrialba 1974, 24, 422–423. [Google Scholar]
- Hunt, R. Plant Growth Analysis. Studies in Biology; Edward Arnold: London, UK, 1978. [Google Scholar]
- Ruzin, S.E. Plant Microtechnique and Microscopy; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Zarlavsky, G.E. Histología Vegetal. Técnicas Simples y Complejas; Sociedad Argentina de Botánica: Buenos Aires, Argentina, 2014. [Google Scholar]
- Gričar, J.; Zupančič, M.; Čufar, K.; Koch, G.; Schmitt, U.; Oven, P. Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). Ann. Bot. 2006, 97, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Angyalossy, V.; Pace, M.R.; Marcati, C.R.; Evert, R.F. Phloem development, growth markers, and sieve-tube longevity in two Neotropical trees. IAWA J. 2021, 44, 31–49. [Google Scholar] [CrossRef]
- Der, G.; Everitt, B.S. A Handbook of Statistical Analyses Using SAS, 2nd ed.; Chapman & Hall/CRC: BocaW E Raton, FL, USA, 2002. [Google Scholar]
- SAS. SAS/STAT User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2008; Release 9.2. [Google Scholar]
- Caffarra, A.; Donnelly, A. The ecological significance of phenology in four different tree species: Effects of light and temperature on bud burst. Int. J. Biometeorol. 2011, 55, 711721. [Google Scholar] [CrossRef]
- Rollinson, C.R.; Kaye, M.W. Experimental warming alters spring phenology of certain plant functional groups in an early successional forest community. Glob. Chang. Biol. 2011, 18, 1108–1116. [Google Scholar] [CrossRef]
- Bisht, V.K.; Kuniyal, C.P.; Nautiyal, B.P.; Prasad, P. Spatial distribution and regeneration of Quercus semecarpifolia and Quercus floribunda in a subalpine forest of Western Himalaya, India. Physiol. Mol. Biol. Plants 2013, 19, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, K.I.; Toumi, J. Costs of reproduction in a historical perspective. Trends Ecol. Evol. 1994, 9, 304–306. [Google Scholar] [CrossRef]
- Vázquez-Santos, Y.; Castillo-Argüero, S.; Martínez-Orea, Y.; Sánchez-Gallen, I.; Vega-Frutis, R.; Camargo-Ricalde, S.L.; Hernández-Cuevas, L.V. The reproductive phenology of Acaena elongata and its relation with arbuscular mycorrhizal fungi. Symbiosis 2019, 79, 129–140. [Google Scholar] [CrossRef]
- Mooney, H.A.; Billings, W.D. The annual carbohydrate cycle of alpine plants as related to growth. Am. J. Bot. 1960, 47, 594–598. [Google Scholar] [CrossRef]
- Jiménez-Noriega, M.S.; López-Mata, L.; Aguilar-Rodríguez, S.; Terrazas, T. Actividad cambial y cribo-xilogenesis en tres especies en un gradiente altitudinal en la Sierra Nevada, México. Bot. Sci. 2019, 97, 725–740. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carrero, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gričar, J.; Zupančič, M.; Čufar, K. Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 2007, 41, 463–475. [Google Scholar] [CrossRef]
- Polgar, A.C.; Primack, B.R. Leaf out phenology in temperate forests. Biodivers. Sci. 2013, 21, 111–116. [Google Scholar] [CrossRef]
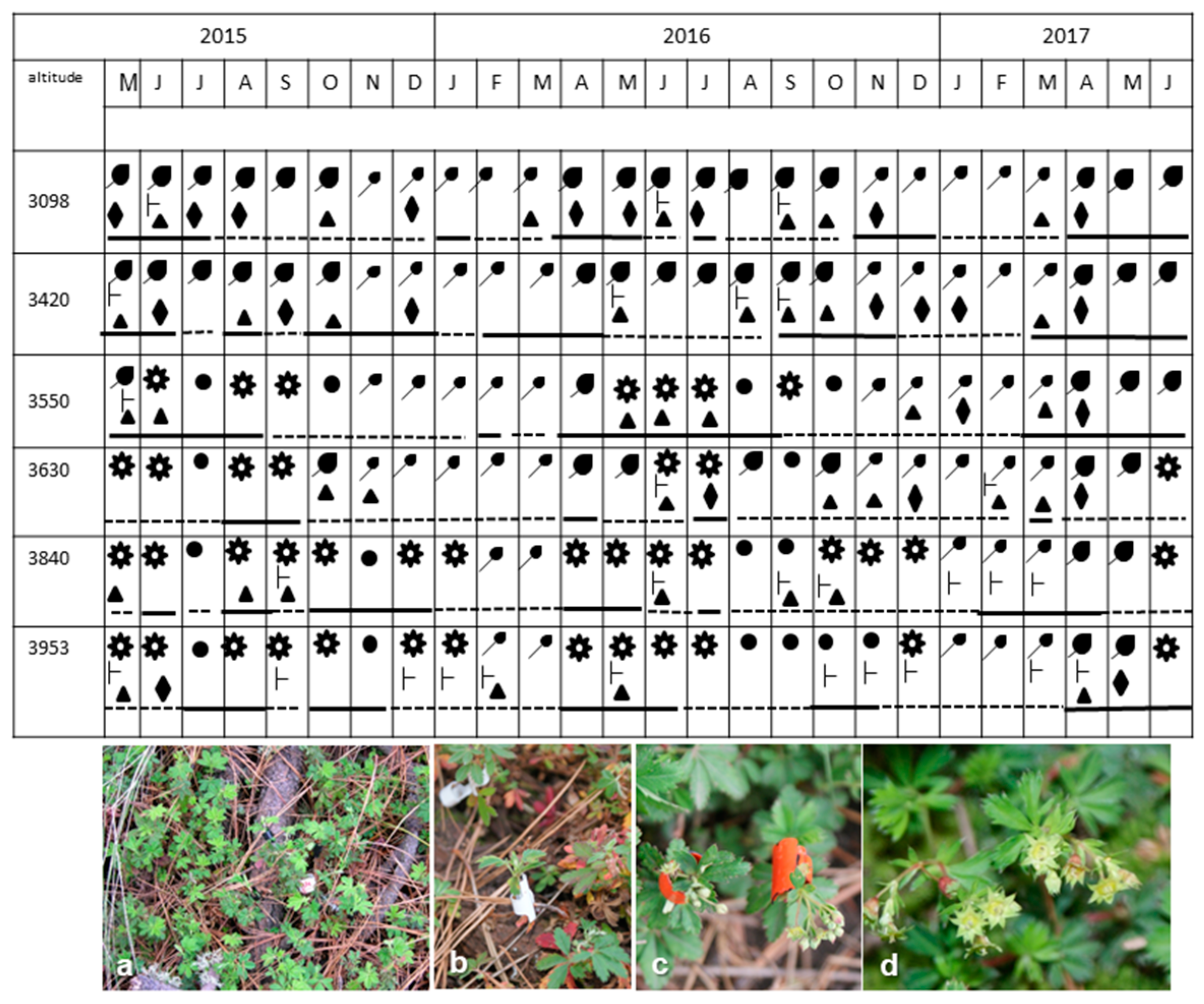
 , leaf development (11)
, leaf development (11)  , leaf development (19)
, leaf development (19)  , branch growth (31/3)
, branch growth (31/3)  , flowering (65)
, flowering (65)  , development of fruit (71/79)
, development of fruit (71/79)  , dormancy (92)
, dormancy (92)  . Cambium: active
. Cambium: active  inactive
inactive  (a–d) = different phenological phages. (a,b) leaf development and branch growth, (c) flowering, (d) development of fruit.
, leaf development (11) , leaf development (19) , branch growth (31/3) , flowering (65) , development of fruit (71/79) , dormancy (92) . Cambium: active inactive (a–d) = different phenological phages. (a,b) leaf development and branch growth, (c) flowering, (d) development of fruit.
(a–d) = different phenological phages. (a,b) leaf development and branch growth, (c) flowering, (d) development of fruit.
, leaf development (11) , leaf development (19) , branch growth (31/3) , flowering (65) , development of fruit (71/79) , dormancy (92) . Cambium: active inactive (a–d) = different phenological phages. (a,b) leaf development and branch growth, (c) flowering, (d) development of fruit.
 , leaf development (11)
, leaf development (11)  , leaf development (19)
, leaf development (19)  , branch growth (31/3)
, branch growth (31/3)  , flowering (65)
, flowering (65)  , development of fruit (71/79)
, development of fruit (71/79)  , dormancy (92)
, dormancy (92)  . Cambium: active
. Cambium: active  inactive
inactive  (a–d) = different phenological phases. (a,b) leaf development and branch growth, (c) flowering, (d) ripening.
, leaf development (11) , leaf development (19) , branch growth (31/3) , flowering (65) , development of fruit (71/79) , dormancy (92) . Cambium: active inactive (a–d) = different phenological phases. (a,b) leaf development and branch growth, (c) flowering, (d) ripening.
(a–d) = different phenological phases. (a,b) leaf development and branch growth, (c) flowering, (d) ripening.
, leaf development (11) , leaf development (19) , branch growth (31/3) , flowering (65) , development of fruit (71/79) , dormancy (92) . Cambium: active inactive (a–d) = different phenological phases. (a,b) leaf development and branch growth, (c) flowering, (d) ripening.
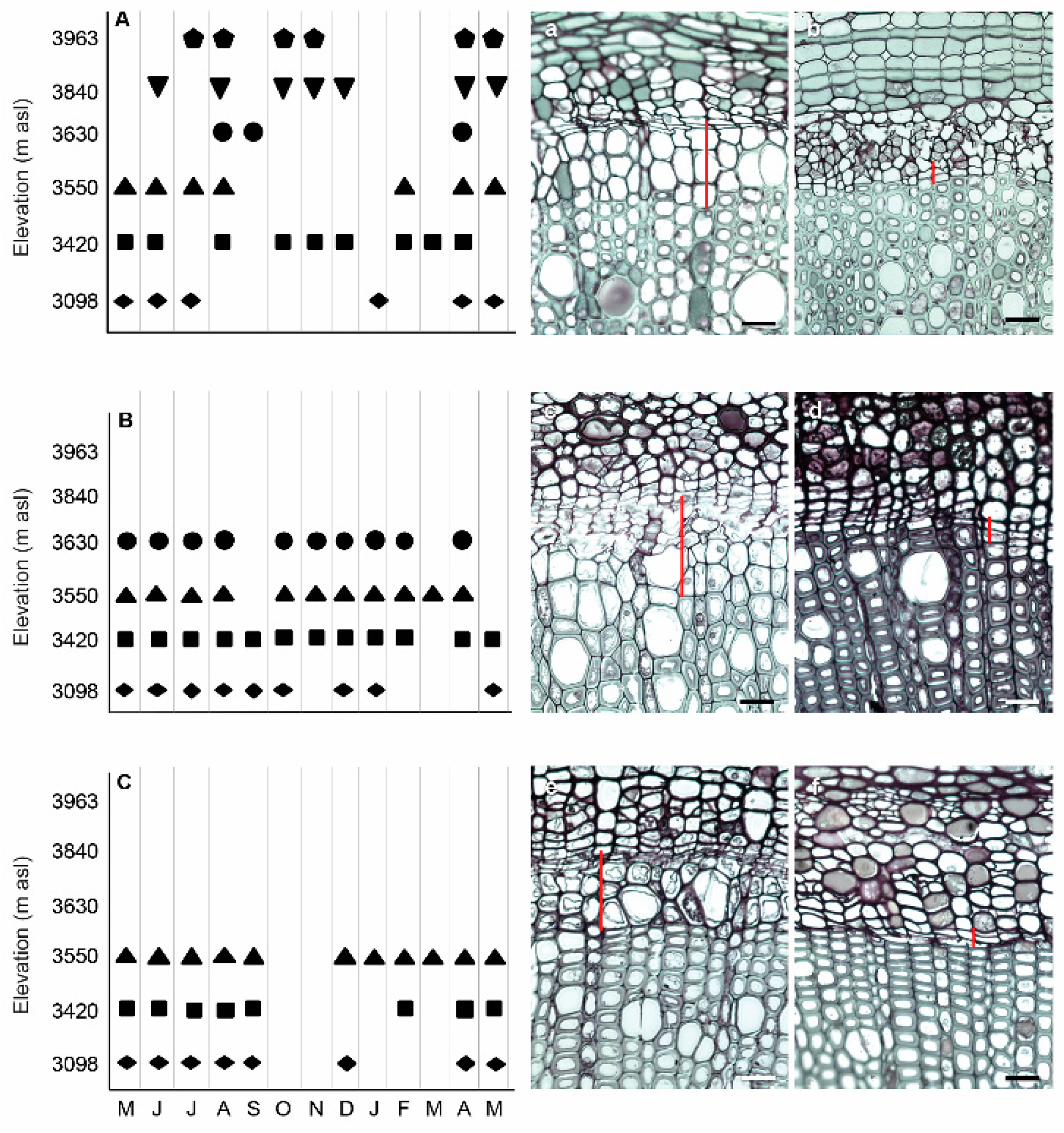
 , leaf development (11)
, leaf development (11)  , leaf development (19)
, leaf development (19)  , branch growth (31/3)
, branch growth (31/3)  , flowering (65)
, flowering (65)  , development of fruit (71/79)
, development of fruit (71/79)  , dormancy (92)
, dormancy (92)  . Cambium: active
. Cambium: active  inactive
inactive  (a–e) = different phenological phases. (a) bud-dormancy, (b) bud beak, (c) leaf development, (d) flowering, (e) fruit development.
, leaf development (11) , leaf development (19) , branch growth (31/3) , flowering (65) , development of fruit (71/79) , dormancy (92) . Cambium: active inactive (a–e) = different phenological phases. (a) bud-dormancy, (b) bud beak, (c) leaf development, (d) flowering, (e) fruit development.
(a–e) = different phenological phases. (a) bud-dormancy, (b) bud beak, (c) leaf development, (d) flowering, (e) fruit development.
, leaf development (11) , leaf development (19) , branch growth (31/3) , flowering (65) , development of fruit (71/79) , dormancy (92) . Cambium: active inactive (a–e) = different phenological phases. (a) bud-dormancy, (b) bud beak, (c) leaf development, (d) flowering, (e) fruit development.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | Code | Phenophase |
|---|---|---|
 | 07 | Beginning of bud break (first green leaf tips just visible) |
 | 11 | Leaf development (first leaves unfolded) |
 | 19 | Leaf development (first leaves fully expanded) |
 | 31/3 | Beginning of shoot or branch growth (axes) |
 | 55 | Inflorescence emergence (flower buds visible) |
 | 65 | Flowering |
 | 71/79 | Development of fruit |
 | 89 | Ripening |
 | 91 | Beginning of dormancy, beginning of axillary bud formation |
 | 92 | Beginning of dormancy, new leaves with smaller lamina |
| Altitude (m asl)—Vegetation Type | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3098 Mixed Forest | 3420 Fir Forest | 3550 Fir Forest | 3630 Pine Forest | 3840 Pine Forest–Alpine Grassland | 3963 Alpine Grassland | ||||||
| N | M | N | M | N | M | N | M | N | M | N | M |
| Alchemilla procumbens | |||||||||||
| 0.89a ± 0.30 | 1.17 B ± 0.43 | 0.40d ± 0.11 | 1.45 B ± 0.34 | 0.78b ± 0.11 | 0.80 C ± 0.12 | 0.88a ± 0.24 | 2.35 A ± 0.17 | 0.50c ± 0.06 | 0.75 C ± 0.17 | 0.56c ± 0.08 | 0.63 D ± 0.11 |
| Acaena elongata | |||||||||||
| 1.23c ± 0.53 | 3.33 B ± 2.12 | 1.53b ± 0.89 | 3.21 B ± 1.63 | 1.81a ± 0.49 | 4.44 A ± 1.06 | 0.64d ± 0.14 | 4.48 A ± 2.11 | ||||
| Ribes ciliatum | |||||||||||
| 7.83b ± 1.34 | 12.1 A ± 3.32 | 12.48a ± 1.25 | 13.78 A ± 2.80 | 3.45c ± 1.75 | 13.53 A ± 3.48 | ||||||
| Canonical Function | Eigenvalues/% Variance | ||
|---|---|---|---|
| Acaena F = 10.81 | Alchemilla F = 7.94 | Ribes F = 9.88 | |
| 1 | 3.16/39 | 2.60/59 | 3.92/38 |
| 2 | 1.93/23 | 0.78/18 | 3.39/33 |
| 3 | 0.97/12 | 0.40/9 | 1.31/13 |
| 4 | 0.82/10 | 0.22/5 | 0.77/7 |
| 5 | 0.56/7 | 0.17/4 | 0.55/5 |
| Environment | V1 | V2 | V3 | V4 | V5 |
|---|---|---|---|---|---|
| Temperature mean | 0.669 | −0.326 | 0.182 | 0.068 | 0.032 |
| Temperature minimum | 0.517 | −0.274 | 0.190 | 0.096 | 0.093 |
| Humidity maximum | 0.190 | 0.141 | 0.052 | −0.375 | 0.164 |
| Direct Site Factor minimum | −0.046 | 0.572 | 0.073 | −0.056 | −0.058 |
| Direct Site Factor maximum | 0.014 | 0.198 | −0.417 | −0.025 | −0.043 |
| Global Site Factor minimum | 0.073 | 0.565 | 0.011 | −0.117 | −0.189 |
| Plants | W1 | W2 | W3 | W4 | W5 |
| Cambium | −0.081 | 0.149 | 0.213 | −0.312 | 0.412 |
| Beginning of bud break (07) | 0.118 | −0.089 | 0.085 | 0.477 | 0.156 |
| New leaves with smaller lamina (92) | −0.700 | −0.033 | 0.130 | −0.144 | −0.058 |
| Relative growth rate | 0.704 | 0.150 | 0.365 | 0.024 | 0.037 |
| Beginning of new branches (31) | −0.339 | −0.581 | −0.030 | −0.009 | 0.094 |
| Inflorescence emergence (55) | −0.004 | −0.160 | 0.470 | 0.215 | 0.132 |
| Environment | V1 | V2 | V3 | V4 | V5 |
|---|---|---|---|---|---|
| Temperature minimum | 0.257 | 0.524 | −0.139 | 0.033 | 0.051 |
| Temperature maximum | 0.155 | −0.18 | 0.100 | −0.281 | 0.001 |
| Humidity minimum | −0.011 | 0.176 | 0.057 | 0.317 | 0.018 |
| Humidity maximum | −0.053 | 0.059 | 0.080 | 0.113 | 0.174 |
| Indirect Site Factor minimum | 0.568 | −0.351 | 0.143 | −0.065 | 0.076 |
| Global Site Factor mean | 0.562 | −0.394 | −0.086 | −0.048 | −0016 |
| Plants | W1 | W2 | W3 | W4 | W5 |
| Cambium | −0.082 | 0.355 | 0.201 | 0.257 | −0.079 |
| Leaf development (fully expanded, 19) | 0.077 | 0.194 | 0.312 | −0.046 | 0.179 |
| New leaves with smaller lamina (92) | −0.552 | −0.435 | 0.009 | −0.013 | −0.069 |
| Beginning of new branches (31) | −0.571 | 0.193 | −0.139 | 0.038 | 0.056 |
| Developing of fruits (71/79) | 0.449 | 0.021 | −0.271 | −0.001 | −0.250 |
| Environment | V1 | V2 | V3 | V4 | V5 |
|---|---|---|---|---|---|
| Temperature mean | 0.481 | 0.093 | −0.476 | 0.059 | 0.170 |
| Temperature minimum | 0.588 | −0.124 | −0.193 | 0.069 | 0.208 |
| Temperature maximum | 0.231 | 0.500 | −0.416 | 0.021 | 0.099 |
| Humidity mean | 0.474 | −0.216 | 0.449 | −0.001 | −0.007 |
| Direct Site Factor mean | 0.022 | 0.436 | −0.035 | 0.107 | −0.323 |
| Direct Site Factor maximum | 0.047 | 0.319 | −0.011 | 0.119 | −0.324 |
| Indirect Site Factor mean | −0.225 | 0.430 | 0.138 | 0.250 | 0.225 |
| Plants | W1 | W2 | W3 | W4 | W5 |
| Cambium | 0.748 | 0.348 | −0.020 | −0.068 | −0.181 |
| Beginning of bud break (07) | 0.392 | −0.282 | −0.497 | −0.103 | −0.085 |
| New leaves with smaller lamina (92) | −0.700 | −0.065 | 0.201 | −0.166 | −0.279 |
| Relative growth rate | 0.219 | 0.295 | −0.370 | 0.329 | −0.015 |
| Inflorescence emergence (55) | −0.216 | −0.152 | 0.493 | 0.065 | −0.104 |
| Ripening (89) | −0.319 | 0.631 | −0.274 | 0.202 | 0.098 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Noriega, M.S.; López-Mata, L.; Terrazas, T. Cambial Activity and Phenology in Three Understory Species along an Altitude Gradient in Mexico. Forests 2021, 12, 506. https://doi.org/10.3390/f12040506
Jiménez-Noriega MS, López-Mata L, Terrazas T. Cambial Activity and Phenology in Three Understory Species along an Altitude Gradient in Mexico. Forests. 2021; 12(4):506. https://doi.org/10.3390/f12040506
Chicago/Turabian StyleJiménez-Noriega, Mayte S., Lauro López-Mata, and Teresa Terrazas. 2021. "Cambial Activity and Phenology in Three Understory Species along an Altitude Gradient in Mexico" Forests 12, no. 4: 506. https://doi.org/10.3390/f12040506