Niche Selection by Soil Bacterial Community of Disturbed Subalpine Forests in Western Sichuan

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Sites and Soil Sampling
2.2. Soil Physiochemical Properties
2.3. DNA Extraction and Sequencing of Soil Bacteria
2.4. Data Preprocessing
2.5. Statistical Analysis
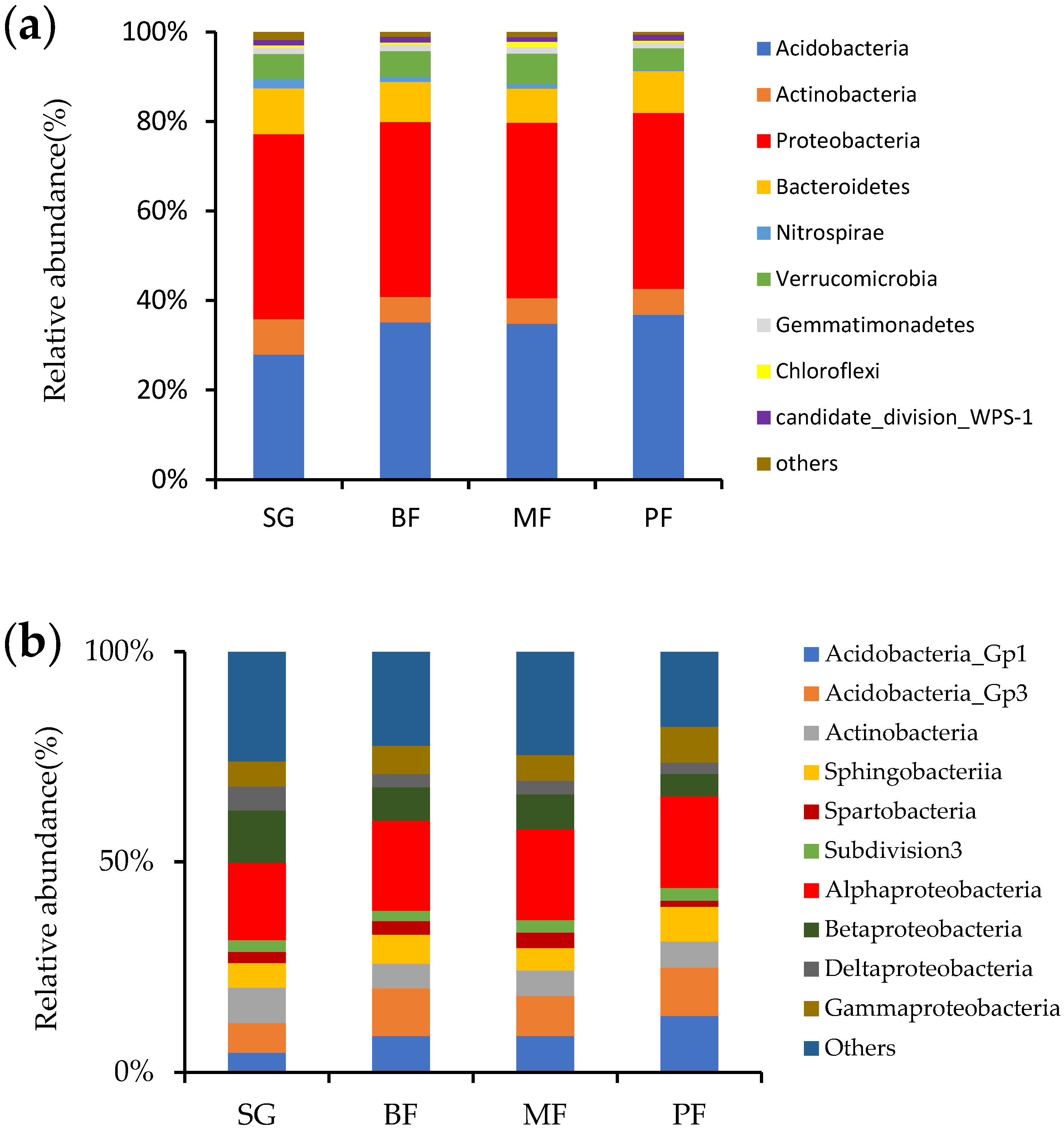
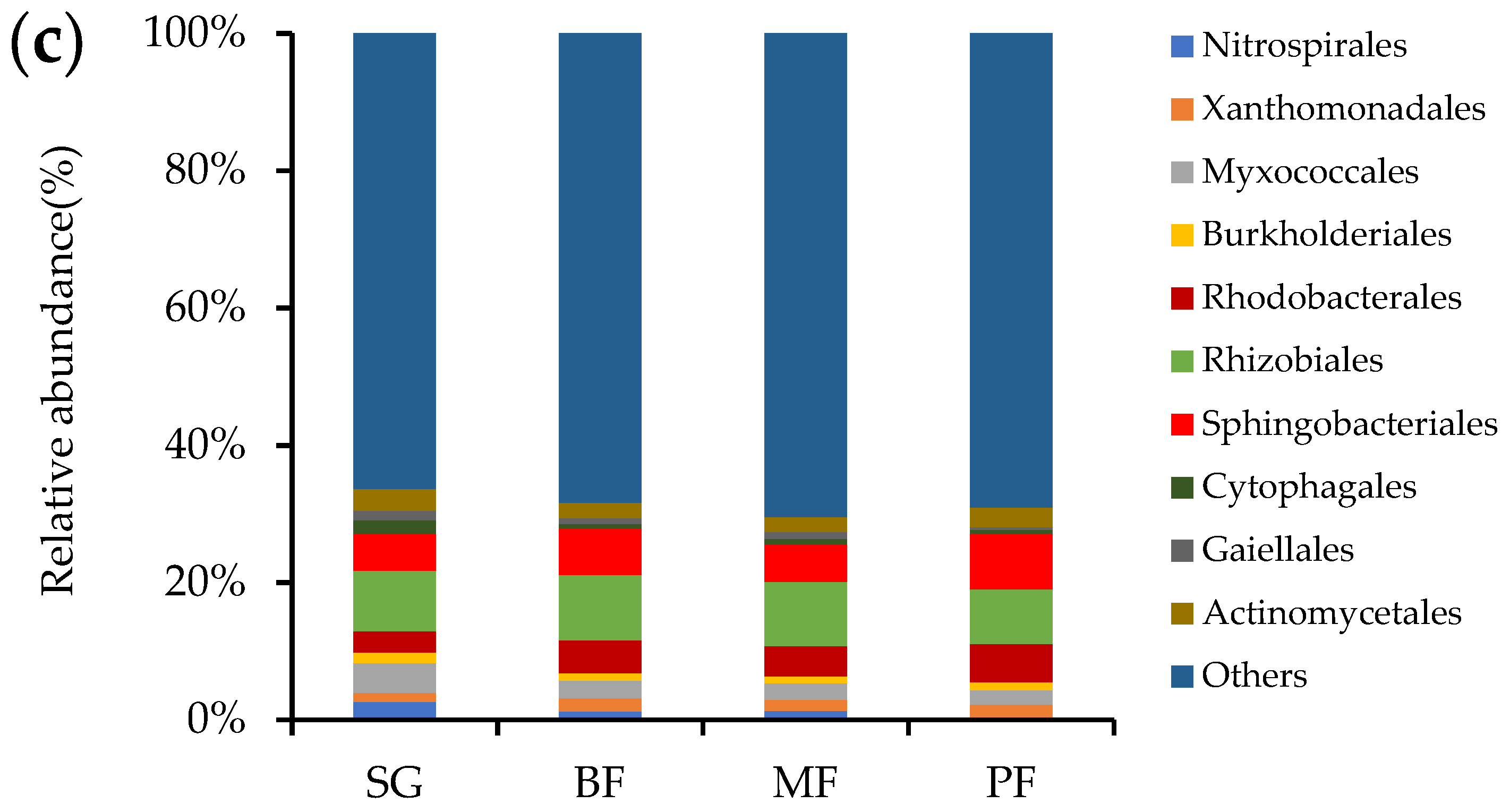
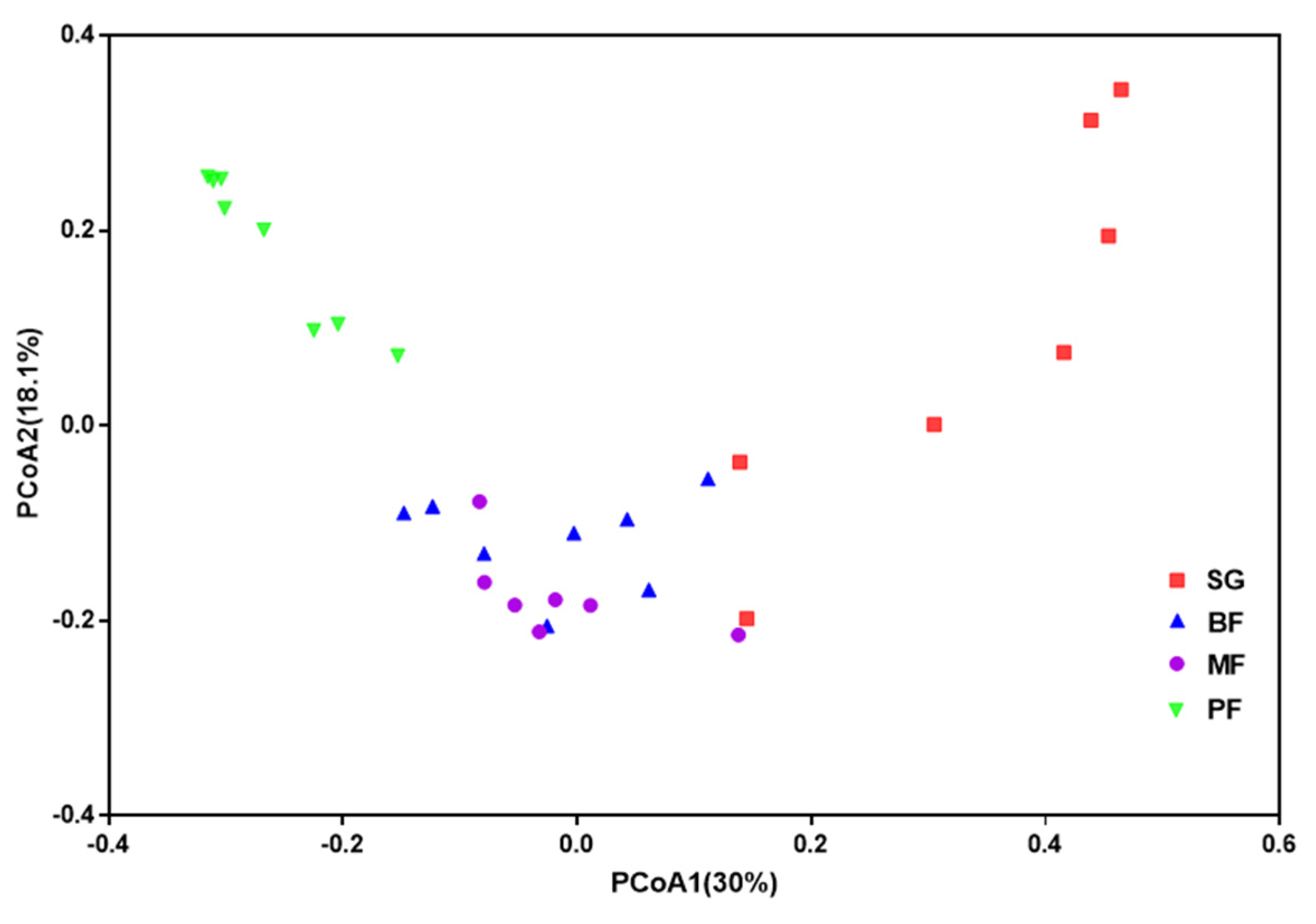
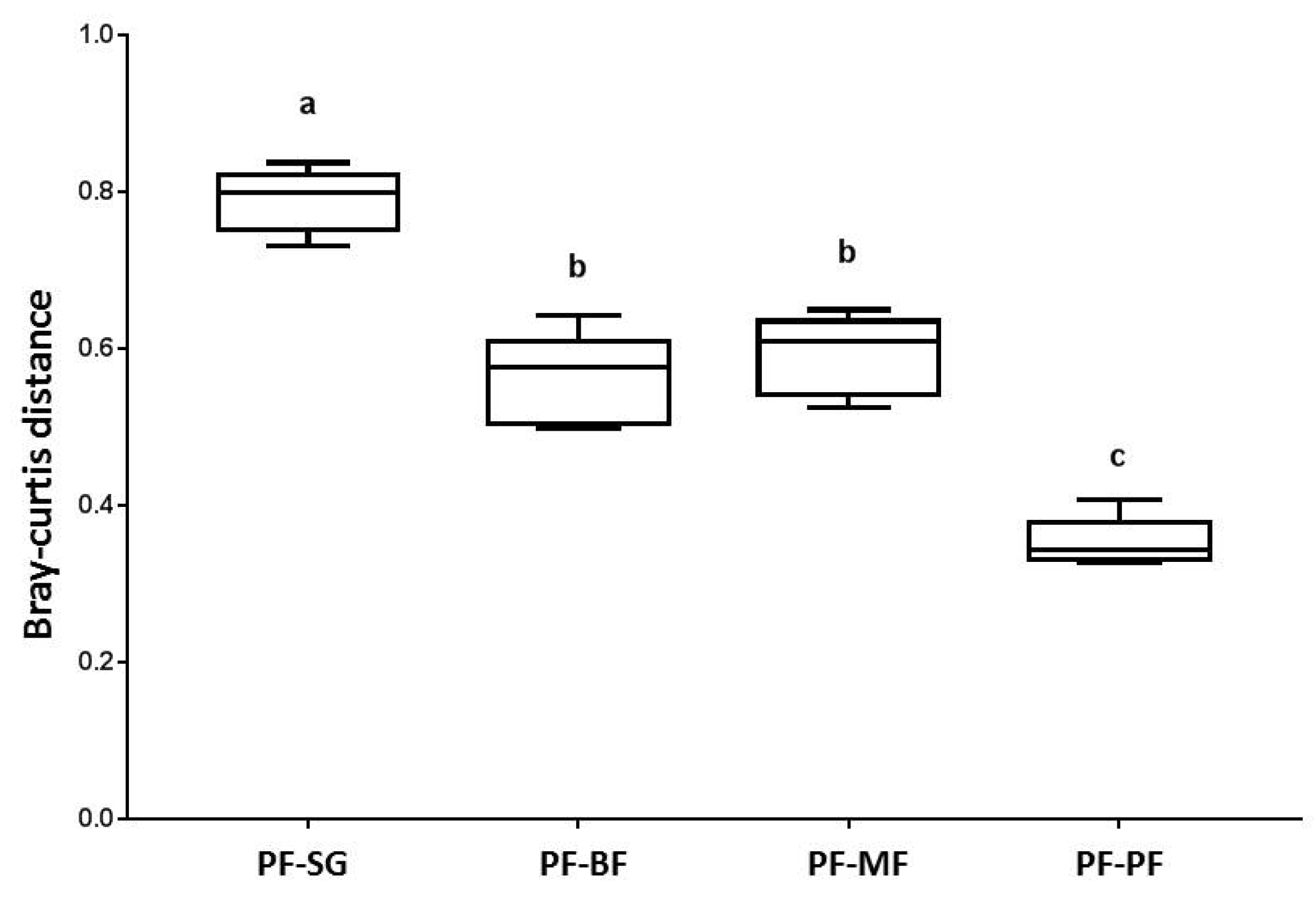
3. Results
3.1. Soil Properties of Disturbed Subalpine Forests
3.2. Bacterial Community Structure and Bray-Curtis Similarity
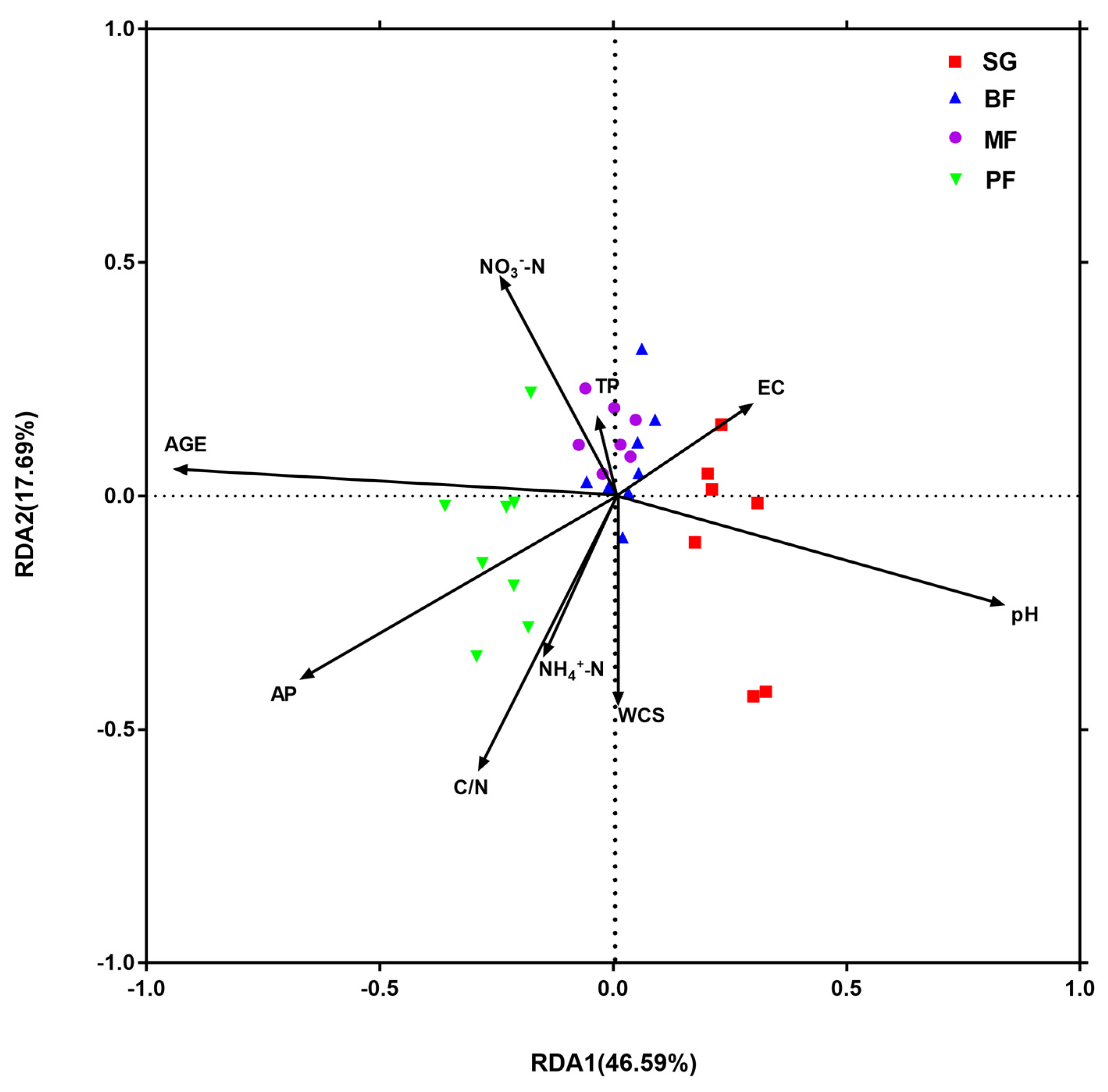
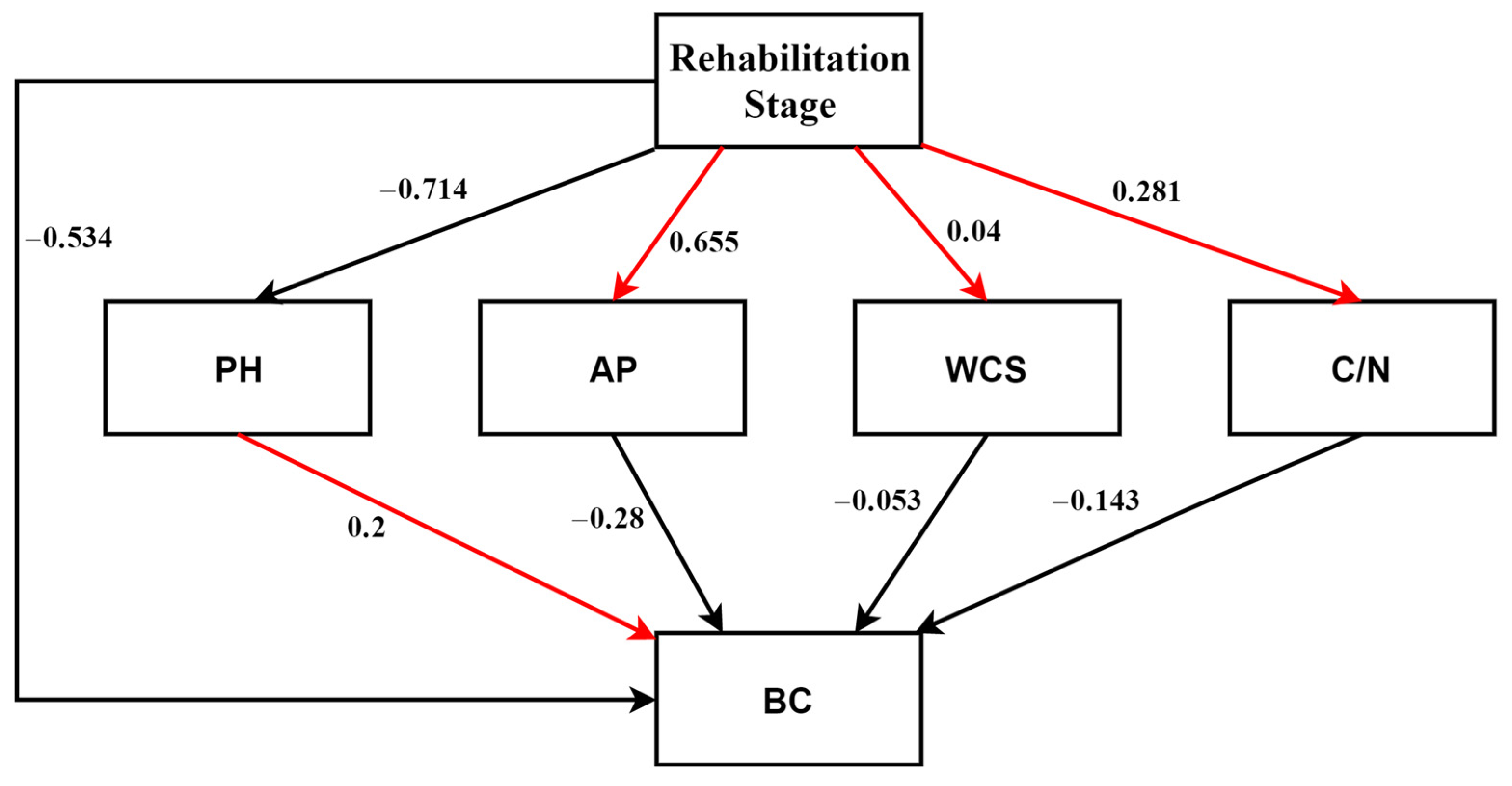
3.3. Relationships between Bacterial Community Structures and Environmetal Variables
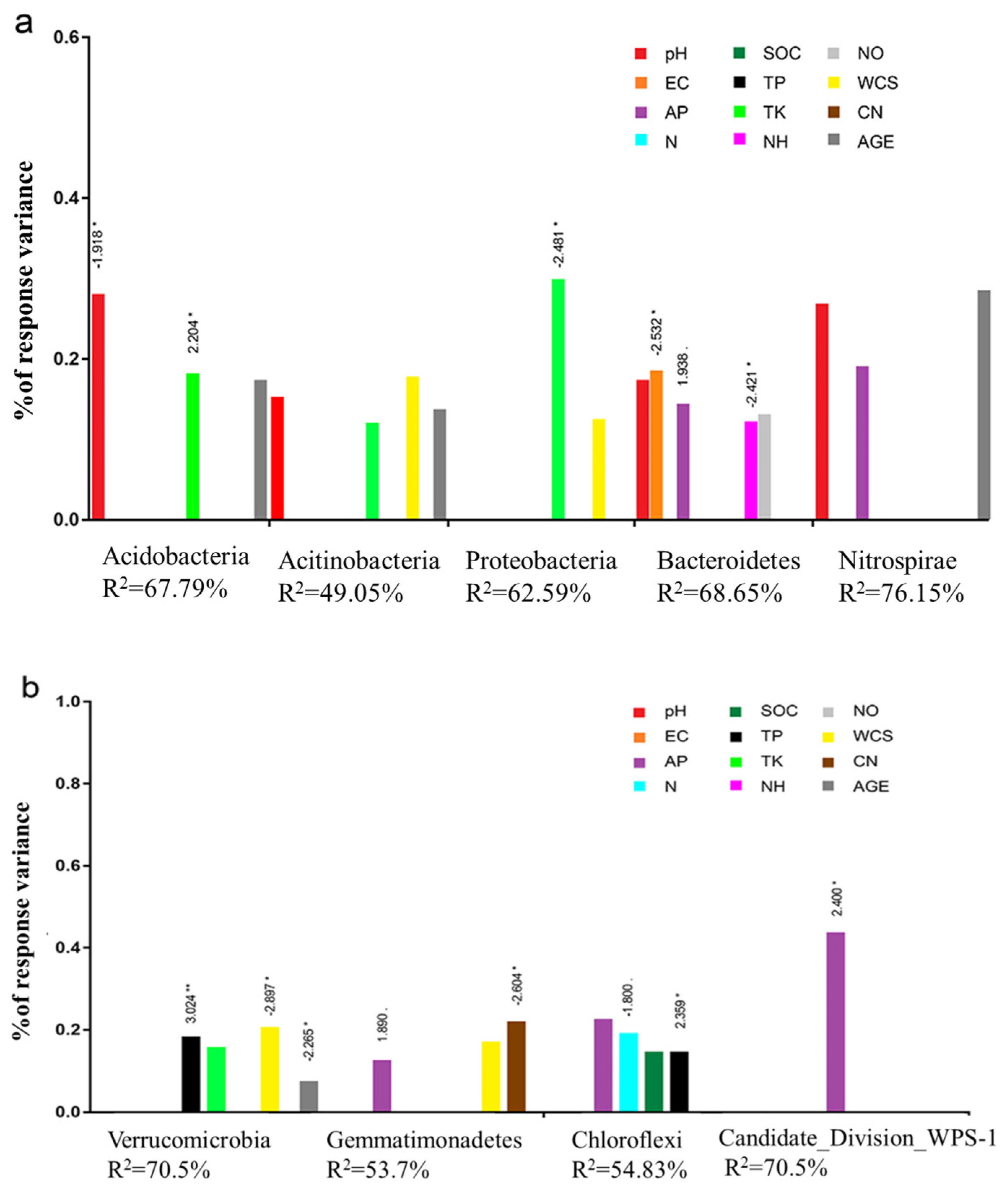
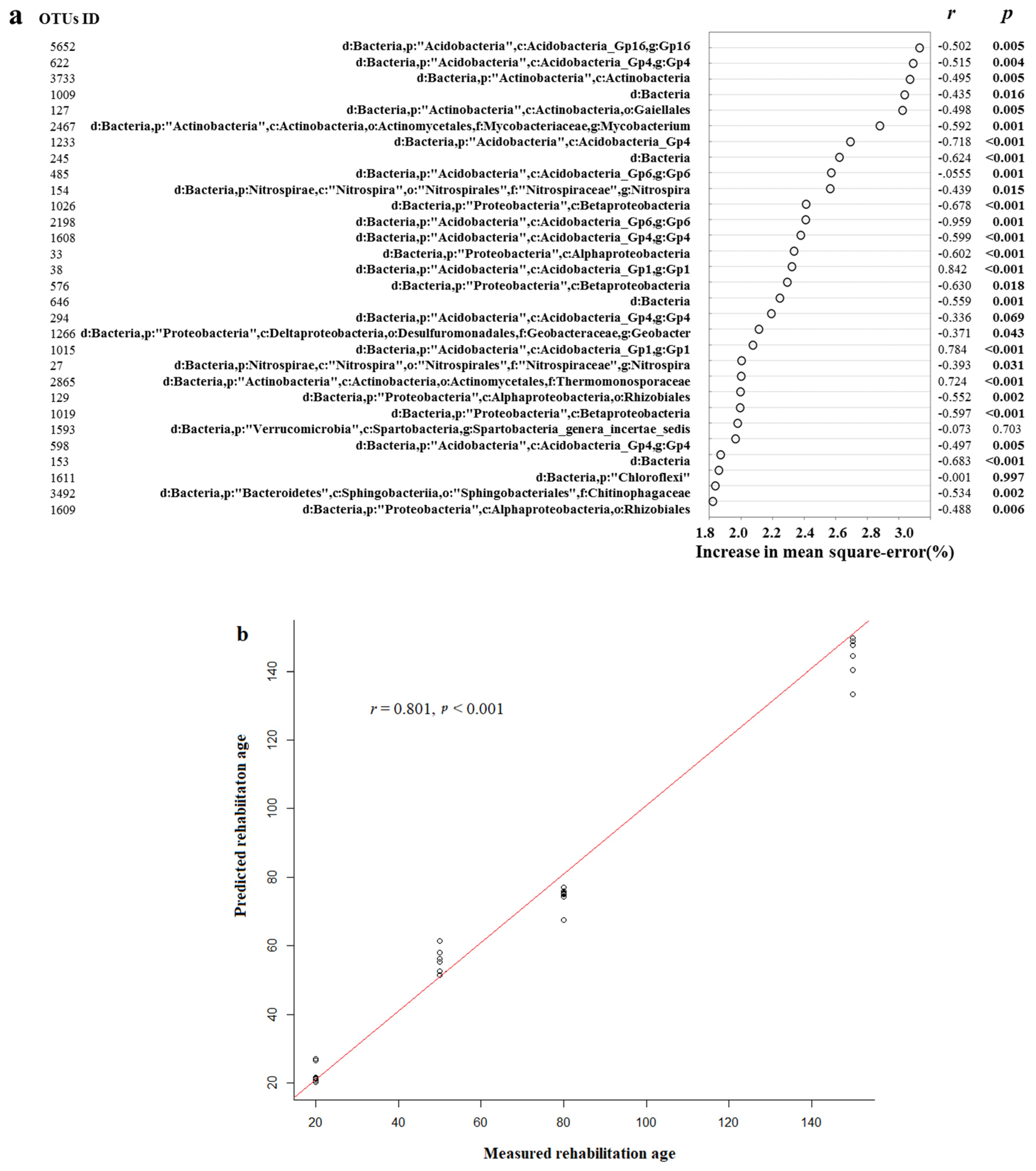
3.4. The Abundance of Sensitive Taxa Correlated with Secondary Succession
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, Z.; Wang, C.; Jiang, L.; Luo, Y. Trends in soil microbial communities during secondary succession. Soil Biol. Biochem. 2017, 115, 92–99. [Google Scholar] [CrossRef]
- Manuel, R.G.; Ostertag, R. Neotropical secondary forest succession: Changes in structural and functional characteristics. For. Ecol. Manag. 2001, 148, 185–206. [Google Scholar]
- Norden, N.; Angarita, H.A.; Bongers, F.; Martinez-Ramos, M.; Granzow-de la Cerda, I.; van Breugel, M.; Lebrija-Trejos, E.; Meave, J.A.; Vandermeer, J.; Williamson, G.B.; et al. Successional dynamics in Neotropical forests are as uncertain as they are predictable. Proc. Natl. Acad. Sci. USA 2015, 112, 8013–8018. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Bing, H.; Fang, L.; Wu, Y.; Yu, J.; Shen, G.; Jiang, M.; Wang, X.; Zhang, X. Diversity patterns of the rhizosphere and bulk soil microbial communities along an altitudinal gradient in an alpine ecosystem of the eastern Tibetan Plateau. Geoderma 2019, 338, 118–127. [Google Scholar] [CrossRef]
- Chen, Y.L.; Deng, Y.; Ding, J.Z.; Hu, H.W.; Xu, T.L.; Li, F.; Yang, G.B.; Yang, Y.H. Distinct microbial communities in the active and permafrost layers on the Tibetan Plateau. Mol. Ecol. 2017, 26, 6608–6620. [Google Scholar] [CrossRef]
- Li, J.; Shen, Z.; Li, C.; Kou, Y.; Wang, Y.; Tu, B.; Zhang, S.; Li, X. Stair-Step Pattern of Soil Bacterial Diversity Mainly Driven by pH and Vegetation Types Along the Elevational Gradients of Gongga Mountain, China. Front. Microbiol. 2018, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.C.; Xiong, J.B.; Zhang, H.Y.; Feng, Y.Z.; Lin, X.G.; Li, X.Y.; Liang, W.J.; Chu, H.Y. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.R.; Li, X.Z.; Wang, J.X.; Ding, Q.; Wang, H.; Tian, C.; Yao, M.J.; An, J.X.; Huang, Y.T. Changes of soil prokaryotic communities after clear-cutting in a karst forest: Evidences for cutting-based disturbance promoting deterministic processes. FEMS Microbiol. Ecol. 2016, 92, 3. [Google Scholar] [CrossRef]
- Cao, J.; Pan, H.; Chen, Z.; Shang, H. Bacterial, fungal, and archaeal community assembly patterns and their determining factors across three subalpine stands at different stages of natural restoration after clear-cutting. J. Soils Sediments 2020, 20, 2794–2803. [Google Scholar] [CrossRef]
- You, Y.Y.; Wang, J.; Huang, X.M.; Tang, Z.X.; Liu, S.R.; Sun, O.J. Relating microbial community structure to functioning in forest soil organic carbon transformation and turnover. Ecol. Evol. 2014, 4, 633–647. [Google Scholar] [CrossRef]
- Wan, X.H.; Huang, Z.Q.; He, Z.M.; Yu, Z.P.; Wang, M.H.; Davis, M.R.; Yang, Y.S. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Pennanen, T.; Caul, S.; Daniell, T.J.; Griffiths, B.S.; Ritz, K.; Wheatley, R.E. Community-level responses of metabolically-active soil microorganisms to the quantity and quality of substrate inputs. Soil Biol. Biochem. 2004, 36, 841–848. [Google Scholar] [CrossRef]
- Carney, K.M.; Matson, P.A. Plant Communities, Soil Microorganisms, and Soil Carbon Cycling: Does Altering the World Belowground Matter to Ecosystem Functioning? Ecosystems 2005, 8, 928–940. [Google Scholar] [CrossRef]
- Yao, M.; Rui, J.; Li, J.; Dai, Y.; Bai, Y.; Heděnec, P.; Wang, J.; Zhang, S.; Pei, K.; Liu, C.; et al. Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe. Soil Biol. Biochem. 2014, 79, 81–90. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Ivanova, N.S. Recovery of tree stand after clear-cutting in the ural mountains. Int. J. Bio-Resour. Stress Manag. 2014, 5, 090–092. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y.; Liu, S.; Zang, R.; He, F.; Spence, J.R.; Cadotte, M. Partial recovery of a tropical rain forest a half-century after clear-cut and selective logging. J. Appl. Ecol. 2015, 52, 1044–1052. [Google Scholar] [CrossRef]
- Banning, N.C.; Gleeson, D.B.; Grigg, A.H.; Grant, C.D.; Andersen, G.L.; Brodie, E.L.; Murphy, D.V. Soil microbial community successional patterns during forest ecosystem restoration. Appl. Environ. Microb. 2011, 77, 6158–6164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrenberg, S.; O’Neill, S.P.; Knelman, J.E.; Todd, B.; Duggan, S.; Bradley, D.; Robinson, T.; Schmidt, S.K.; Townsend, A.R.; Williams, M.W.; et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance. ISME J. 2013, 7, 1102–1111. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.; Lin, X.; Zhang, H.; Zeng, J.; Chu, H. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 03829. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, S.; Wang, Z.; Zhang, Z.; Qin, H.; Wei, Z.; Feng, K.; Li, S.; Wu, Y.; Yin, H.; et al. Soil microbiome mediated nutrients decline during forest degradation process. Soil Ecol. Lett. 2019, 1, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Wang, Y.; Zhang, Y.; Zhang, X. Responses of soil bacterial communities, enzyme activities, and nutrients to agricultural-to-natural ecosystem conversion in the Loess Plateau, China. J. Soils Sediments 2018, 19, 1427–1440. [Google Scholar] [CrossRef]
- Allison, S.D. A trait-based approach for modelling microbial litter decomposition. Ecol. Lett. 2012, 15, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Thimo, K.; Klaus, K.; Georg, G.; Christiane, G.; Karsten, K. A new conceptual model for the fate of lignin in decomposing plant litter. Ecology 2011, 92, 1052–1062. [Google Scholar]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial contributions to delignification and lignocellulose degradation in forest soils with metagenomic and quantitative stable isotope probing. ISME J. 2019, 13, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Fischer, T.; Agarwal, A.; Hess, H. A smart dust biosensor powered by kinesin motors. Nat. Nanotechnol. 2009, 4, 162–166. [Google Scholar] [CrossRef]
- Wu, J.; Park, J.P.; Dooley, K.; Cropek, D.M.; West, A.C.; Banta, S. Rapid development of new protein biosensors utilizing peptides obtained via phage display. PLoS ONE. 2011, 6, e24948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbeta, A.; Julio Camarero, J.; Sanguesa-Barreda, G.; Muffler, L.; Penuelas, J. Contrasting effects of fog frequency on the radial growth of two tree species in a Mediterranean-temperate ecotone. Agric. For. Meteorol. 2019, 264, 297–308. [Google Scholar] [CrossRef]
- Xuan, L.; Sheng, Z.; Lu, J.; Qiu, Q.; Chen, J.; Xiong, J. Bacterioplankton community responses and the potential ecological thresholds along disturbance gradients. Sci. Total Environ. 2019, 696, 134015. [Google Scholar] [CrossRef]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment. Sci. Total Environ. 2016, 566–567, 949–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, Y.; Liu, S. Aboveground biomass, ANPP and stem volume of birch stands in natural restoration process of subalpine secondary forest in Western Sichuan. Acta Ecol. Sin. 2010, 30, 594–601, (In Chinese with English abstract). [Google Scholar]
- Zhang, Y.; Liu, S.; Ma, J.; Shi, Z.; Liu, X. Woodland hydrological effects of birch forests in sub-alpine region of western Sichuan, China. Acta Ecol. Sin. 2005, 25, 2939–2946, (In Chinese with English abstract). [Google Scholar]
- Shi, L.X.; Wang, J.X.; Su, Y.M.; Hou, G.W. Earlier-stage succession of vegetation on the clear-cuts in mialou forest district in western Sichuan. Acta Phytoecol. Sci. 1988, 12, 306–313, (In Chinese with English abstract). [Google Scholar]
- Wang, Q.; Wu, N.; Luo, P. Simulation of light regimes in typical subalpine forest succession series of eastern Tibetan Plateau. Pol. J. Ecol. 2009, 57, 251–260, (In Chinese with English abstract). [Google Scholar]
- Jiang, Y.; Lei, Y.; Yang, Y.; Korpelainen, H.; Niinemets, Ü.; Li, C. Divergent assemblage patterns and driving forces for bacterial and fungal communities along a glacier forefield chronosequence. Soil Biol. Biochem. 2018, 118, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Shi, Z.; Zhang, Y.; Chen, B. Quantitative analysis of different restoration stages during natural succession processes of subalpine dark brown coniferous forests in western Sichuan, China. Chin. J. Appl. Ecol. 2007, 18, 1695–1701, (In Chinese with English abstract). [Google Scholar]
- Xu, Z.; Yin, H.; Xiong, P.; Wan, C.; Liu, Q. Short-term responses of Picea asperata seedlings of different ages grown in two contrasting forest ecosystems to experimental warming. Environ. Exp. Bot. 2012, 77, 1–11. [Google Scholar] [CrossRef]
- Jones, D.; Willett, V. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA Circular 939; US Government Printing Office: Washington, DC, USA, 1954.
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.; Fierer, N.; Pena, A.; Goodrich, J.; Gordon, J. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.; Tiedje, J.; Cole, J. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Kenkel, N.; Orlóci, L. Applying metric and nonmetric multidimensional scaling to ecological studies: Some new results. Ecology 1986, 67, 919–928. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Stevens, M.; Oksanen, M. Vegan: Community Ecology Package–R Package v.2.3–0; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.r-project.org (accessed on 16 April 2021).
- McArdle, B.; Anderson, M. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: http://www.r-project.org (accessed on 16 April 2021).
- Grömping, U. Relative importance for linear regression in R: The package relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, G. PLS Path Modeling with R; Trowchez Editions: Berkeley, CA, USA, 2013. [Google Scholar]
- Cutler, D.; Edwards, T.; Beard, K.; Cutler, A.; Hess, K.; Gibson, J.; Lawler, J. Random Forests or classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.; Van Der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Fierer, N.; Nemergut, D.; Knight, R.; Craine, J.M. Changes through time: Integrating microorganisms into the study of succession. Res. Microbiol. 2010, 161, 635–642. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Norton, D.A. Towards a Conceptual Framework for Restoration Ecology. Restor. Ecol. 1996, 4, 93–110. [Google Scholar] [CrossRef]
- Rui, J.P.; Li, J.B.; Wang, S.P.; An, J.X.; Liu, W.T.; Lin, Q.Y.; Yang, Y.F.; He, Z.L.; Li, X.Z. Responses of Bacterial Communities to Simulated Climate Changes in Alpine Meadow Soil of the Qinghai-Tibet Plateau. Appl. Environ. Microbiol. 2015, 81, 6070–6077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.T.; Cao, P.; Hu, H.W.; Li, J.; Han, L.L.; Zhang, L.M.; Zheng, Y.M.; He, J.Z. Altitudinal distribution patterns of soil bacterial and archaeal communities along mt. Shegyla on the Tibetan Plateau. Microb. Ecol. 2015, 69, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.X.; Fang, L.C.; Guo, X.B.; Wang, X.; Zhang, Y.J.; Li, P.F.; Zhang, X.C. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Güsewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Davidson, E.; Verchot, L.; Cattânio, J.; Ackerman, I.; Carvalho, J. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry 2000, 48, 53–69. [Google Scholar] [CrossRef]
- Xu, M.; Qi, Y. Soil-surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Glob. Chang. Biol. 2001, 7, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Zheng, H.; Zhao, H.; Xie, S.; Liu, L.; Ouyang, Z. Land-use intensity indirectly affects soil multifunctionality via a cascade effect of plant diversity on soil bacterial diversity. Glob. Ecol. Conserv. 2020, 23, e01061. [Google Scholar] [CrossRef]
- Tsyganov, A.N.; Milbau, A.; Beyens, L. Environmental factors influencing soil testate amoebae in herbaceous and shrubby vegetation along an altitudinal gradient in subarctic tundra (Abisko, Sweden). Eur. J. Protistol. 2013, 49, 238–248. [Google Scholar] [CrossRef]
- Bates, S.; Clemente, J.; Flores, G.; Walters, W.; Parfrey, L.; Knight, R.; Fierer, N. Global biogeography of highly diverse protistan communities in soil. ISME J. 2013, 7, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Lim, H.; Hyun, C.; Cho, A.; Noh, H.; Hong, S.; Kim, O. Local-scale variation of soil bacterial communities in ice-free regions of maritime Antarctica. Soil Biol. Biochem. 2019, 133, 165–173. [Google Scholar] [CrossRef]
- Stursova, M.; Zifcakova, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. FEMS Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Baath, E. Fungal and bacterial growth in soil with plant materials of different C/N ratios. FEMS Microbiol. Ecol. 2007, 62, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marklein, A.; Houlton, B. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef]
- Bryant, D.A.; Liu, Z.F.; Li, T.; Zhao, F.Q.; Garcia Costas, A.M.; Klatt, C.G.; Ward, D.M.; Frigaard, N.U.; Overmann, J. Comparative and functional genomics of anoxygenic green bacteria from the taxa Chlorobi, Chloroflexi, and Acidobacteria. In Functional Genomics and Evolution of Photosynthetic Systems; Advances in Photosynthesis and Respiration; Burnap, R., Vermaas, W., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 3, pp. 47–102. [Google Scholar]
- Fierer, N.; Bradford, M.; Jackson, R. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Agren, G.; Bosatta, N. Theoretical analysis of the long-term dynamics of carbon and nitrogen in soils. Ecology 1987, 68, 1181–1189. [Google Scholar] [CrossRef]
- Youssef, N.H.; Elshahed, M.S. Diversity rankings among bacterial lineages in soil. ISME J. 2009, 3, 305–313. [Google Scholar] [CrossRef]
- Serkebaeva, Y.; Kim, Y.; Liesack, W.; Dedysh, S. Pyrosequencing-based assessment of the bacteria diversity in surface and subsurface peat layers of a northern wetland, with focus on poorly studied phyla and candidate divisions. PLoS ONE. 2013, 8, e63994. [Google Scholar] [CrossRef] [Green Version]
- Vecherskaya, M.; Dijkema, C.; Saad, H.; Stams, A. Microaerobic and anaerobic metabolism of a Methylocystis parvus strain isolated from a denitrifying bioreactor. Environ. Microbiol. Rep. 2009, 1, 442–449. [Google Scholar] [CrossRef]
- Ai, C.; Liang, G.; Sun, J.; Wang, X.; He, P.; Zhou, W.; He, X. Reduced dependence of rhizosphere microbiome on plant-derived carbon in 32-year long-term inorganic and organic fertilized soils. Soil Biol. Biochem. 2015, 80, 70–78. [Google Scholar] [CrossRef]
- Shen, C.; Ni, Y.; Liang, W.; Wang, J.; Chu, H. Distinct soil bacterial communities along a small-scale elevational gradient in alpine tundra. Front. Microbiol. 2015, 6, 00582. [Google Scholar] [CrossRef] [Green Version]
- Marshall, V. Impacts of forest harvesting on biological processes in northern forest soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Jones, R.; Robeson, M.; Lauber, C.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Dimitriu, P.; Grayston, S. Relationship between soil properties and patterns of bacterial beta-diversity across reclaimed and natural boreal forest soils. Microb. Ecol. 2010, 59, 563–573. [Google Scholar] [CrossRef]
- Chu, H.; Fierer, N.; Lauber, C.; Caporaso, J.; Knight, R.; Grogan, P. Soil bacterial diversity in the Arctic is not fundamentally different from that found in other biomes. Environ. Microbiol. 2010, 12, 2998–3006. [Google Scholar] [CrossRef] [PubMed]
- BÁRcenas-Moreno, G.; GÓMez-BrandÓN, M.; Rousk, J.; BÅÅTh, E. Adaptation of soil microbial communities to temperature: Comparison of fungi and bacteria in a laboratory experiment. Glob. Chang. Biol. 2009, 15, 2950–2957. [Google Scholar] [CrossRef]
- Xiong, J.; Ye, X.; Wang, K.; Chen, H.; Hu, C.; Zhu, J.; Zhang, D. Biogeography of the sediment bacterial community responds to a nitrogen pollution gradient in the East China Sea. Appl. Environ. Microbiol. 2014, 80, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Hui, N.; Jumpponen, A.; Francini, G.; Kotze, D.; Liu, X.; Romantschuk, M.; Strömmer, R.; Setälä, H. Soil microbial communities are shaped by vegetation type and park age in cities under cold climate. Environ. Microbiol. 2017, 19, 1281–1295. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SG | BF | MF | PF | |
|---|---|---|---|---|
| pH | 6.16 ± 0.568 a | 5.27 ± 0.31 b | 5.18 ± 0.23 b,c | 4.76 ± 0.41 c |
| EC (μs/cm) | 128 ± 27.08 a | 123.75 ± 56.37 a | 136.97 ± 22.84 a | 109.53 ± 32.17 a |
| WCS (%) | 44.36 ± 11.64 a | 44.59 ± 8.83 a | 44.34 ± 4.33 a | 45.2 ± 5.83 a |
| SOC (%) | 11.42 ± 5.79 a,b | 12.05 ± 4.71 a,b | 8.12 ± 2.75 b | 14.68 ± 5.56 a |
| TN (%) | 0.84 ± 0.32 a,b | 0.93 ± 0.32 a,b | 0.65 ± 0.18 b | 1.02 ± 0.35 a |
| TP (g/kg) | 0.99 ± 0.27 a | 1.10 ± 0.32 a | 1.14 ± 0.37 a | 1.02 ± 0.17 a |
| TK (g/kg) | 10.93 ± 4.14 b | 12.23 ± 3.41 a,b | 14.90 ± 2.62 a | 10.08 ± 2.73 b |
| NH4+-N (mg/kg) | 7.61 ± 1.82 a | 6.09 ± 2.20 a | 6.66 ± 2.62 a | 7.87 ± 1.84 a |
| NO3−-N (mg/kg) | 1.89 ± 1.17 b | 4.36 ± 1.79 a | 3.80 ± 2.56 a,b | 2.94 ± 1.69 a,b |
| AP (mg/kg) | 15.29 ± 7.56 b | 17.22 ± 4.92 b | 13.52 ± 3.14 b | 31.57 ± 8.92 a |
| AK (g/kg) | 0.20 ± 0.08 b | 0.29 ± 0.07 a | 0.26 ± 0.08 a | 0.27 ± 0.04 a |
| C/N | 13.26 ± 2.50 a,b | 12.67 ± 0.97 a,b | 12.25 ± 0.94 b | 14.19 ± 1.13 a |
| SG | BF | MF | PF | |
|---|---|---|---|---|
| SG | 0.603 | 0.565 | 0.859 | |
| BF | 0.001 | 0.181 | 0.891 | |
| MF | 0.001 | 0.066 | 0.958 | |
| PF | 0.001 | 0.001 | 0.003 |
| Variable | SS (Trace) | Pseudo-F | p | Prop | Cumulative |
|---|---|---|---|---|---|
| pH | 5823.2225 | 10.7593 | 0.001 | 0.2776 | 0.2776 |
| AP | 1665.0396 | 3.3327 | 0.003 | 0.0794 | 0.357 |
| WCS | 1425.8123 | 3.073 | 0.003 | 0.068 | 0.4249 |
| C/N | 926.7141 | 2.0803 | 0.024 | 0.0442 | 0.4691 |
| Rehabilitation stage | 510.6014 | 1.1532 | 0.2480 | 0.0243 | 0.4935 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, Z.; Zhu, W.; Yao, H.; Shu, S.; Li, X.; Ma, S.; Li, Y.; Xiong, J. Niche Selection by Soil Bacterial Community of Disturbed Subalpine Forests in Western Sichuan. Forests 2021, 12, 505. https://doi.org/10.3390/f12040505
Sheng Z, Zhu W, Yao H, Shu S, Li X, Ma S, Li Y, Xiong J. Niche Selection by Soil Bacterial Community of Disturbed Subalpine Forests in Western Sichuan. Forests. 2021; 12(4):505. https://doi.org/10.3390/f12040505
Chicago/Turabian StyleSheng, Zheliang, Wanze Zhu, Huaiying Yao, Shumiao Shu, Xia Li, Shenglan Ma, Yaying Li, and Jinbo Xiong. 2021. "Niche Selection by Soil Bacterial Community of Disturbed Subalpine Forests in Western Sichuan" Forests 12, no. 4: 505. https://doi.org/10.3390/f12040505